
Intronic Polymorphisms in the CDKN2B-AS1 Gene Are Strongly Associated with the Risk of Myocardial Infarction and Coronary Artery Disease in the Saudi Population


 ,
,
Abstract
:
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. TaqMan® Single Nucleotide Polymorphisms (SNPs) Genotyping
4.3. Quality for SNP Genotyping—Sanger Sequencing
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zou, J.G.; Ma, Y.T.; Xie, X.; Yang, Y.N.; Pan, S.; Adi, D.; Liu, F.; Chen, B.D. The association between CYP1A1 genetic polymorphisms and coronary artery disease in the Uygur and Han of China. Lipids Health Dis. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Borden, W.B.; Bravata, D.M.; Dai, S.; Ford, E.S.; Fox, C.S.; et al. Executive summary. Circulation 2013, 127, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Wellcome Trust Case Control Consort. Genome-wide association study of 14,000 cases of seven common diseases and 3000 shared controls. Nature 2007, 447, 661–678. [Google Scholar]
- Samani, N.J.; Erdmann, J.; Hall, A.S.; Hengstenberg, C.; Mangino, M.; Mayer, B.; Dixon, R.J.; Meitinger, T.; Braund, P.; Wichmann, H.E.; et al. Genomewide association analysis of coronary artery disease. N. Engl. J. Med. 2007, 357, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, K.G.; Li, L.; Shen, G.Q.; Hu, Y.; Yang, Y.; MacKinlay, K.G.; Topol, E.J.; Wang, Q.K. Four SNPS on chromosome 9p21 confer risk to premature, familial CAD and MI in an American Caucasian population (GeneQuest). Ann. Hum. Genet. 2008, 72, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Durda, P.; Sabourin, J.; Lange, E.M.; Nalls, M.A.; Mychaleckyj, J.C.; Jenny, N.S.; Li, J.; Walston, J.; Harris, T.B.; Psaty, B.M.; et al. Plasma levels of soluble interleukin-2 receptor α associations with clinical cardiovascular events and genome-wide association scan. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 2246–2253. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.; Stewart, A.F. 9p21 and the genetic revolution for coronary artery disease. Clin. Chem. 2012, 58, 104–112. [Google Scholar] [CrossRef] [PubMed]
- McPherson, R.; Pertsemlidis, A.; Kavaslar, N.; Stewart, A.; Roberts, R.; Cox, D.R.; Hinds, D.A.; Pennacchio, L.A.; Tybjaerg-Hansen, A.; Folsom, A.R.; et al. A common allele on chromosome 9 associated with coronary heart disease. Science 2007, 16, 1488–1491. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, H.M.; Peden, J.F.; Lorkowski, S.; Goel, A.; Ongen, H.; Green, F.; Clarke, R.; Collins, R.; Franzosi, M.G.; Tognoni, G.; et al. Susceptibility to coronary artery disease and diabetes is encoded by distinct, tightly linked SNPs in the ANRIL locus on chromosome 9p. Hum. Mol. Gen. 2008, 17, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Szpakowicz, A.; Pepinski, W.; Waszkiewicz, E.; Maciorkowska, D.; Skawronska, M.; Niemcunowicz-Janica, A.; Milewski, R.; Dobrzycki, S.; Musial, W.J.; Kaminski, K.A. Polymorphism of 9p21. 3 locus is associated with 5-year survival in high-risk patients with myocardial infarction. PLoS ONE 2013, 8, e72333. [Google Scholar] [CrossRef] [PubMed]
- Helgadottir, A.; Thorleifsson, G.; Manolescu, A.; Gretarsdottir, S.; Blondal, T.; Jonasdottir, A.; Jonasdottir, A.; Sigurdsson, A.; Baker, A.; Palsson, A.; et al. A common variant on chromosome 9p21 affects the risk of myocardial infarction. Science 2007, 8, 1491–1493. [Google Scholar] [CrossRef] [PubMed]
- Björck, H.M.; Länne, T.; Alehagen, U.; Persson, K.; Rundkvist, L.; Hamsten, A.; Dahlström, U.; Eriksson, P. Association of genetic variation on chromosome 9p21. 3 and arterial stiffness. J. Intern. Med. 2009, 265, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Gori, F.; Specchia, C.; Pietri, S.; Crociati, L.; Barlera, S.; Franciosi, M.; Nicolucci, A.; Signorini, S.; Brambilla, P.; Franzosi, M.G. Common genetic variants on chromosome 9p21 are associated with myocardial infarction and type 2 diabetes in an Italian population. BMC Med. Genet. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Ma, J.; Qi, Q.; Hartiala, J.; Allayee, H.; Campos, H. Genetic risk score and risk of myocardial infarction in Hispanics. Circulation 2011, 123, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Saade, S.; Cazier, J.B.; Ghassibe-Sabbagh, M.; Youhanna, S.; Badro, D.A.; Kamatani, Y.; Hager, J.; Yeretzian, J.S.; El-Khazen, G.; Haber, M.; et al. Large scale association analysis identifies three susceptibility loci for coronary artery disease. PLoS ONE 2011, 6, e29427. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Li, J.M.; Sun, H.; Huang, X.Q.; Lin, K.Q.; Chu, J.Y.; Yang, Z.Q. Association between gene polymorphisms and myocardial infarction in Han Chinese of Yunnan province. Chin. J. Med. Genet. 2012, 29, 413–419. [Google Scholar]
- Gränsbo, K.; Almgren, P.; Sjögren, M.; Smith, J.G.; Engström, G.; Hedblad, B.; Melander, O. Chromosome 9p21 genetic variation explains 13% of cardiovascular disease incidence but does not improve risk prediction. J. Intern. Med. 2013, 274, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Lee, B.S.; Shin, D.J.; Park, K.W.; Shin, Y.A.; Kim, K.J.; Heo, L.; Lee, J.Y.; Kim, Y.K.; Kim, Y.J.; et al. A genome-wide association study of a coronary artery disease risk variant. J. Hum. Genet. 2013, 58, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Lövkvist, H.; Sjögren, M.; Höglund, P.; Engström, G.; Jern, C.; Olsson, S.; Smith, J.G.; Hedblad, B.; Andsberg, G.; Delavaran, H.; et al. Are 25 SNPs from the CARDIoGRAM study associated with ischaemic stroke? Eur. J. Neurol. 2013, 20, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Sakalar, C.; Gurbuz, E.; Kalay, N.; Kaya, M.G. Higher frequency of rs4977574 (the G Allele) on chromosome 9p21. 3 in patients with myocardial infarction as revealed by PCR-RFLP analysis. Tohoku J. Exp. Med. 2013, 230, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Tragante, V.; Doevendans, P.A.; Nathoe, H.M.; van der Graaf, Y.; Spiering, W.; Algra, A.; de Borst, G.J.; de Bakker, P.I.; Asselbergs, F.W. The impact of susceptibility loci for coronary artery disease on other vascular domains and recurrence risk. Eur. Heart J. 2013, 34, 2896–2904. [Google Scholar] [CrossRef] [PubMed]
- van Setten, J.; Isgum, I.; Smolonska, J.; Ripke, S.; de Jong, P.A.; Oudkerk, M.; de Koning, H.; Lammers, J.W.; Zanen, P.; Groen, H.J.; et al. Genome-wide association study of coronary and aortic calcification implicates risk loci for coronary artery disease and myocardial infarction. Atherosclerosis 2013, 228, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Hamrefors, V.; Hedblad, B.; Hindy, G.; Smith, J.G.; Almgren, P.; Engström, G.; Sjögren, M.; Gränsbo, K.; Orho-Melander, M.; Melander, O. Smoking modifies the associated increased risk of future cardiovascular disease by genetic variation on chromosome 9p21. PLoS ONE 2014, 9, e85893. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ye, H.; Hong, Q.; Xu, X.; Jiang, D.; Xu, L.; Dai, D.; Sun, J.; Gao, X.; Duan, S. Association of CDKN2BAS polymorphism rs4977574 with coronary heart disease: A case-control study and a meta-analysis. Int. J. Mol. Sci. 2014, 15, 17478–17492. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.T.; Goodarzi, M.O.; Lee, W.J.; Rotter, J.I.; Chen, Y.D.; Liang, K.W.; Lee, W.L.; Sheu, W.H. The chromosome 9p21 variant not predicting long-term cardiovascular mortality in Chinese with established coronary artery disease: An eleven-year follow-up study. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Liu, X.; Zhang, Y.; Yu, L.; Zhang, F.; Liu, L.; Cai, J.; Yang, X.; Wang, X. Genetic variants associated with myocardial infarction and the risk factors in Chinese population. PLoS ONE 2014, 9, e86332. [Google Scholar] [CrossRef] [PubMed]
- Yinko, S.L.; Thanassoulis, G.; Stark, K.D.; Tsadok, M.A.; Engert, J.C.; Pilote, L. ω-3 Fatty acids and the genetic risk of early onset acute coronary syndrome. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1234–1239. [Google Scholar] [CrossRef] [PubMed]
