1. Introduction
With an annual incidence of approximating 13 to 23 per 100,000 persons per year [
1,
2,
3], the consequences of peripheral nerve injuries are notoriously devastating and life-altering. The morbidity and unintended negative effects (both physical and psychological) of current treatment on patients’ lives necessitates the need for continued translational research to provide alternative and potentially more efficacious treatment modalities. Peripheral nerves are comprised of myelinated and umyelinated fibers, with myelinated nerves surrounded by the specialized Schwann cells to provide insulation. Injuries to these structures are typically secondary to sudden stretch of a limb, laceration, compression, or ischemia. The primary insult is thought to result from direct forces applied while secondary injury is due to subsequent vascular ischemic damage. Anatomic redundancy in nerve length precludes injury with common tension, however, when exceeded; injury occurs [
4,
5]. Concurrent vascular injury to the vasa nervorum may lead to a compressive hematoma resulting in nerve ischemia, thereby exacerbating the primary injury.
In 1943, Seddon introduced a classification system to describe nerve injury, which included descriptions of neurapraxia, axonotmesis, and neurotmesis [
6]. In 1951, Sunderland expanded this classification of nerve injury into five categories [
7]. Axonotmesis is expanded to three grades for a total of five grades (
Table 1). Grade 4 and 5 require surgical intervention.
Although the classifications above are based on histology and correlate to specific injury models, most nerve lesions are of mixed pathology. A 6th degree nerve injury has been proposed to describe mixed pathology [
8]. Endogenous repair follows injury but is limited to 12–18 months [
8] due to loss of neuromuscular junction endplates and muscle fibrosis. Repair is not sustained thereafter. Thus, the degree of recovery is inversely proportional to the extent of the damage [
8,
9,
10]. Outside of a year from the injury, recovery is less likely and the patient may experience loss of function and chronic pain. Surgical options may serve to realign damaged nerves, through approximation of the ends or interposition of a graft; however, these procedures may be limited by the location of injury and length of the graft required, consequently providing suboptimal recovery [
2,
10,
11].
2. Endogenous Nerve Healing in Response to Injury
Following peripheral nerve injury, the distal and proximal segments to the lesion, surrounding neurons, and non-neural cells all respond. Initially, the neuronal cell body enlarges, Nissl bodies dissolve, and the nucleus migrates peripherally, inducing protein synthesis [
12]. Wallerian degeneration then proceeds, within 10 to 14 days, with fragmentation of the nerve distal to the lesion, however, this process does not occur immediately following injury. Primate studies demonstrate intact axons for days following the insult that may still function to transmit electric potentials upon stimulation [
13,
14]. Once initiated, fragmentation proceeds rapidly, completed within hours due to intrinsic proteases [
15,
16,
17]. Schwann cells and macrophages further facilitate this process.
Schwann cells serve a pivotal role in nerve repair in several aspects such as degeneration, remyelination, and axonal growth. Their role begins with activation after the injury occurs. They then regress to a more primitive phenotype while upregulating growth-related genes that include neurotrophic and transcription factors that stimulate axonal growth [
2,
18,
19]. Following axonal injury, the tip of the injured axon comes into contact with the intracellular contents of nearby Schwann cells and axons, as well as with inflamed neural tissue. Numerous trophic and growth-inducing molecules are secreted from these surrounding cells, resulting in a signaling cascade that begins with the release of ciliary neurotrophic factor by the damaged myelinated Schwann cells. Acute inflammatory changes at the site of the peripheral nerve lesion include an increase in local leukocytes that, along with the Schwann cells, help to clear the cell body of myelin debris [
2,
18,
19]. The concomitant interaction of Schwann cells and surrounding environment promotes axonal sprouting, typically at a rate of 1 to 4 mm per day [
2,
20,
21]. This endogenous repair yields a nerve with thinner myelin sheets, shorter nodal lengths, and decreased functional capacity [
22]. The importance of Schwann cells in the regeneration phase of nerve repair is further underscored by studies demonstrating the crossing of significantly fewer axons into the distal area of injury following deletion of the gene for the gamma 1 laminin chain of peripheral Schwann cells [
18,
19,
20,
21,
22]. When intervention is delayed, Schwann cells undergo apoptosis and the potential for repair diminishes.
3. Treatment for Peripheral Nerve Injury
The treatment for traumatic nerve injury is variable, ranging from conservative to surgical. Initial approaches may include early mobilization, physical therapy, and patient comfort. Alternatively, early surgical intervention may be favored to promote nerve regeneration. The timing of surgical intervention is dependent on whether the injury is open or closed. Closed injuries are often the result of stretch or crush injuries. Nerve injury occurs by demyelination, axonal loss, or a combination. When there is no clinical observation of improvement in a closed injury, an electrodiagnostic study is helpful to evaluate for neurapraxia and axonotmesis. Fourth and fifth degree injuries require surgical intervention [
23]. The EDX study can differentiate these injuries, however, it only indirectly distinguishes between 3rd and 4th degree injuries. Nerve regeneration occurs in 2 ways, either by novel axonal regrowth or by collateral sprouting. The goal of an EDX study at 6 weeks is to localize the injury and assess for degree of nerve injury as well as make a determination of whether there is early reinnervation. If there is no clinical, improvement a repeat, study at 3 months may demonstrate evidence of early reinnervation by either novel axonal regrowth or collateral sprouting. If there is no evidence of either, surgical intervention is an option [
23].
External neurolysis seeks to promote healing by isolating nerve from surrounding scarred tissue. More technical is the internal neurolysis procedure, which isolates conducting fascicles of nerves, and is recommended for axonotmetic lesions. This procedure can be advanced an additional step by end-to-end suturing of lesioned fascicles, termed split repair. If end-to-end approximation is not feasible then nerve grafting may be used to bridge the gap. This harvested nerve is interposed between the two free ends of the nerve and may be procured from several peripheral sites. Of note, algorithms for surgical intervention are described in the literature, but for the topic of this review, we will focus on another novel technique, nerve transfers, and the role Schwann cells may play in their efficacy.
