
Advances in Lipidomics for Cancer Biomarkers Discovery

 , ,
, ,  , ,
, ,  , and
, and 
Abstract
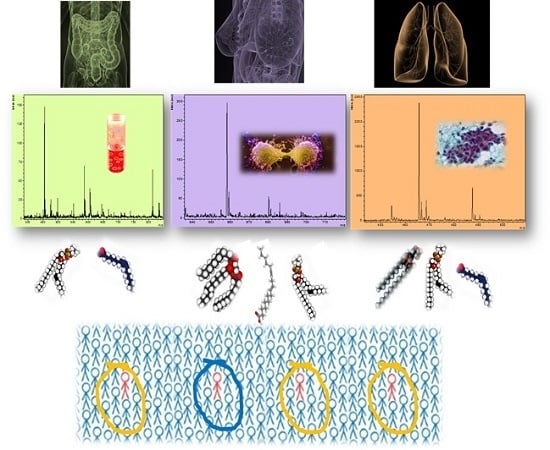
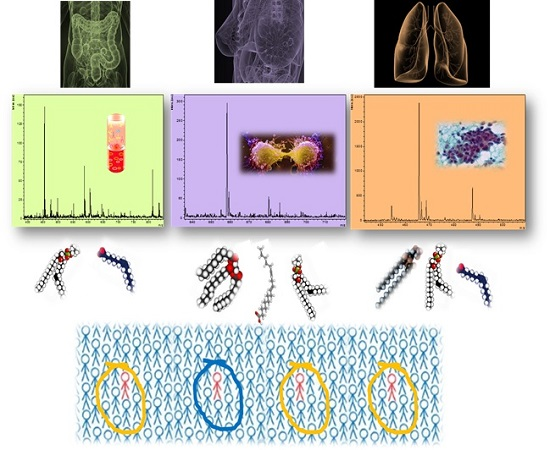
:
1. Introduction
2. Lipid Classes
3. Sample Preparation
4. Analytical Technology in Lipidomics
4.1. Electrospray Ionization Mass Spectrometry (ESI-MS/MS)
4.2. Matrix-Assisted Laser Desorption/Ionization Time-of-Flight/Mass Spectrometry (MALDI-TOF/MS)
5. Lipidomics in Cancer Research
5.1. Lung Cancer
5.2. Breast Cancer
5.3. Prostate Cancer
5.4. Colorectal Cancer
5.5. Ovarian Cancer
5.6. Pancreatic Cancer
5.7. Gastric Cancer
5.8. Bladder Cancer
5.9. Esophageal Carcinoma
5.10. Kidney Cancer
5.11. Thyroid Cancer
6. Lipidomic Highlights in Cancer Research
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, 359–386. [Google Scholar] [CrossRef] [PubMed]
- Szasz, A.M.; Gyorffy, B.; Marko-Varga, G. Cancer heterogeneity determined by functional proteomics. Semin. Cell Dev. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Bandu, R.; Mok, H.J.; Kim, K.P. Phospholipids as cancer biomarkers: Mass spectrometry-based analysis. Mass Spectrom. Rev. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, P.T.; Cerda, B.A.; Horn, D.M.; Cohen, J.S.; McLafferty, F.W.; Brown, H.A. Electrospray ionization mass spectrometry analysis of changes in phospholipids in RBL-2H3 mastocytoma cells during degranulation. Proc. Natl. Acad. Sci. USA 2001, 98, 7152–7157. [Google Scholar] [CrossRef] [PubMed]
- Astigarraga, E.; Barreda-Gomez, G.; Lombardero, L.; Fresnedo, O.; Castano, F.; Giralt, M.T.; Ochoa, B.; Rodriguez-Puertas, R.; Fernandez, J.A. Profiling and imaging of lipids on brain and liver tissue by matrix-assisted laser desorption/ionization mass spectrometry using 2-mercaptobenzothiazole as a matrix. Anal. Chem. 2008, 80, 9105–9114. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Simons, K. Lipidomics: Coming to grips with lipid diversity. Nat. Rev. Mol. Cell Biol. 2010, 11, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Wenk, M.R. The emerging field of lipidomics. Nat. Rev. Drug Discov. 2005, 4, 594–610. [Google Scholar] [CrossRef] [PubMed]
- Adibhatla, R.M.; Hatcher, J.F.; Dempsey, R.J. Lipids and lipidomics in brain injury and diseases. AAPS J. 2006, 8, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Miao, H.; Cheng, X.L.; Wei, F. Lipidomics: Novel insight into the biochemical mechanism of lipid metabolism and dysregulation-associated disease. Chem. Biol. Interact. 2015, 240, 220–238. [Google Scholar] [CrossRef] [PubMed]
- Tuncer, S.; Banerjee, S. Eicosanoid pathway in colorectal cancer: Recent updates. World J. Gastroenterol. 2015, 21, 11748–11766. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Gross, R.W. Global analyses of cellular lipidomes directly from crude extracts of biological samples by ESI mass spectrometry: A bridge to lipidomics. J. Lipid Res. 2003, 44, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, M.; Geloen, A.; Record, M.; Vance, D.; Spener, F. Lipidomics is emerging. Biochim. Biophys. Acta 2003, 1634, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Bennett, R.; Strum, J.; Ellsworth, B.B.; Hamilton, D.; Tomlinson, M.; Wolf, R.W.; Housley, M.; Roberts, B.A.; Welsh, J.; et al. Screening phosphatidylcholine biomarkers in mouse liver extracts from a hypercholesterolemia study using ESI-MS and chemometrics. Anal. Bioanal. Chem. 2009, 393, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.K.; Lee, H.S.; Park, Y.S.; Lee, J.H.; Lee, S.C.; Lee, J.H.; Lee, S.J.; Shanta, S.R.; Park, H.M.; Kim, H.R.; et al. Lipid MALDI profile classifies non-small cell lung cancers according to the histologic type. Lung Cancer 2012, 76, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–861. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.N.; Wang, H.Y.; Woods, A.S. Direct profiling of lipid distribution in brain tissue using MALDI-TOFMS. Anal. Chem. 2005, 77, 4523–4527. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.N.; Wang, H.Y.; Woods, A.S. In situ structural characterization of glycerophospholipids and sulfatides in brain tissue using MALDI-MS/MS. J. Am. Soc. Mass Spectrom. 2007, 18, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Allegood, J.; Liu, Y.; Wang, E.; Cachon-Gonzalez, B.; Cox, T.M.; Merrill, A.H.; Sullards, M.C. Imaging MALDI mass spectrometry using an oscillating capillary nebulizer matrix coating system and its application to analysis of lipids in brain from a mouse model of Tay-Sachs/Sandhoff disease. Anal. Chem. 2008, 80, 2780–2788. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Y.; Allegood, J.; Wang, E.; Cachon-Gonzalez, B.; Cox, T.M.; Merrill, A.H.; Sullards, M.C. Imaging MALDI mass spectrometry of sphingolipids using an oscillating capillary nebulizer matrix application system. Methods Mol. Biol. 2010, 656, 131–146. [Google Scholar] [PubMed]
- Hayasaka, T.; Goto-Inoue, N.; Sugiura, Y.; Zaima, N.; Nakanishi, H.; Ohishi, K.; Nakanishi, S.; Naito, T.; Taguchi, R.; Setou, M. Matrix-assisted laser desorption/ionization quadrupole ion trap time-of-flight (MALDI-QIT-TOF)-based imaging mass spectrometry reveals a layered distribution of phospholipid molecular species in the mouse retina. Rapid Commun. Mass Spectrom. 2008, 22, 3415–3426. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, Y.; Konishi, Y.; Zaima, N.; Kajihara, S.; Nakanishi, H.; Taguchi, R.; Setou, M. Visualization of the cell-selective distribution of PUFA-containing phosphatidylcholines in mouse brain by imaging mass spectrometry. J. Lipid Res. 2009, 50, 1776–1788. [Google Scholar] [CrossRef] [PubMed]
- Greene, E.R.; Huang, S.; Serhan, C.N.; Panigrahy, D. Regulation of inflammation in cancer by eicosanoids. Prostaglandins Other Lipid Mediat. 2011, 96, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.J.; Kwon, Y.W.; Jang, I.H.; Kim, D.K.; Lee, S.I.; Choi, E.J.; Kim, K.H.; Suh, D.S.; Lee, J.H.; Choi, K.U.; et al. Autotaxin regulates maintenance of ovarian cancer stem cells through lysophosphatidic acid-mediated autocrine mechanism. Stem Cells 2016, 34, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Welti, R.; Wang, X. Lipid species profiling: A high-throughput approach to identify lipid compositional changes and determine the function of genes involved in lipid metabolism and signaling. Curr. Opin. Plant Biol. 2004, 7, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Bielawski, J.; Szulc, Z.M.; Hannun, Y.A.; Bielawska, A. Simultaneous quantitative analysis of bioactive sphingolipids by high-performance liquid chromatography-tandem mass spectrometry. Methods 2006, 39, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Seppanen-Laakso, T.; Oresic, M. How to study lipidomes. J. Mol. Endocrinol. 2009, 42, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Del Boccio, P.; Pieragostino, D.; di Ioia, M.; Petrucci, F.; Lugaresi, A.; De Luca, G.; Gambi, D.; Onofrj, M.; Di Ilio, C.; Sacchetta, P.; et al. Lipidomic investigations for the characterization of circulating serum lipids in multiple sclerosis. J. Proteom. 2011, 74, 2826–2836. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Han, X. Lipidomics: Techniques, applications, and outcomes related to biomedical sciences. Trends Biochem. Sci. 2016, 41, 954–969. [Google Scholar] [CrossRef] [PubMed]
- Ellis, D.I.; Dunn, W.B.; Griffin, J.L.; Allwood, J.W.; Goodacre, R. Metabolic fingerprinting as a diagnostic tool. Pharmacogenomics 2007, 8, 1243–1266. [Google Scholar] [CrossRef] [PubMed]
- Del Boccio, P.; Raimondo, F.; Pieragostino, D.; Morosi, L.; Cozzi, G.; Sacchetta, P.; Magni, F.; Pitto, M.; Urbani, A. A hyphenated microLC-Q-TOF-MS platform for exosomal lipidomics investigations: Application to RCC urinary exosomes. Electrophoresis 2012, 33, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, M.; Han, X. Applications of mass spectrometry for cellular lipid analysis. Mol. BioSyst. 2015, 11, 698–713. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.A.; Lewis, P.D.; Dunstan, P.R.; Harris, D.A. Role of Raman spectroscopy and surface enhanced Raman spectroscopy in colorectal cancer. World J. Gastrointest. Oncol. 2016, 8, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Kendall, C.; Isabelle, M.; Bazant-Hegemark, F.; Hutchings, J.; Orr, L.; Babrah, J.; Baker, R.; Stone, N. Vibrational spectroscopy: A clinical tool for cancer diagnostics. Analyst 2009, 134, 1029–1045. [Google Scholar] [CrossRef] [PubMed]
- Shim, M.G.; Song, L.M.; Marcon, N.E.; Wilson, B.C. In vivo near-infrared Raman spectroscopy: Demonstration of feasibility during clinical gastrointestinal endoscopy. Photochem. Photobiol. 2000, 72, 146–150. [Google Scholar] [CrossRef]
- Berger, A.J.; Koo, T.W.; Itzkan, I.; Horowitz, G.; Feld, M.S. Multicomponent blood analysis by near-infrared Raman spectroscopy. Appl. Opt. 1999, 38, 2916–2926. [Google Scholar] [CrossRef] [PubMed]
- Premasiri, W.R.; Clarke, R.H.; Womble, M.E. Urine analysis by laser Raman spectroscopy. Lasers Surg. Med. 2001, 28, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Pulfer, M.; Murphy, R.C. Electrospray mass spectrometry of phospholipids. Mass Spectrom. Rev. 2003, 22, 332–364. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Suss, R.; Schiller, J. An update of MALDI-TOF mass spectrometry in lipid research. Prog. Lipid Res. 2011, 49, 450–475. [Google Scholar] [CrossRef] [PubMed]
- Serna, J.; Garcia-Seisdedos, D.; Alcazar, A.; Lasuncion, M.A.; Busto, R.; Pastor, O. Quantitative lipidomic analysis of plasma and plasma lipoproteins using MALDI-TOF mass spectrometry. Chem. Phys. Lipids 2015, 189, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Pieragostino, D.; D’Alessandro, M.; di Ioia, M.; Rossi, C.; Zucchelli, M.; Urbani, A.; di Ilio, C.; Lugaresi, A.; Sacchetta, P.; Del Boccio, P. An integrated metabolomics approach for the research of new cerebrospinal fluid biomarkers of multiple sclerosis. Mol. BioSyst. 2015, 11, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network. Comprehensive molecular profiling of lung adenocarcinoma. Nature 2014, 511, 543–550. [Google Scholar]