Nerve transfer serves as a novel approach to rescue degenerative nerves following injury. The transfer, also known as neurotization, utilizes a nearby intact nerve and redirects it to the intact portion of the axon distal to the lesion. This technique effectively bypasses the region of the damaged nerve that may be impairing function. This procedure continues to gain traction as an alternative to nerve grafting, circumnavigating graft length limitations. The literature enumerates the potential nerve sites that can be transferred, such as the spinal accessory nerve, intercostal nerves, triceps nerve, ulnar nerve, and others [
24].
Nerve repair is predicated on a host of factors, including the participation of Schwann cells. The rate of peripheral nerve regeneration is increased in the presence of endogenous Schwann cells. Thus, these cells may also serve a purposeful role in aiding in nerve repair following nerve transfer. Schwann cells have also demonstrated an enhanced response when associated with other cells, such as olfactory ensheathing cells (OEC). The literature provides evidence that these cells promote better myelination and repair to sciatic axon injury synergistically [
25,
26]. Further potential resides in the ability to modify Schwann cells, such as bioengineering adhesion molecules [
27,
28].
A review of the literature corroborates evidence that there is practicality for the use of stem cells to promote peripheral nerve repair [
29]. Various stem cell lines have been investigated in repair during nerve grafts and conduits; however, their potential in nerve transfer must still be explored. Previous studies would suggest that Scwhann cells, and their progenitor cell line, will provide increased efficacy during nerve transfers, similarly to other treatment modalities. In this review, we will continue to define, explore the sources, discuss trials, and expand the potential beneficial role that stem cells and Schwann cells may serve in nerve transfer.
4. Sources of Schwann Cells
Neural crest cells provide the embryologic origin for Schwann cells [
30,
31]. The capacity of Schwann cells for proliferation, growth factor secretion, immune modulation, remyelination, and migration make them well suited for endeavors in neural repair, which have been reported in the literature for many years [
32,
33,
34]. Functional recovery was evidenced in one such study, in which autologous Schwann cells in tubes were used to connect sciatic nerve stumps in rats [
35]. This experimental method involving the use of transplanted Schwann cells in addition to axonal frameworks [
36,
37,
38] avoids many of the drawbacks associated with surgical repair and can be accomplished using one of three different delivery systems: (1) a hollow tube filled with cells may be placed in the injured site; (2) the cells can be set in hydrogel prior to being packed into the tube lumen; or (3) the aforementioned cell-laden hydrogel can be loaded into a tube that contains a supportive scaffolding [
39]. The first method of delivery is the most commonly studied; however, significant loss of cells is an important potential drawback [
35,
40,
41,
42]. The second [
37,
38] and third [
36] techniques are associated with both better results as well as higher levels of cost and technical complexity. The risk of culture contamination by fibroblasts, possible harm to the donor nerve, and the large quantity of cells needed all restrict the efficacy and practicality of these methods [
43] and have necessitated investigation into other sources of supportive cells.
It has been evidenced that differentiation of human NCSC into Schwann cells can be promoted with the use of synthetic matrix topography [
44]. Cell alignment can be stimulated with the use of aligned fiber matrices comprised of electrospun fibers of varying alignments and diameters. In one study by Ren et al., the NCSCs that were most likely to develop a Schwann cell fate were those that used fiber matrices with average diameters between 600 nm and 1.6 μm [
44]. Additionally, 2 weeks of predifferentiation in Schwann cell medium caused the human NCSC to be more responsive to the influence of aligned fiber topography than undifferentiated NCSC [
44]. These results imply that a combination of the specified differentiation medium and the most favorable nanofiber matrix could be a very useful way to heavily influence NCSC differentiation into a Schwann cell lineage.
Although Schwann cells may be harvested from a variety of tissues, bone marrow mesenchymal cells, which have been investigated for many years within the realm of in vitro expansion and transplantation [
45], are another potential source of support cells due to their availability, simplicity of retrieval, and associated safety. Mesenchymal stromal cells (MSC) are a type of multipotent somatic stem cell originating from a non-hematopoietic predecessor found in the bone marrow and are capable of differentiating into neural phenotypes, mesodermal cell lineages, and Schwann-like cells [
46,
47]. MSCs undergo transdifferentiation into a glial cell phenotype following exposure to suitable culture media, after which they express the proteins GFAP, p75, and S100 [
46,
47]. Cells differentiated in vitro have been shown to assist in functional and axonal recovery when coupled with artificial tubes and acellular grafts [
48,
49,
50].
In addition to bone marrow and nervous tissue, skin, adipose, and olfactory tissues provide other potential cell sources. Skin precursor cells (SKPC), which are neural crest-related precursors found in the dermis, are capable of in vitro differentiation into neural crest cell types that display similar features to Schwann cells and peripheral neurons [
51,
52]. Furthermore, NCSC can be found in the bulge region of hair and whisker follicles. These can, in turn, provide a highly available supply of transplantable cells due to their ability to differentiate into Schwann-like cells. Functional improvement in rat models has been demonstrated in studies involving both NCSC and SKPC transplantation in spinal cord and peripheral nerve injuries [
51,
53,
54,
55].
In reference to adipose tissue, a variety of intrinsic neurotrophic factors are reportedly expressed by differentiated adipose-derived Schwann cells [
56,
57]. According to Reid et al., expression of anti-apoptotic m-RNA of Bcl-2 in the dorsal root ganglia was notably elevated following the addition of nerve-derived or adipocyte-derived Schwann cells into the location of repair [
57]. Similarly, expression of pro-apoptotic m-RNA Bax and caspase-3 was reduced, resulting in some degree of neuroprotection [
57]. One comparative study that focused on the use of various cell types within a fibrin tube for the repair of the sciatic nerve found that primary Schwann cells resulted in better outcomes than adipose or bone marrow-derived mesenchymal stem cells [
58]. Nevertheless, some benefit to axonal restoration was achieved using adipose and bone marrow-derived mesenchymal stem cells when compared with the control group. Schwann cells derived from adipose tissue have also been employed in experimental models of facial palsy, with results including the electrophysiological and functional recovery of 0.8 cm facial nerve lesions in rats [
59].
Olfactory epithelium may provide yet another source of Schwann cells for use in the treatment of peripheral nerve injury. A study by Ohnishi et al. demonstrated olfactory sphere (OS) cells generated in culture from rat olfactory mucosa expressed both astrocyte and oligodendrocyte progenitor cell (OPC) markers [
60]. The OS cells differentiated into oligodendrocytes both in vitro and when they were grafted in vivo into the host tissue of injured rat spinal cords. However, when transected saphenous nerve endings were enclosed by silicone tubes containing collagen, the transplanted OS cells differentiated into Schwann cells, suggesting that the cells may have a higher affinity for a Schwann cell lineage in the context of peripheral nerve damage [
60].
More recently, induced pluripotent stem cells (iPSC) have been given more attention for their promising role in tissue engineering and cell therapies [
61,
62]. One study used NCSCs derived from human iPSCs to form a bridge across transected sciatic nerves in a rat model. At one month, the NCSC-engrafted nerve conduits demonstrated accelerated regeneration compared to the ESC group and showed advanced axonal myelination [
62]. Additionally, the cells derived from iPSCs had differentiated into Schwann cells and assimilated into the myelin sheath surrounding the axons, demonstrating the potential for the use of iPSC-derived Schwann cells in peripheral nerve injury.
When transplanted into the area of peripheral nerve damage, neural stem cells differentiate into Schwann-like cells [
18]. These cells promote nerve regeneration, release neural growth factors, and reduce the need for immunosuppressants by augmenting the compatibility of transplanted and host tissues. Additionally, functional recovery within a spinal cord injury model is enhanced by the co-culture and transplantation of Schwann cells with neural stem cells by promoting the stem cells' differentiation into neurons [
63].
5. Experimental Studies on Schwann-Like MSCs
A study of median nerve injuries in monkeys by Wakao et al. evidenced histological, electromyographical, and functional improvements by utilizing collagen nerve conduits packed with autologous bone marrow-derived MSC which were differentiated into Schwann cells in vitro [
48]. Remarkably, another investigation focusing on ulnar nerve injuries in non-human primates showed that functional recovery was also achieved with the use of allogenic acellular nerves filled with autologous bone marrow MSCs that had not been differentiated [
64]. The similarity in success rates between this study and studies using autografts and acellular nerves filled with autologous Schwann cells suggests that repair may be possible without pre-differentiation of the MSCs into Schwann cells prior to transplantation. Interestingly, an experiment involving the local transplantation of allogenic Schwann cells into a rat model of spinal cord injury resulted in the migration of endogenous Schwann cells to the injured area [
65]. While this cannot yet be translated into peripheral nerve damage, it does give rise to the hypothesis of a paracrine effect whereby recovery is supported by stimulating the infiltration of nearby host cells into the site of injury, moderating the immune response, and increasing cell survival as opposed to pure replacement. However, a study using decellularized allogeneic arteries containing Schwann cells derived from adipose tissue for the treatment of facial nerve injuries in rats found that the grafted cells lived over 8 weeks following transplantation [
66].
Schwann cells may exist in un-differentiated, differentiated, and de-differentiated states, and have the fate plasticity to transition from one state to another depending on conditions [
67,
68]. It is possible to maintain Schwann cells in a given state during culture, and indeed some literature supports the ability to influence the fate of these cells even following transplantation through such means as fibrin regulation [
69]. However, it must be noted that it is possible that the differentiation states and fate maintenance of Schwann cells in culture and following transplantation may interfere with the therapeutic effects of cell-based therapy when using the cultured cells as the source of Schwann cells.
6. Clinical Trials of Novel Therapies in Peripheral Nerve Injury
While many studies have investigated the benefits of cell therapies in animal models of peripheral nerve injury, clinical applications of these technologies in the form of randomized controlled trials continue to be elusive. One such study conducted by the University of Stanford investigated the use of scaffolding tubes containing autologous cultured Schwann cells for the repair of short and long gaps in peripheral nerve injuries. However, as of yet no clinical data from this study has been reported in the literature. One study currently recruiting participants is investigating the safety of autologous human Schwann cells in patients with subacute spinal cord injury (ClinicalTrials.gov identifier NCT01739023). A similar study is evaluating the safety of local administration of autologous bone marrow stromal cells in chronic paraplegia (ClinicalTrials.gov identifier NCT01909154). However, these studies employ cell-based therapy for the treatment of central lesions rather than focusing only on peripheral nerve injuries.
Clinical trials involving conventional nerve transfer techniques without the adjunct of cell therapies are more numerous. One such study piloted by the Washington University School of Medicine is currently enrolling patients to investigate the efficacy of nerve transfer in restoring upper extremity function following cervical spine injury (ClinicalTrials.gov identifier NCT01899664). Another study currently underway from the Lawson Health Research Institute is looking at the benefits of distal nerve transfers in the treatment of severe compressive ulnar nerve injuries (ClinicalTrials.gov identifier NCT02281656). Existing clinical data shows that nerve transfer procedures lead to functional motor and sensory recovery in several types of peripheral nerve injury. However, the application of cell therapies in combination with nerve transfer has not been studied and represents an area of potential clinical interest.