- Soucek, L.; Whitfield, J.R.; Sodir, N.M.; Masso-Valles, D.; Serrano, E.; Karnezis, A.N.; Swigart, L.B.; Evan, G.I. Inhibition of Myc family proteins eradicates KRas-driven lung cancer in mice. Genes Dev. 2013, 27, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Hall, Z.; Ament, Z.; Wilson, C.H.; Burkhart, D.L.; Ashmore, T.; Koulman, A.; Littlewood, T.; Evan, G.I.; Griffin, J.L. Myc Expression drives aberrant lipid metabolism in lung cancer. Cancer Res. 2016, 76, 4608–4618. [Google Scholar] [CrossRef] [PubMed]
- Pirman, D.A.; Efuet, E.; Ding, X.P.; Pan, Y.; Tan, L.; Fischer, S.M.; DuBois, R.N.; Yang, P. Changes in cancer cell metabolism revealed by direct sample analysis with MALDI mass spectrometry. PLoS ONE 2013, 8, e61379. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, X.; Qiu, L.; Qin, X.; Liu, H.; Wang, Y.; Li, F.; Wang, X.; Chen, G.; Song, G.; et al. Probing gender-specific lipid metabolites and diagnostic biomarkers for lung cancer using Fourier transform ion cyclotron resonance mass spectrometry. Clin. Chim. Acta 2012, 414, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Marien, E.; Meister, M.; Muley, T.; Fieuws, S.; Bordel, S.; Derua, R.; Spraggins, J.; van de Plas, R.; Dehairs, J.; Wouters, J.; et al. Non-small cell lung cancer is characterized by dramatic changes in phospholipid profiles. Int. J. Cancer 2015, 137, 1539–1548. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Ekroos, K.; Ejsing, C.S.; Bahr, U.; Karas, M.; Simons, K.; Shevchenko, A. Charting molecular composition of phosphatidylcholines by fatty acid scanning and ion trap MS3 fragmentation. J. Lipid Res. 2003, 44, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.S.; Lee, S.C.; Park, Y.S.; Jeon, Y.E.; Lee, J.H.; Jung, S.Y.; Park, I.H.; Jang, S.H.; Park, H.M.; Yoo, C.W.; et al. Protein and lipid MALDI profiles classify breast cancers according to the intrinsic subtype. BMC Cancer 2011, 11, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Min, H.K.; Kong, G.; Moon, M.H. Quantitative analysis of urinary phospholipids found in patients with breast cancer by nanoflow liquid chromatography-tandem mass spectrometry: II. Negative ion mode analysis of four phospholipid classes. Anal. Bioanal. Chem. 2010, 396, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, M.; Iwamoto, N.; Kawaguchi-Sakita, N.; Sugimoto, M.; Ueno, T.; Mikami, Y.; Terasawa, K.; Sato, T.A.; Tanaka, K.; Shimizu, K.; et al. High-resolution imaging mass spectrometry reveals detailed spatial distribution of phosphatidylinositols in human breast cancer. Cancer Sci. 2013, 104, 1372–1379. [Google Scholar] [CrossRef] [PubMed]
- Cifkova, E.; Holcapek, M.; Lisa, M.; Vrana, D.; Melichar, B.; Student, V. Lipidomic differentiation between human kidney tumors and surrounding normal tissues using HILIC-HPLC/ESI-MS and multivariate data analysis. J. Chromatogr. B 2015, 1000, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Hilvo, M.; Denkert, C.; Lehtinen, L.; Muller, B.; Brockmoller, S.; Seppanen-Laakso, T.; Budczies, J.; Bucher, E.; Yetukuri, L.; Castillo, S.; et al. Novel theranostic opportunities offered by characterization of altered membrane lipid metabolism in breast cancer progression. Cancer Res. 2011, 71, 3236–3245. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, X.; Luan, H.; Gao, D.; Lin, S.; Cai, Z.; Liu, J.; Liu, H.; Jiang, Y. Matrix-assisted laser desorption/ionization mass spectrometry imaging of cell cultures for the lipidomic analysis of potential lipid markers in human breast cancer invasion. Rapid Commun. Mass Spectrom. 2016, 30, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Doria, M.L.; Cotrim, C.Z.; Simoes, C.; Macedo, B.; Domingues, P.; Domingues, M.R.; Helguero, L.A. Lipidomic analysis of phospholipids from human mammary epithelial and breast cancer cell lines. J. Cell. Physiol. 2013, 228, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Lee, K.M.; Kim, S.H.; Kwon, Y.J.; Chun, Y.J.; Choi, H.K. Comparative metabolic and lipidomic profiling of human breast cancer cells with different metastatic potentials. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Chughtai, K.; Jiang, L.; Greenwood, T.R.; Glunde, K.; Heeren, R.M. Mass spectrometry images acylcarnitines, phosphatidylcholines, and sphingomyelin in MDA-MB-231 breast tumor models. J. Lipid Res. 2013, 54, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Hilvo, M.; Gade, S.; Hyotylainen, T.; Nekljudova, V.; Seppanen-Laakso, T.; Sysi-Aho, M.; Untch, M.; Huober, J.; von Minckwitz, G.; Denkert, C.; et al. Monounsaturated fatty acids in serum triacylglycerols are associated with response to neoadjuvant chemotherapy in breast cancer patients. Int. J. Cancer 2014, 134, 1725–1733. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Liu, L.; Zhang, J.; Bowers, J.; Gowda, G.A.; Seeger, H.; Fehm, T.; Neubauer, H.J.; Vogel, U.; Clare, S.E.; et al. Metabolomics approach for predicting response to neoadjuvant chemotherapy for breast cancer. Mol. Oncol. 2013, 7, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics, 2010. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Min, H.K.; Lim, S.; Chung, B.C.; Moon, M.H. Shotgun lipidomics for candidate biomarkers of urinary phospholipids in prostate cancer. Anal. Bioanal. Chem. 2011, 399, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, F.; da Costa, K.A.; Wang, S.; Kruhoffer, M.; Lewin, T.M.; Orntoft, T.F.; Coleman, R.A.; Birkenkamp-Demtroder, K. Lysophosphatidylcholine acyltransferase 1 (LPCAT1) overexpression in human colorectal cancer. J. Mol. Med. 2009, 87, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Faas, F.H.; Dang, A.Q.; White, J.; Schaefer, R.; Johnson, D. Increased prostatic lysophosphatidylcholine acyltransferase activity in human prostate cancer: A marker for malignancy. J. Urol. 2001, 165, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Lawrence, T.J.; He, Z.; Pound, C.R.; Mao, J.; Bigler, S.A. The expression level of lysophosphatidylcholine acyltransferase 1 (LPCAT1) correlates to the progression of prostate cancer. Exp. Mol. Pathol. 2012, 92, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Terada, N.; Inoue, T.; Nakayama, K.; Okada, Y.; Yoshikawa, T.; Miyazaki, Y.; Uegaki, M.; Sumiyoshi, S.; Kobayashi, T.; et al. The expression profile of phosphatidylinositol in high spatial resolution imaging mass spectrometry as a potential biomarker for prostate cancer. PLoS ONE 2014, 9, e90242. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Terada, N.; Inoue, T.; Kobayashi, T.; Nakayama, K.; Okada, Y.; Yoshikawa, T.; Miyazaki, Y.; Uegaki, M.; Utsunomiya, N.; et al. Decreased expression of lysophosphatidylcholine (16:0/OH) in high resolution imaging mass spectrometry independently predicts biochemical recurrence after surgical treatment for prostate cancer. Prostate 2015, 75, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Vogel, R.; Chandra-Kuntal, K.; Glasgow, W.; Kelavkar, U. A novel three serum phospholipid panel differentiates normal individuals from those with prostate cancer. PLoS ONE 2014, 9, e88841. [Google Scholar] [CrossRef] [PubMed]
- Duscharla, D.; Bhumireddy, S.R.; Lakshetti, S.; Pospisil, H.; Murthy, P.V.; Walther, R.; Sripadi, P.; Ummanni, R. Prostate cancer associated lipid signatures in serum studied by ESI-tandem mass spectrometryas potential new biomarkers. PLoS ONE 2016, 11, e0150253. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xiao, Y.; Elson, P.; Tan, H.; Plummer, S.J.; Berk, M.; Aung, P.P.; Lavery, I.C.; Achkar, J.P.; Li, L.; et al. Plasma lysophosphatidylcholine levels: Potential biomarkers for colorectal cancer. J. Clin. Oncol. 2007, 25, 2696–2701. [Google Scholar] [CrossRef] [PubMed]
- Dobrzynska, I.; Szachowicz-Petelska, B.; Sulkowski, S.; Figaszewski, Z. Changes in electric charge and phospholipids composition in human colorectal cancer cells. Mol. Cell. Biochem. 2005, 276, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Kurabe, N.; Hayasaka, T.; Ogawa, M.; Masaki, N.; Ide, Y.; Waki, M.; Nakamura, T.; Kurachi, K.; Kahyo, T.; Shinmura, K.; et al. Accumulated phosphatidylcholine (16:0/16:1) in human colorectal cancer; possible involvement of LPCAT4. Cancer Sci. 2013, 104, 1295–1302. [Google Scholar] [CrossRef] [PubMed]
- Mirnezami, R.; Spagou, K.; Vorkas, P.A.; Lewis, M.R.; Kinross, J.; Want, E.; Shion, H.; Goldin, R.D.; Darzi, A.; Takats, Z.; et al. Chemical mapping of the colorectal cancer microenvironment via MALDI imaging mass spectrometry (MALDI-MSI) reveals novel cancer-associated field effects. Mol. Oncol. 2014, 8, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Qiu, Y.; Zou, X.; Chen, T.; Xie, G.; Cheng, Y.; Dong, T.; Zhao, L.; Feng, B.; Hu, X.; et al. Metabonomics identifies serum metabolite markers of colorectal cancer. J. Proteome Res. 2013, 12, 3000–3009. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; Patterson, N.H.; Marcinkiewicz, M.M.; Lazaris, A.; Metrakos, P.; Chaurand, P. Histology-driven data mining of lipid signatures from multiple imaging mass spectrometry analyses: Application to human colorectal cancer liver metastasis biopsies. Anal. Chem. 2013, 85, 2860–2866. [Google Scholar] [CrossRef] [PubMed]
- Coviello, G.; Tutino, V.; Notarnicola, M.; Caruso, M.G. Erythrocyte membrane fatty acids profile in colorectal cancer patients: A preliminary study. Anticancer Res. 2014, 34, 4775–4779. [Google Scholar] [PubMed]
- Zhang, Y.; Liu, Y.; Li, L.; Wei, J.; Xiong, S.; Zhao, Z. High resolution mass spectrometry coupled with multivariate data analysis revealing plasma lipidomic alteration in ovarian cancer in Asian women. Talanta 2016, 150, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Lee, A.; Park, Y.S.; Lee, S.C.; Park, S.Y.; Han, S.Y.; Kim, K.P.; Kim, Y.H.; Yoo, C.W.; Kim, H.K. Alteration in lipid and protein profiles of ovarian cancer: Similarity to breast cancer. Int. J. Gynecol. Cancer 2011, 21, 1566–1572. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Cai, Q.; Xu, Y. The Lipidomic analyses in low and highly aggressive ovarian cancer cell lines. Lipids 2016, 51, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Sutphen, R.; Xu, Y.; Wilbanks, G.D.; Fiorica, J.; Grendys, E.C., Jr.; LaPolla, J.P.; Arango, H.; Hoffman, M.S.; Martino, M.; Wakeley, K.; et al. Lysophospholipids are potential biomarkers of ovarian cancer. Cancer Epidemiol. Biomark. Prev. 2004, 13, 1185–1191. [Google Scholar]