7. Building a Case for Combined Peripheral Nerve Transfer and Cell Therapy
Peripheral nerve injuries frequently produce functional disability in young and productive individuals. Spontaneous peripheral nerve regeneration is a frustratingly slow process, occurring at a rate of approximately 1 mm per day, and is a function of multiple factors including degree of injury, time, distance, scar formation, end organ atrophy, and distal nerve degeneration. Without intervention, many patients would be left with some degree of functional disability significantly affecting quality of life. As mentioned previously, there are several available methods for the treatment of peripheral nerve injury, but nerve transfer is emerging as the preferred modality in many types of nerve injury.
Nerve transfers were originally described in the setting of brachial plexus injuries and this remains the area in which they are most often utilized. In adult upper brachial plexus injuries, nerve transfers were found to be more effective at restoring elbow flexion and shoulder abduction when compared to nerve grafting [
70]. Recently, the use of nerve transfers has been widened to include repair of cervical spine injuries, ulnar and radial nerve injuries in the upper extremity, femoral nerve injuries in the lower extremity, and many more [
71,
72,
73,
74]. Current indications for nerve transfer include proximal nerve root avulsion, high peripheral nerve injuries, trauma with significant scarring at site of injury, and with large neuromas in-continuity and/or with multilevel injuries [
75]. However, as our understanding of nerve topography grows, new transfers continue to be developed and we can expect these indications to broaden in the future.
Cell-based therapy is a promising branch of regenerative medicine. The idea of using cultured Schwann cells or bone marrow mesenchymal cells in conjunction with nerve transfer procedures may be an attractive alternative to more aggressive therapies. If successful, the treatment may lead to functional improvement as well as shortened recovery times, avoiding the hurdles of additional surgeries. Although the regenerative potential and the properties of the cells change slightly from tissue to tissue, the bone marrow still stands as a preferred source of hematopoietic and mesenchymal cells due safety and established clinical use, despite greater ease of harvest with other tissue types including skin and fat [
45]. The use of allogeneic bone marrow-derived MSCs as an adjunct to nerve transfer for repair of peripheral nerve injury would be proposed as an alternative to the standard autologous nerve graft implant. The acute nature of many peripheral nerve injuries necessitates a readily available source of cells. MSCs are immunoprivileged and therefore, not readily rejected by the host, making them an ideal source for “off-the shelf” accessibility.
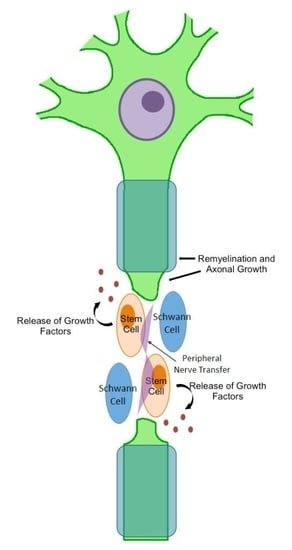
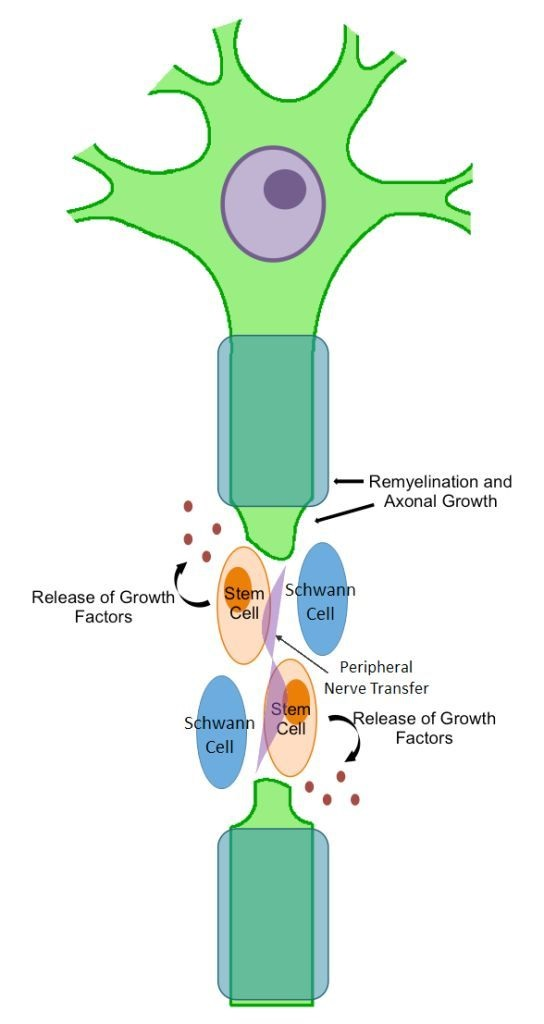
Cells would be harvested in advance, expanded in culture without further manipulation, tested for safety and quality, and then cryopreserved to ensure availability for clinical use. The necessary amount of cells would then be thawed and implanted around the neurotization site, ideally during the final stages of microsurgery. Immunosuppressive therapy would not be required, sequential functional and electrodiagnostic evaluations in the form of nerve conduction studies, and electromyography would be used to determine meaningful recovery. Either expected results would include repair and remyelination of axons as well as progressive functional improvement, through differentiation of the transplanted cells into Schwann-like cells or, more likely, through paracrine effects of the bone marrow-derived MSCs on the transferred nerve and remaining endogenous Schwann cells stimulating regeneration.
The application of MSCs to augment nerve transfer for repair of peripheral nerve injury may be associated with certain practical obstacles that would have to be navigated to ensure optimal results. The most obvious hurdle is cell viability after thawing and insertion into the surgical site. Reconstructive surgeries are often lengthy affairs and prolonged exposure of the cells to a hostile environment may reduce potential survival. Therefore, delay of cell transplantation until the final stages of the surgery, immediately followed by closure of the incision, may be critical to ensuring highest chances of cell survival. Care must be taken to avoid additional insults in the form of infection, and acidic or ischemic environments. Another concern is that MSC transplantation may lead to irregular, non-linear nerve growth patterns due to aberrant stimulation of axonal buds. This problem has been observed in nerves stimulated with growth factors, however, because of the direct end-to-end or end-to side union of donor and recipient nerve, aberrant sprouting is not expected following nerve transfer procedures augmented with MSCs. Finally, whenever stem cells are used there is a possibility that they may differentiate into lineages other than the one that is desired. Malignancy is not reported as a usual consequence of MSC administrations, but it is possible that due to the local host conditions the cells could differentiate into non-neural cell types. Therefore, clinical trials with adequate follow up would be needed to ensure the safety and efficacy of the proposed treatment.
In summary, despite a growing literature base illuminating the therapeutic benefits of cell-based therapy in experimental models of peripheral nerve injury, specific translational hurdles will need to be addressed before the realization of large clinical trials for this treatment. Moving forward, this form of cell therapy may evolve to include more advanced features such as the addition of trophic factors and manipulation of the cells for expression of neurotrophic factors or even adhesion molecules. To this end, pursuing cell therapy in combination with nerve transfer may allow improved outcomes by complementing each individual treatment’s weaknesses and strengths. Transplanted cells, via secreted trophic and paracrine factors, may aid in stimulating the regenerative capabilities of the transferred nerve. In tandem, the transferred nerve may close the physical gap of the injured peripheral nerve facilitating the survival of the transplanted cells, as well as providing the stem cells the ability to form appropriate nerve growth patterns and axonal projections.
8. Conclusions
Repair of peripheral nerve injury through spontaneous regeneration is often unsatisfactory even after microsurgical intervention, especially in the setting of severe injuries. Ensuing chronic pain and functional disability have been shown to affect victims of all ages, including those in an economically productive phase of their life. Nerve transfers are emerging as the preferred modality for treatment of several types of peripheral nerve injuries. Advantages include earlier muscle re-innervation, restoration of sensation, avoidance of surgery in scarred fields, and rare need for grafts. Despite the potential benefits, to our knowledge, there are currently no studies investigating the application of cell-based therapies as an adjunct to traditional nerve transfer procedures. Schwann cell cultures have demonstrated favorable results in the experimental models of peripheral nerve injury; however, the ideal source of cells has not yet been established. Bone marrow-derived mesenchymal cells present encouraging results and numerous advantageous properties including minimally invasive harvesting, high cell viability rates, immunoprivileged status, and the secretion of multiple trophic factors make them a possible front-runner for clinical translation.
Cultured cells, combined with adjunctive treatments such as nerve transfers may be a feasible approach for future clinical studies. The neurotropic effect of these cells at the transfer site, combined with a favorable effect over the surrounding environment, may offer shortened recovery times and improved functional restoration. Nerve transfers and cell-based therapy individually have evidence supporting their efficacy in the treatment of peripheral nerve injury; however, the marriage of these individually strong elements into one modality is a novel prospect that should be an exciting area of active research in the future.
Acknowledgments
Cesario V. Borlongan is supported by National Institutes of Health, National Institute of Neurological Disorders, and Stroke 1R01NS071956, 1R01NS090962, and 1R21NS089851, Department of Defense W81XWH-11-1-0634, James and Esther King Foundation for Biomedical Research Program, SanBio Inc., KM Pharmaceuticals, NeuralStem Inc, and Karyopharm Inc. The content is solely the responsibility of the authors and does not necessarily represent the official views of the sponsors.
Author Contributions
Naomi Abel and Cesario V. Borlongan conceptualized the topics; Robert Sullivan, Travis Dailey, Kelsey Duncan, Naomi Abel, and Cesario V. Borlongan wrote the draft and finalized the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| EDX | electrodiagnostic |
| OEC | olfactory ensheathing cells |
| NCSC | neural crest stem cells |
| MSC | mesenchymal stromal cells |
| SKPC | skin precursor cells |
| OS | olfactory sphere |
| OPC | oligodendrocyte progenitor cell |
| iPSC | induced pluripotent stem cells |
References