- Meleh, M.; Pozlep, B.; Mlakar, A.; Meden-Vrtovec, H.; Zupancic-Kralj, L. Determination of serum lysophosphatidic acid as a potential biomarker for ovarian cancer. J. Chromatogr. B 2007, 858, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shen, Z.; Wiper, D.W.; Wu, M.; Morton, R.E.; Elson, P.; Kennedy, A.W.; Belinson, J.; Markman, M.; Casey, G. Lysophosphatidic acid as a potential biomarker for ovarian and other gynecologic cancers. JAMA 1998, 280, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Chen, Y.; Kennedy, A.W.; Belinson, J.; Xu, Y. Evaluation of plasma lysophospholipids for diagnostic significance using electrospray ionization mass spectrometry (ESI-MS) analyses. Ann. N. Y. Acad. Sci. 2000, 905, 242–259. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.R.; Kim, H.; Cho, S.H. Quantitative analysis of acyl-lysophosphatidic acid in plasma using negative ionization tandem mass spectrometry. J. Chromatogr. B 2003, 788, 85–92. [Google Scholar] [CrossRef]
- Sedlakova, I.; Vavrova, J.; Tosner, J.; Hanousek, L. Lysophosphatidic acid: An ovarian cancer marker. Eur. J. Gynaecol. Oncol. 2008, 29, 511–514. [Google Scholar] [PubMed]
- Sedlakova, I.; Vavrova, J.; Tosner, J.; Hanousek, L. Lysophosphatidic acid (LPA)—A perspective marker in ovarian cancer. Tumour Biol. 2011, 32, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Wang, Y.; Dou, A.; Chen, J.; Lu, X.; Cao, R.; Xu, C.; Xu, G. Study of phospholipid profile of ovarian tumor by high performance liquid chromatography-mass spectrometry. Chin. J. Chromatogr. 2011, 29, 843–850. [Google Scholar]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; DiVittore, N.A.; Young, M.M.; Jia, Z.; Xie, K.; Ritty, T.M.; Kester, M.; Fox, T.E. Altered sphingolipid metabolism in patients with metastatic pancreatic cancer. Biomolecules 2013, 3, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Sato, K.; Komachi, M.; Malchinkhuu, E.; Tobo, M.; Kimura, T.; Kuwabara, A.; Yanagita, Y.; Ikeya, T.; Tanahashi, Y.; et al. Lysophosphatidic acid (LPA) in malignant ascites stimulates motility of human pancreatic cancer cells through LPA1. J. Biol. Chem. 2004, 279, 6595–6605. [Google Scholar] [CrossRef] [PubMed]
- Komachi, M.; Sato, K.; Tobo, M.; Mogi, C.; Yamada, T.; Ohta, H.; Tomura, H.; Kimura, T.; Im, D.S.; Yanagida, K.; et al. Orally active lysophosphatidic acid receptor antagonist attenuates pancreatic cancer invasion and metastasis in vivo. Cancer Sci. 2012, 103, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Mu, G.; Zhang, L.; Zhou, W.; Zhang, J.; Yu, H. Lysophosphatidic acid stimulates activation of focal adhesion kinase and paxillin and promotes cell motility, via LPA1–3, in human pancreatic cancer. Dig. Dis. Sci. 2013, 58, 3524–3533. [Google Scholar] [CrossRef] [PubMed]
- Macasek, J.; Vecka, M.; Zak, A.; Urbanek, M.; Krechler, T.; Petruzelka, L.; Stankova, B.; Zeman, M. Plasma fatty acid composition in patients with pancreatic cancer: Correlations to clinical parameters. Nutr. Cancer 2012, 64, 946–955. [Google Scholar] [CrossRef] [PubMed]
- Zuijdgeest-van Leeuwen, S.D.; van der Heijden, M.S.; Rietveld, T.; van den Berg, J.W.; Tilanus, H.W.; Burgers, J.A.; Wilson, J.H.; Dagnelie, P.C. Fatty acid composition of plasma lipids in patients with pancreatic, lung and oesophageal cancer in comparison with healthy subjects. Clin. Nutr. 2002, 21, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Uehara, T.; Kikuchi, H.; Miyazaki, S.; Iino, I.; Setoguchi, T.; Hiramatsu, Y.; Ohta, M.; Kamiya, K.; Morita, Y.; Tanaka, H.; et al. Overexpression of lysophosphatidylcholine acyltransferase 1 and concomitant lipid alterations in gastric cancer. Ann. Surg. Oncol. 2016, 23, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Shida, D.; Kitayama, J.; Yamaguchi, H.; Yamashita, H.; Mori, K.; Watanabe, T.; Yatomi, Y.; Nagawa, H. Sphingosine 1-phosphate transactivates c-Met as well as epidermal growth factor receptor (EGFR) in human gastric cancer cells. FEBS Lett. 2004, 577, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, V.; Chin, J.L.; Izawa, J.I. Histologic variants of urothelial bladder cancer and nonurothelial histology in bladder cancer. Can. Urol. Assoc. J. 2009, 3, 193–198. [Google Scholar] [CrossRef]
- Dill, A.L.; Eberlin, L.S.; Costa, A.B.; Zheng, C.; Ifa, D.R.; Cheng, L.; Masterson, T.A.; Koch, M.O.; Vitek, O.; Cooks, R.G. Multivariate statistical identification of human bladder carcinomas using ambient ionization imaging mass spectrometry. Chemistry 2011, 17, 2897–2902. [Google Scholar] [CrossRef] [PubMed]