- Asplund, M.; Nilsson, M.; Jacobsson, A.; von Holst, H. Incidence of traumatic peripheral nerve injuries and amputations in Sweden between 1998 and 2006. Neuroepidemiology 2009, 32, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Evans, G.R. Peripheral nerve injury: A review and approach to tissue engineered constructs. Anat. Rec. 2001, 263, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.A.; Braza, D.; Rice, J.B.; Dillingham, T. The incidence of peripheral nerve injury in extremity trauma. Am. J. Phys. Med. Rehabil. Assoc. Acad. Physiatr. 2008, 87, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Grant, G.A.; Goodkin, R.; Kliot, M. Evaluation and surgical management of peripheral nerve problems. Neurosurgery 1999, 44, 825–839. [Google Scholar] [CrossRef] [PubMed]
- Sunderland, S. The anatomy and physiology of nerve injury. Muscle Nerve 1990, 13, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Seddon, H.J. Three types of nerve injury. Brain 1943, 66, 237–288. [Google Scholar] [CrossRef]
- Sunderland, S. A classification of peripheral nerve injuries producing loss of function. Brain 1951, 74, 491–516. [Google Scholar] [CrossRef] [PubMed]
- Boyd, K.U.; Nimigan, A.S.; Mackinnon, S.E. Nerve reconstruction in the hand and upper extremity. Clin. Plast. Surg. 2011, 38, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L. Traumatic injury to peripheral nerves. Muscle Nerve 2000, 23, 863–873. [Google Scholar] [CrossRef]
- Sulaiman, O.A.; Gordon, T. Effects of short- and long-term Schwann cell denervation on peripheral nerve regeneration, myelination, and size. Glia 2000, 32, 234–246. [Google Scholar] [CrossRef]
- Campbell, W.W. Evaluation and management of peripheral nerve injury. Clin. Neurophysiol. 2008, 119, 1951–1965. [Google Scholar] [CrossRef] [PubMed]
- Perrin, F.E.; Lacroix, S.; Aviles-Trigueros, M.; David, S. Involvement of monocyte chemoattractant protein-1, macrophage inflammatory protein-1α and interleukin-1β in Wallerian degeneration. Brain 2005, 128, 854–866. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, V.; Glass, J.D.; Griffin, J.W. Wallerian degeneration in peripheral nerve disease. Neurol. Clin. 1992, 10, 613–627. [Google Scholar] [PubMed]
- Gilliatt, R.W.; Hjorth, R.J. Nerve conduction during Wallerian degeneration in the baloon. J. Neurol. Neurosurg. Psychiatry 1972, 35, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Beirowski, B.; Adalbert, R.; Wagner, D.; Grumme, D.S.; Addicks, K.; Ribchester, R.R.; Coleman, M.P. The progressive nature of Wallerian degeneration in wild-type and slow Wallerian degeneration (WldS) nerves. BMC Neurosci. 2005, 6, 6–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touma, E.; Kato, S.; Fukui, K.; Koike, T. Calpain-mediated cleavage of collapsin response mediator protein (CRMP)-2 during neurite degeneration in mice. Eur. J. Neurosci. 2007, 26, 3368–3381. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Wang, J.; Kim, A.; Liu, Q.; Watts, R.; Hoopfer, E.; Mitchison, T.; Luo, L.; He, Z. Involvement of the ubiquitin-proteasome system in the early stages of Wallerian degeneration. Neuron 2003, 39, 217–225. [Google Scholar] [CrossRef]
- Dong, M.M.; Yi, T.H. Stem cell and peripheral nerve injury and repair. Facial Plast. Surg. FPS 2010, 26, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Midha, R. Practical considerations concerning the use of stem cells for peripheral nerve repair. Neurosurg. Focus 2009, 26. [Google Scholar] [CrossRef] [PubMed]
- George, E.B.; Glass, J.D.; Griffin, J.W. Axotomy-induced axonal degeneration is mediated by calcium influx through ion-specific channels. J. Neurosci. 1995, 15, 6445–6452. [Google Scholar] [PubMed]
- Stoll, G.; Jander, S.; Myers, R.R. Degeneration and regeneration of the peripheral nervous system: From Augustus Waller’s observations to neuroinflammation. J. Peripher. Nerv. Syst. 2002, 7, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, A.D.; Popovich, P.G.; Ramer, M.S. Wallerian degeneration: Gaining perspective on inflammatory events after peripheral nerve injury. J. Neuroinflamm. 2011, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Ray, W.Z.; Mackinnon, S.E. Management of nerve gaps: Autografts, allografts, nerve transfers, and end-to-side neurorrhaphy. Exp. Neurol. 2010, 223, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Rohde, R.S.; Wolfe, S.W. Nerve transfers for adult traumatic brachial plexus palsy (brachial plexus nerve transfer). HSS J. 2007, 3, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Verdu, E.; Navarro, X.; Gudino-Cabrera, G.; Rodriguez, F.J.; Ceballos, D.; Valero, A.; Nieto-Sampedro, M. Olfactory bulb ensheathing cells enhance peripheral nerve regeneration. Neuroreport 1999, 10, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Wei, L.; Liu, Y.; Oudega, M.; Jiao, S.S.; Feng, S.N.; Chen, Y.; Chen, J.M.; Li, B.C. Olfactory ensheathing cells enhance Schwann cell-mediated anatomical and functional repair after sciatic nerve injury in adult rats. Exp. Neurol. 2011, 229, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Lavdas, A.A.; Franceschini, I.; Dubois-Dalcq, M.; Matsas, R. Schwann cells genetically engineered to express PSA show enhanced migratory potential without impairment of their myelinating ability in vitro. Glia 2006, 53, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Papastefanaki, F.; Chen, J.; Lavdas, A.A.; Thomaidou, D.; Schachner, M.; Matsas, R. Grafts of Schwann cells engineered to express PSA-NCAM promote functional recovery after spinal cord injury. Brain 2007, 130, 2159–2174. [Google Scholar] [CrossRef] [PubMed]
- Hundepool, C.A.; Nijhuis, T.H.; Mohseny, B.; Selles, R.W.; Hovius, S.E. The effect of stem cells in bridging peripheral nerve defects: A meta-analysis. J. Neurosurg. 2014, 121, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Le Douarin, N.M.; Dupin, E. Multipotentiality of the neural crest. Curr. Opin. Genet. Dev. 2003, 13, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Woodhoo, A.; Sommer, L. Development of the Schwann cell lineage: From the neural crest to the myelinated nerve. Glia 2008, 56, 1481–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armati, P.J.; Mathey, E.K. An update on Schwann cell biology—Immunomodulation, neural regulation and other surprises. J. Neurol. Sci. 2013, 333, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Borlongan, C.V. Remyelinating the transected peripheral nerve by fabricated Schwann cells derived from bone marrow. Exp. Neurol. 2010, 225, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Duncan, I.D.; Aguayo, A.J.; Bunge, R.P.; Wood, P.M. Transplantation of rat Schwann cells grown in tissue culture into the mouse spinal cord. J. Neurol. Sci. 1981, 49, 241–252. [Google Scholar] [CrossRef]
- Hood, B.; Levene, H.B.; Levi, A.D. Transplantation of autologous Schwann cells for the repair of segmental peripheral nerve defects. Neurosurg. Focus 2009, 26. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Chen, Z. Fabricating autologous tissue to engineer artificial nerve. Microsurgery 2002, 22, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Evans, G.R.; Brandt, K.; Katz, S.; Chauvin, P.; Otto, L.; Bogle, M.; Wang, B.; Meszlenyi, R.K.; Lu, L.; Mikos, A.G.; et al. Bioactive poly (l-lactic acid) conduits seeded with Schwann cells for peripheral nerve regeneration. Biomaterials 2002, 23, 841–848. [Google Scholar] [CrossRef]
- Udina, E.; Rodriguez, F.J.; Verdu, E.; Espejo, M.; Gold, B.G.; Navarro, X. FK506 enhances regeneration of axons across long peripheral nerve gaps repaired with collagen guides seeded with allogeneic Schwann cells. Glia 2004, 47, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Pfister, L.A.; Papaloizos, M.; Merkle, H.P.; Gander, B. Nerve conduits and growth factor delivery in peripheral nerve repair. J. Peripher. Nerv. Syst. 2007, 12, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Ansselin, A.D.; Fink, T.; Davey, D.F. Peripheral nerve regeneration through nerve guides seeded with adult Schwann cells. Neuropathol. Appl. Neurobiol. 1997, 23, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Koshimune, M.; Takamatsu, K.; Nakatsuka, H.; Inui, K.; Yamano, Y.; Ikada, Y. Creating bioabsorbable Schwann cell coated conduits through tissue engineering. Bio-Med. Mater. Eng. 2003, 13, 223–229. [Google Scholar]
- Zhang, F.; Blain, B.; Beck, J.; Zhang, J.; Chen, Z.; Chen, Z.W.; Lineaweaver, W.C. Autogenous venous graft with one-stage prepared Schwann cells as a conduit for repair of long segmental nerve defects. J. Reconstruct. Microsurg. 2002, 18, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Niapour, A.; Karamali, F.; Karbalaie, K.; Kiani, A.; Mardani, M.; Nasr-Esfahani, M.H.; Baharvand, H. Novel method to obtain highly enriched cultures of adult rat Schwann cells. Biotechnol. Lett. 2002, 32, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.J.; Zhang, S.; Mi, R.; Liu, Q.; Zeng, X.; Rao, M.; Hoke, A.; Mao, H.Q. Enhanced differentiation of human neural crest stem cells towards the Schwann cell lineage by aligned electrospun fiber matrix. Acta Biomater. 2013, 9, 7727–7736. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Ringden, O. Immunomodulation by mesenchymal stem cells and clinical experience. J. Intern. Med. 2007, 262, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Wakao, S.; Hayashi, T.; Kitada, M.; Kohama, M.; Matsue, D.; Teramoto, N.; Ose, T.; Itokazu, Y.; Koshino, K.; Watabe, H.; et al. Long-term observation of auto-cell transplantation in non-human primate reveals safety and efficiency of bone marrow stromal cell-derived Schwann cells in peripheral nerve regeneration. Exp. Neurol. 2010, 223, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Dezawa, M.; Takahashi, I.; Esaki, M.; Takano, M.; Sawada, H. Sciatic nerve regeneration in rats induced by transplantation of in vitro differentiated bone-marrow stromal cells. Eur. J. Neurosci. 2001, 14, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Keilhoff, G.; Langnaese, K.; Wolf, G.; Fansa, H. Inhibiting effect of minocycline on the regeneration of peripheral nerves. Dev. Neurobiol. 2007, 67, 1382–1395. [Google Scholar] [CrossRef] [PubMed]
- Ladak, A.; Olson, J.; Tredget, E.E.; Gordon, T. Differentiation of mesenchymal stem cells to support peripheral nerve regeneration in a rat model. Exp. Neurol. 2011, 228, 242–252. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, I.A.; Biernaskie, J.; Toma, J.G.; Midha, R.; Miller, F.D. Skin-derived precursors generate myelinating Schwann cells for the injured and dysmyelinated nervous system. J. Neurosci. 2006, 26, 6651–6660. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.E.; Paratore, C.; Dours-Zimmermann, M.T.; Rochat, A.; Pietri, T.; Suter, U.; Zimmermann, D.R.; Dufour, S.; Thiery, J.P.; Meijer, D.; et al. Neural crest-derived cells with stem cell features can be traced back to multiple lineages in the adult skin. J. Cell Biol. 2006, 175, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Amoh, Y.; Li, L.; Yang, M.; Jiang, P.; Moossa, A.R.; Katsuoka, K.; Hoffman, R.M. Hair follicle-derived blood vessels vascularize tumors in skin and are inhibited by Doxorubicin. Cancer Res. 2005, 65, 2337–2343. [Google Scholar] [CrossRef] [PubMed]