- Dill, A.L.; Ifa, D.R.; Manicke, N.E.; Costa, A.B.; Ramos-Vara, J.A.; Knapp, D.W.; Cooks, R.G. Lipid profiles of canine invasive transitional cell carcinoma of the urinary bladder and adjacent normal tissue by desorption electrospray ionization imaging mass spectrometry. Anal. Chem. 2009, 81, 8758–8764. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Siewert, J.R.; Ott, K. Are squamous and adenocarcinomas of the esophagus the same disease? Semin. Radiat. Oncol. 2007, 17, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.M.; Devesa, S.S.; Chow, W.H. Incidence of adenocarcinoma of the esophagus among white Americans by sex, stage, and age. J. Natl. Cancer Inst. 2008, 100, 1184–1187. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chen, Y.; Zhang, R.; Song, Y.; Cao, J.; Bi, N.; Wang, J.; He, J.; Bai, J.; Dong, L.; et al. Global and targeted metabolomics of esophageal squamous cell carcinoma discovers potential diagnostic and therapeutic biomarkers. Mol. Cell. Proteom. 2013, 12, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Peng, Y.; Li, X.; Wang, Y.; Pan, E.; Guo, W.; Pu, Y.; Yin, L. Identification of plasma metabolomic profiling for diagnosis of esophageal squamous-cell carcinoma using an UPLC/TOF/MS platform. Int. J. Mol. Sci. 2013, 14, 8899–8911. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, Y.; Hayasaka, T.; Masaki, N.; Watanabe, Y.; Masumoto, K.; Nagata, T.; Katou, F.; Setou, M. Imaging mass spectrometry distinguished the cancer and stromal regions of oral squamous cell carcinoma by visualizing phosphatidylcholine (16:0/16:1) and phosphatidylcholine (18:1/20:4). Anal. Bioanal. Chem. 2014, 406, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Moch, H.; Cubilla, A.L.; Humphrey, P.A.; Reuter, V.E.; Ulbright, T.M. The 2016 WHO classification of tumours of the urinary system and male genital organs-part A: Renal, penile, and testicular tumours. Eur. Urol. 2016, 70, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Diekstra, M.H.; Swen, J.J.; Gelderblom, H.; Guchelaar, H.J. A decade of pharmacogenomics research on tyrosine kinase inhibitors in metastatic renal cell cancer: A systematic review. Expert Rev. Mol. Diagn. 2016, 16, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Hakimi, A.A.; Reznik, E.; Lee, C.H.; Creighton, C.J.; Brannon, A.R.; Luna, A.; Aksoy, B.A.; Liu, E.M.; Shen, R.; Lee, W.; et al. An integrated metabolic atlas of clear cell renal cell carcinoma. Cancer Cell 2016, 29, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Monteiro, M.; Jeronimo, C.; Henrique, R.; Belo, L.; Bastos, M.L.; de Pinho, P.G.; Carvalho, M. Renal cell carcinoma: A critical analysis of metabolomic biomarkers emerging from current model systems. Transl. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Huang, Z.; Gao, Y.; Chen, Y.; Hang, W.; Xing, J.; Yan, X. LC-MS-based serum metabolic profiling for genitourinary cancer classification and cancer type-specific biomarker discovery. Proteomics 2012, 12, 2238–2246. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.E.; Powers, T.W.; Neely, B.A.; Cazares, L.H.; Troyer, D.A.; Parker, A.S.; Drake, R.R. MALDI imaging mass spectrometry profiling of proteins and lipids in clear cell renal cell carcinoma. Proteomics 2014, 14, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Arai, E.; Maekawa, K.; Ishikawa, M.; Fujimoto, H.; Taguchi, R.; Matsumoto, K.; Kanai, Y.; Saito, Y. Lipidomic signatures and associated transcriptomic profiles of clear cell renal cell carcinoma. Sci. Rep. 2016, 6, 28932–28944. [Google Scholar] [CrossRef] [PubMed]
- Cifkova, E.; Holcapek, M.; Lisa, M.; Vrana, D.; Gatek, J.; Melichar, B. Determination of lipidomic differences between human breast cancer and surrounding normal tissues using HILIC-HPLC/ESI-MS and multivariate data analysis. Anal. Bioanal. Chem. 2015, 407, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Dill, A.L.; Eberlin, L.S.; Zheng, C.; Costa, A.B.; Ifa, D.R.; Cheng, L.; Masterson, T.A.; Koch, M.O.; Vitek, O.; Cooks, R.G. Multivariate statistical differentiation of renal cell carcinomas based on lipidomic analysis by ambient ionization imaging mass spectrometry. Anal. Bioanal. Chem. 2010, 398, 2969–2978. [Google Scholar] [CrossRef] [PubMed]
- Jonklaas, J.; Nogueras-Gonzalez, G.; Munsell, M.; Litofsky, D.; Ain, K.B.; Bigos, S.T.; Brierley, J.D.; Cooper, D.S.; Haugen, B.R.; Ladenson, P.W.; et al. The impact of age and gender on papillary thyroid cancer survival. J. Clin. Endocrinol. Metab. 2012, 97, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Tateya, I.; Hayasaka, T.; Masaki, N.; Takizawa, Y.; Ohno, S.; Kojima, T.; Kitani, Y.; Kitamura, M.; Hirano, S.; et al. Increased expression of phosphatidylcholine (16:0/18:1) and (16:0/18:2) in thyroid papillary cancer. PLoS ONE 2012, 7, e48873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Qiu, L.; Wang, Y.; Qin, X.; Liu, H.; He, M.; Zhang, Y.; Li, Z.; Chen, X. Tissue imaging and serum lipidomic profiling for screening potential biomarkers of thyroid tumors by matrix-assisted laser desorption/ionization—Fourier transform ion cyclotron resonance mass spectrometry. Anal. Bioanal. Chem. 2014, 406, 4357–4370. [Google Scholar] [CrossRef] [PubMed]
- Mills, G.B.; Moolenaar, W.H. The emerging role of lysophosphatidic acid in cancer. Nat. Rev. Cancer 2003, 3, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Morad, S.A.; Cabot, M.C. Ceramide-orchestrated signalling in cancer cells. Nat. Rev. Cancer 2013, 13, 51–65. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Class of Lipids | Subclasses of Lipids | Structures |
|---|---|---|
| FAs | Saturated FAs, Mono-Unsaturated FAs, Poly-Unsaturated FAs (Arachidonic acid and derivatives), Hydroxy Fas |  |
| GLs | Monoacylglycerols, Diacylglycerols, Triacylglicerols (TGs) |  |
| GPLs | PC, PE, PG, PS, PI, PA, LPLs, Plasmalogens (ether phospholipids) |  |