- Biernaskie, J.; Sparling, J.S.; Liu, J.; Shannon, C.P.; Plemel, J.R.; Xie, Y.; Miller, F.D.; Tetzlaff, W. Skin-derived precursors generate myelinating Schwann cells that promote remyelination and functional recovery after contusion spinal cord injury. J. Neurosci. 2007, 27, 9545–9559. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, C.; Pluderi, M.; Colleoni, F.; Belicchi, M.; Meregalli, M.; Farini, A.; Parolini, D.; Draghi, L.; Fruguglietti, M.E.; Gavina, M.; et al. Skin-derived stem cells transplanted into resorbable guides provide functional nerve regeneration after sciatic nerve resection. Glia 2007, 55, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Kingham, P.J.; Kalbermatten, D.F.; Mahay, D.; Armstrong, S.J.; Wiberg, M.; Terenghi, G. Adipose-derived stem cells differentiate into a Schwann cell phenotype and promote neurite outgrowth in vitro. Exp. Neurol. 2007, 207, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.J.; Sun, M.; Wiberg, M.; Downes, S.; Terenghi, G.; Kingham, P.J. Nerve repair with adipose-derived stem cells protects dorsal root ganglia neurons from apoptosis. Neuroscience 2011, 199, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Di Summa, P.G.; Kingham, P.J.; Raffoul, W.; Wiberg, M.; Terenghi, G.; Kalbermatten, D.F. Adipose-derived stem cells enhance peripheral nerve regeneration. J. Plast. Reconstruct. Aesthet. Surg. 2010, 63, 1544–1552. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhou, K.; Mi, W.J.; Qiu, J.H. Repair of facial nerve defects with decellularized artery allografts containing autologous adipose-derived stem cells in a rat model. Neurosci. Lett. 2011, 499, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, Y.; Iwatsuki, K.; Shinzawa, K.; Ishihara, M.; Moriwaki, T.; Umegaki, M.; Kishima, H.; Yoshimine, T. Adult olfactory sphere cells are a source of oligodendrocyte and Schwann cell progenitors. Stem Cell Res. 2013, 11, 1178–1190. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Cox, C.S.; Li, Y. Induced pluripotent stem cells for peripheral nerve regeneration. J. Stem Cells 2011, 6, 39. [Google Scholar] [PubMed]
- Wang, A.; Tang, Z.; Park, I.H.; Zhu, Y.; Patel, S.; Daley, G.Q.; Li, S. Induced pluripotent stem cells for neural tissue engineering. Biomaterials 2011, 32, 5023–5032. [Google Scholar] [CrossRef] [PubMed]
- Niapour, A.; Karamali, F.; Nemati, S.; Taghipour, Z.; Mardani, M.; Nasr-Esfahani, M.H.; Baharvand, H. Cotransplantation of human embryonic stem cell-derived neural progenitors and schwann cells in a rat spinal cord contusion injury model elicits a distinct neurogenesis and functional recovery. Cell Transpl. 2012, 21, 827–843. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhu, Q.T.; Liu, X.L.; Xu, Y.B.; Zhu, J.K. Repair of extended peripheral nerve lesions in rhesus monkeys using acellular allogenic nerve grafts implanted with autologous mesenchymal stem cells. Exp. Neurol. 2007, 204, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.E.; Moon, L.D.; Wood, P.M.; Bunge, M.B. Labeled Schwann cell transplantation: Cell loss, host Schwann cell replacement, and strategies to enhance survival. Glia 2006, 53, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhou, K.; Mi, W.J.; Qiu, J.H. Combined use of decellularized allogeneic artery conduits with autologous transdifferentiated adipose-derived stem cells for facial nerve regeneration in rats. Biomaterials 2011, 32, 8118–8128. [Google Scholar] [CrossRef] [PubMed]
- Real, C.; Glavieux-Pardanaud, C.; Vaigot, P.; Le-Douarin, N.; Dupin, E. The instability of the neural crest phenotypes: Schwann cells can differentiate into myofibroblasts. Int. J. Dev. Biol. 2003, 49, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Keilhoff, G.; Goihl, A.; Langnäse, K.; Fansa, H.; Wolf, G. Transdifferentiation of mesenchymal stem cells into Schwann cell-like myelinating cells. Eur. J. Cell Biol. 2006, 85, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Akassoglou, K.; Yu, W.M.; Akpinar, P.; Strickland, S. Fibrin inhibits peripheral nerve remyelination by regulating Schwann cell differentiation. Neuron 2002, 33, 861–875. [Google Scholar] [CrossRef]
- Ali, Z.S.; Heuer, G.G.; Faught, R.W.; Kaneriya, S.H.; Sheikh, U.A.; Syed, I.S.; Stein, S.C.; Zager, E.L. Upper brachial plexus injury in adults: Comparative effectiveness of different repair techniques. J. Neurosurg. 2015, 122, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, J.A.; Ghizoni, M.F. Nerve transfers for elbow and finger extension reconstruction in midcervical spinal cord injuries. J. Neurosurg. 2015, 122, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.M.; Franco, M.; Tung, T.H. Motor and sensory nerve transfers in the forearm and hand. Plast. Reconstruct. Surg. 2014, 134, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.Z.; Franco, M.J.; Yee, A.; Tung, T.H.; Mackinnon, S.E.; Fox, I.K. Direct radial to ulnar nerve transfer to restore intrinsic muscle function in combined proximal median and ulnar nerve injury: Case report and surgical technique. J. Hand Surg. 2014, 39, 1358–1362. [Google Scholar] [CrossRef] [PubMed]
- Suppaphol, S.; Watcharananan, I.; Tawonsawatruk, T.; Woratanarat, P.; Sasivongsbhakdi, T.; Kawinwonggowit, V. The sensory restoration in radial nerve injury using the first branch of dorsal ulnar cutaneous nerve—A cadaveric study for the feasibility of procedure and case demonstration. J. Med. Assoc. Thail. 2014, 97, 328–332. [Google Scholar]
- Hale, H.B.; Bae, D.S.; Waters, P.M. Current concepts in the management of brachial plexus birth palsy. J. Hand Surg. 2010, 35, 322–331. [Google Scholar] [CrossRef] [PubMed]
Table 1.
Peripheral Nerve Injury Grading System.
Table 1.
Peripheral Nerve Injury Grading System.
| Grade | Description |
|---|
| Grade I | -Neurapraxia, conduction block -Focal demyelination without any axonal degradation -Secondary to mild injury such as ischemia, compression, or toxins
|
| Grade II | -Axonotmesis -The axon, in addition to the myelin sheath, is disrupted by irreversible damage -Neuronal stroma (endoneurium, perineurium, and epineurium) remains intact -Seen in crush, over-stretching, and percussion (such as blast or bullet) injuries -Nerve distal to the lesion undergoes Wallerian degeneration
|
| Grade III | |
| Grade IV | |
| Grade V | -Neurotemesis -The axon, myelin sheath, and stroma are all irreversibly damaged -Follows severe lesions such as laceration, percussion, or neurotoxins
|
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).




{kind=link}