| SLs | SM, Lyso SM, Ceramides, Cerebrosides, Gangliosides, Sulfatides |  |
| STLs | Sterols, Steroids, Steroid conjugates |  |
| PRLs | Isoprenoids, Quinones and Hidroquinones, Polyprenols |  |
| SCLs | Acylaminosugars, Acylaminosugars glycan, Acyltrehaloses |  |
| Lipid Class | Name of Lipid | Tumor Type | Sample Type | Up | Down | Diag. Factor | Prognostic Factor | Predictive Factor | Reference |
|---|---|---|---|---|---|---|---|---|---|
| GPLs | PI arachidonate-containing phospholipids | Lung cancer (adenocarcinomas) | Tissue | X | – | X | – | – | [46] |
| GPLs | PC 32:0 | Lung cancer (adenocarcinomas) | Tissue | – | X | X | – | – | [46] |
| PC 32:1 | |||||||||
| PGs | |||||||||
| FAs | free arachidonic acid | Lung cancer (adenocarcinomas) | Tissue | X | – | X | – | – | [46] |
| GPLs | PC 34:1 | Lung cancer (NSCLC) | Tissue | X* | – | X | – | – | [14] |
| PC 36:2 | |||||||||
| PC 36:3 | |||||||||
| GPLs, SLs | PC 32:0 | Lung cancer (NSCLC) | Tissue | X* | – | X | – | – | [14] |
| ST-OH 42:1 | |||||||||
| m/z 906.89 | |||||||||
| FAs | EPA | Lung cancer (NSCLC) | Tissue | X | X | – | – | X | [47] |
| GPLs | SM 16:0/1 | Lung cancer (Different cancer type) | Serum | X | – | X | – | – | [48] |
| LPC 18:1 | |||||||||
| LPC 20:4 | |||||||||
| LPC 20:3 | |||||||||
| LPC 22:6 | |||||||||
| GPLs | PI 38:3 | Lung cancer (NSCLC) | Tissue | X | – | X | – | – | [49] |
| PI 40:3 | |||||||||
| PI 38:2 | |||||||||
| SLs | SM 40:1 | Lung cancer (NSCLC) | Tissue | – | X | X | – | – | [49] |
| SM 42:1 | |||||||||
| SM 36:1 | |||||||||
| GPLs, SLs | PC | Breast cancer NS | Tissue | X | – | – | X | – | [56] |
| PE | |||||||||
| PI | |||||||||
| SMs | |||||||||
| GPLs | PC 34:1 | Breast cancer (luminal, HER2+, and triple-negative) | Tissue | X* | – | X | – | – | [52] |
| Palmitoyl carnitines, stearoyl carnitine GPLs, SLs | PC16:0/16:0 | Breast cancer (MDA-MB-231 model) | Tissue | X* | – | X | – | – | [60] |
| PC16:0/18:1 | |||||||||
| PC18:1/18:1 | |||||||||
| PC18:0/18:1 | |||||||||
| PC16:0/22:1 | |||||||||
| SMd18:1/16:0 | |||||||||
| GPLs | PI18:0/20:4 | Breast cancer NS | Urine | – | X | X | – | – | [53] |
| GPLs | PS (18:1/18:1 18:2/18:0) | Breast cancer NS | Urine | X | – | X | – | – | [53] |
| GPLs | PI 18:0/18:1 | Breast cancer NS | Tissue | X* | – | X | X | – | [54] |
| PI 18:0/20:3 | |||||||||
| GPLs | PCs | Breast cancer (mammary epithelial and breast cancer ) | Cell lines | X | X | X | X | – | [58] |
| PI 22:5/18:0 | |||||||||
| PI 18:0/18:1 | |||||||||
| GPLs | PS 18:0/20:4 | Breast cancer NS | Cell lines | X | – | – | X | X | [59] |
| PI 18:0/20:4 | |||||||||
| PC 18:0/20:4 | |||||||||
| GPLs | PIs | Breast cancer NS | Tissue | X | – | X | – | – | [115] |
| PEs | |||||||||
| PCs | |||||||||
| LPCs | |||||||||
| GLs | TGs containing C18:1 fatty acyl chains | Breast cancer NS | Serum | – | X° | – | – | X | [61] |
| FAs | linoleic acid (C18:2) | Breast cancer NS | Serum | – | X° | – | – | X | [62] |
| GPLs | PS 18:0/18:1 | Prostate cancer NS | Urine | X | – | X | – | – | [64] |
| PS 16:0/22:6 | |||||||||
| GPLs | PS 18:1/18:0 | Prostate cancer NS | Urine | – | X | X | – | – | [64] |
| PS 18:0/20:5 | |||||||||
| GPLs | PI 18:0/18:1 | Prostate cancer NS | Tissue | X* | – | X | – | – | [68] |
| PI 18:0/20:3 | |||||||||
| PI 18:0/20:2 | |||||||||
| GPLs | LPC 16:0/OH | Localized prostate cancer | Tissue | – | X* | X | – | – | [69] |
| SM d18:1/16:0 | |||||||||
| GPLs | LPC 16:0/OH | Localized prostate cancer | Tissue | – | X* | – | X | – | [69] |
| GPLs | PC 40:3 | Newly diagnosed Prostate cancer | Serum | X§ | – | X | – | – | [70] |
| PC 42:4 | |||||||||
| GPLs | PC 39:6 | Prostate cancer NS | Serum | X§ | – | X | – | – | [71] |
| FAs | FA 22:3 | ||||||||
| GPLs | LPC 18:1 | Colorectal cancer NS | Plasma | – | X | X | – | – | [72] |
| LPC 18:2 | |||||||||
| GPLs | PC/PE ratio | Colorectal cancer (pT 3 stage, various grades (G2, G3)) | Cell lines | X | – | – | X | – | [73] |
| GPLs | PC 16:0/16:1 | Colorectal cancer NS | Tissue | X | – | X | – | – | [74] |
| GPLs | PC 16:0/18:1 | Colorectal cancer NS | Tissue | X* | – | X | – | – | [75] |
| LPC 16:0 | |||||||||
| LPC 18:1 | |||||||||
| GPLs | PE 38:6 | Colorectal cancer liver metastasis | Tissue | X* | – | X | – | – | [77] |
| PE 40:4 | |||||||||
| FAs | n-3 PUFAs | Colorectal cancer NS | Red blood cell | – | X | X | – | – | [78] |
| FAs | n-6-PUFA/n-3-PUFA | Colorectal cancer NS | Red blood cell | X | – | X | – | – | [78] |
| GPLs | LPC | Ovarian cancer NS | Plasma | X | – | X | – | – | [79] |
| GPLs | PC | Ovarian cancer NS | Plasma | – | X | X | – | – | [79] |
| TG | |||||||||
| GLs | TGs 50:2 50:1 | Epithelial ovarian cancer | Cell lines | X | – | – | X | – | [81] |
| 52:2 54:4 54:3 | |||||||||
| GPLs | PC 32:3 | Ovarian cancer NS | Tissue | X | – | X | – | – | [80] |
| PC 34:1 | |||||||||
| PC 36:2 | |||||||||
| GPLs | LPA 16:0 | Ovarian cancer NS | Plasma | X | – | X | – | – | [82] |
| LPA 20:4 | |||||||||
| GPLs | LPA | Ovarian cancer and other gynecological cancers | Serum/Plasma | X | – | X | – | – | [83,84,86] |
| GPLs | LPA 16:0 | Ovarian cancer and other gynecological cancers | Plasma | X | – | X | – | – | [85] |
| LPA 18:2 | |||||||||
| LPA18:1 | |||||||||
| LPA18:0 | |||||||||
| LPI 16:0 | |||||||||
| LPI 18:0 | |||||||||
| LPI 20:4 | |||||||||
| GPLs | LPA | Benign and malignant ovarian cancer | Plasma | X | – | X | X | – | [87] |
| GPLs | LPA | Benign and malignant ovarian cancer | Plasma | X | – | X | – | – | [88] |
| GPLs | Plasmalogen phospatidylethanol, PC, plasmalogen PC, SM and LPC | Benign and malignant ovarian cancer | X | – | X | – | – | [89] | |
| SLs | Ceramides species (C16:0 and C24:1) | Metastatic pancreatic cancer | Tissue/Plasma | X | – | X | X | – | [91] |
| SLs | C18:0 | Metastatic pancreatic cancer | Tissue/Plasma | – | X | X | – | – | [91] |
| C20:0 | |||||||||
| C22:0 | |||||||||
| C24:0 | |||||||||
| C24:1 | |||||||||
| SLs | C16:0 | Metastatic pancreatic cancer | Tissue/Plasma | X | – | X | X | – | [91] |
| C20:0 | |||||||||
| C22:0 | |||||||||
| C24:0 | |||||||||
| C24:1 | |||||||||
| GPLs | LPA | Pancreatic cancer PANC-1 cells | Cell lines | X | – | X | – | – | [94] |
| FAs | MUFA | Pancreatic cancer NS | Plasma | X | – | X | – | – | [95] |
| GPLs | PC16:0/18:1 | Gastric cancer NS | Tissue | X | – | X | – | – | [97] |
| GPLs | LPC 16:0 | Gastric cancer NS | Tissue | – | X | X | – | – | [97] |
| GPLs | PS 18:0/18:1 | Bladder Cancer NS | Tissue | X | – | X | – | – | [100] |
| GPLs | PI 18:0/20:4 | Bladder cancer | Tissue | X | – | X | – | – | [100] |
| GPLs | PS 18:0/18:1 | Bladder cancer (Model of human invasive bladder cancer) | Tissue | X | – | X | – | – | [101] |
| GPLs | PG 18:1/18:1 | Bladder cancer (Model of human invasive bladder cancer) | Tissue | X | – | X | – | – | [101] |
| GPLs | PI 16:0/18:1 | Bladder cancer (Model of human invasive bladder cancer) | Tissue | X | – | X | – | – | [101] |
| GPLs | PI 18:0/18:1 | Bladder cancer (Model of human invasive bladder cancer) | Tissue | X | – | X | – | – | [101] |
| – | PS 18:1/18:1 | Bladder cancer (Model of human invasive bladder cancer) | Tissue | X | – | X | – | – | [101] |
| GPLs | Octanoylcarnitine LPC 16:1 Decanoylcarnitine | Esophageal cancer (ESCC) | Plasma | X | – | X | – | X | [105] |
| GPLs | PC 16:0/16:1 | Esophageal cancer (OSCC) | Tissue | X | – | X | – | – | [107] |
| GPLs | PC 18:1/20:4 | Esophageal cancer (OSCC) | Tissue | – | X | X | – | – | [107] |
| GPLs | PS | Esophageal cancer (ESCC) | Plasma | X | – | X | – | – | [106] |
| PA | |||||||||
| PC | |||||||||
| PI | |||||||||
| PE | |||||||||
| GLs SLs | PE (P-16:0e/0:0) ganglioside GM3 (d18:1/22:1) sphinganine C17 SMd18:0/16:1(9Z) | Kidney cancer NS | Serum | X | – | X | – | – | [112] |
| GPLs STLs Gls | PC | Kidney cancer NS | Tissue | X | – | X | – | – | [114] |
| Plasmalogens | |||||||||
| Cholesterol esters | |||||||||
| TGs | |||||||||
| GPLs | PE | Kidney cancer NS | Tissue | – | X | X | – | – | [114] |
| FAs | Unsaturated FAs | ||||||||
| GPLs | PL | Kidney cancer NS | Tissue | X | – | X | – | – | [55] |
| PE 36:1 | |||||||||
| PC 38:4 | |||||||||
| PC 36:2 | |||||||||
| PC 32:0 | |||||||||
| GPLs | PE 34:2 | Kidney cancer NS | Tissue | – | X | X | – | – | [55] |
| PE 36:4 | |||||||||
| PE 38:4 | |||||||||
| PC 34:1 | |||||||||
| PC34:2 | |||||||||
| PC 36:4 | |||||||||
| PI36:4 | |||||||||
| GPLs | PI18:0/20:4 | Kidney cancer NS | Tissue | X | – | X | – | – | [116] |
| PI22:4/18:0 | |||||||||
| PS18:0/18:1 | |||||||||
| PG18:1/18:1 | |||||||||
| FAs | FA12:0 | Kidney cancer (Human papillary renal carcinoma) | Tissue | – | X | X | – | – | [116] |
| GPLs | PC 16:0/18:1 | Thyroid cancer (Thyroid papillary cancer) | Tissue | X | – | X | – | – | [118] |
| PC 16:0/18:2 | |||||||||
| SLs | SMd18:0/16:1 | Thyroid cancer (Thyroid papillary cancer) | Tissue | X | – | X | – | – | [118] |
| GPLs | PC 34:1 | Malignant and benign thyroid cancer | Tissue/serum | X | – | X | X | – | [119] |
| PC 36:1 | |||||||||
| PC 32:0 | |||||||||
| GPLs | PA 36:62 | Malignant and benignant thyroid cancer | Tissue/serum | X | – | X | X | – | [119] |
| PA 36:3 | |||||||||
| PA 38:4 | |||||||||
| PA 38:5 | |||||||||
| PA 40:5 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrotti, F.; Rosa, C.; Cicalini, I.; Sacchetta, P.; Del Boccio, P.; Genovesi, D.; Pieragostino, D. Advances in Lipidomics for Cancer Biomarkers Discovery. Int. J. Mol. Sci. 2016, 17, 1992. https://doi.org/10.3390/ijms17121992
Perrotti F, Rosa C, Cicalini I, Sacchetta P, Del Boccio P, Genovesi D, Pieragostino D. Advances in Lipidomics for Cancer Biomarkers Discovery. International Journal of Molecular Sciences. 2016; 17(12):1992. https://doi.org/10.3390/ijms17121992
Chicago/Turabian StylePerrotti, Francesca, Consuelo Rosa, Ilaria Cicalini, Paolo Sacchetta, Piero Del Boccio, Domenico Genovesi, and Damiana Pieragostino. 2016. "Advances in Lipidomics for Cancer Biomarkers Discovery" International Journal of Molecular Sciences 17, no. 12: 1992. https://doi.org/10.3390/ijms17121992
APA StylePerrotti, F., Rosa, C., Cicalini, I., Sacchetta, P., Del Boccio, P., Genovesi, D., & Pieragostino, D. (2016). Advances in Lipidomics for Cancer Biomarkers Discovery. International Journal of Molecular Sciences, 17(12), 1992. https://doi.org/10.3390/ijms17121992