- Yayla, Ç.; Okyay, K.; Yılmaz, A.; Sahinarslan, A.; Seda, A.; Eyiol, A.; Bolayır, A.; Sezenöz, B.; Menevse, S.; Çengel, A. OP-195 the relationship of coronary artery disease with alleles of chromosome 9p21 in Turkish population. Am. J. Cardiol. 2015, 115, S88–S89. [Google Scholar] [CrossRef]
- Nofal, H.; Abdulmohsen, M.; Khamis, A. Incidence and causes of sudden death in a university hospital in eastern Saudi Arabia. East. Mediterr. Health 2011, 17, 665–670. [Google Scholar]
- Fedele, F.; Mancone, M.; Chilian, W.M.; Severino, P.; Canali, E.; Logan, S.; De Marchis, M.L.; Volterrani, M.; Palmirotta, R.; et al. Role of genetic polymorphisms of ion channels in the pathophysiology of coronary microvascular dysfunction and ischemic heart disease. Basic Res. Cardiol. 2013, 108, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fedele, F.; Severino, P.; Bruno, N.; Stio, R.; Caira, C.; D’Ambrosi, A.; Brasolin, B.; Ohanyan, V.; Mancone, M. Role of ion channels in coronary microcirculation: A review of the literature. Future Cardiol. 2013, 9, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Cauchi, S.; Meyre, D.; Durand, E.; Proença, C.; Marre, M.; Hadjadj, S.; Choquet, H.; De Graeve, F.; Gaget, S.; Allegaert, F.; et al. Post genome-wide association studies of novel genes associated with type 2 diabetes show gene-gene interaction and high predictive value. PLoS ONE 2008, 3, e2031. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.S.; Qaw, F.; Borgio, J.F.; Albuali, W.; Suliman, A.; Nasserullah, Z.; Al-Jarrash, S.; Al-Ali, A. Spectrum of alpha-thalassemia mutations in transfusion-dependent beta-thalassemia patients from the Eastern Province of Saudi Arabia. Hemoglobin 2013, 37, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Borgio, J.F.; AbdulAzeez, S.; Al-Nafie, A.N.; Naserullah, Z.A.; Al-Jarrash, S.; Al-Madan, M.S.; Al-Muhanna, F.; Steinberg, M.H.; Al-Ali, A.K. A novel HBA2 gene conversion in cis or trans: “alpha12 allele” in a Saudi population. Blood Cells Mol. Dis. 2014, 53, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.M.; Al Halees, Z.; Molina, C.; Paterson, R.M. Consanguinity and congenital heart disease in Saudi Arabia. Am. J. Med. Genet. 2001, 99, 8–13. [Google Scholar] [CrossRef]
- Al-Nafie, A.N.; Borgio, J.F.; AbdulAzeez, S.; Al-Suliman, A.M.; Qaw, F.S.; Naserullah, Z.A.; Al-Jarrash, S.; Al-Madan, M.S.; Al-Ali, R.A.; AlKhalifah, M.A.; et al. Co-inheritance of novel ATRX gene mutation and globin (α & β) gene mutations in transfusion dependent beta-thalassemia patients. Blood Cells Mol. Dis. 2015, 55, 27–29. [Google Scholar] [PubMed]
- Borgio, J.F. Molecular nature of alpha-globin genes in the Saudi population. Saudi Med. J. 2015, 36, 1271–1276. [Google Scholar] [CrossRef] [PubMed]
- Al-Nozha, M.M.; Arafah, M.R.; Al-Mazrou, Y.Y.; Al-Maatouq, M.A.; Khan, N.B.; Khalil, M.Z.; Al-Khadra, A.H.; Al-Marzouki, K.; Abdullah, M.A.; Al-Harthi, S.S.; et al. Coronary artery disease in Saudi Arabia. Saudi Med. J. 2004, 25, 1165–1171. [Google Scholar] [PubMed]
- Al-Sheikh, S.O.; Aljabri, B.A.; Al-Ansary, L.A.; Al-Khayal, L.A.; Al-Salman, M.M.; Al-Omran, M.A. Prevalence of and risk factors for peripheral arterial disease in Saudi Arabia. A pilot cross-sectional study. Saudi Med. J. 2007, 28, 412–414. [Google Scholar] [PubMed]
- Al-Omran, M. Atherosclerotic disease and risk factor modification in Saudi Arabia: A call to action. Vasc. Health Risk Manag. 2012, 8, 349. [Google Scholar] [CrossRef] [PubMed]
- Al-Nozha, M.M.; Al-Mazrou, Y.Y.; Arafah, M.R.; Al-Maatouq, M.A.; Khalil, M.Z.; Khan, N.B.; Al-Khadra, A.; Al-Marzouki, K.; Al-Harthi, S.S.; Abdullah, M.; Al-Shahid, M.S. Smoking in Saudi Arabia and its relation to coronary artery disease. J Saudi Heart Assoc. 2009, 2, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Sampsize. Available online: http://sampsize. sourceforge.net/iface/s3.html (accessed on 5 July 2015).
- MedCalc. Available online: http://www.medcalc.org (accessed on 1 August 2015).
- Barrett, J.C.; Fry, B.; Maller, J.D.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 15, 263–265. [Google Scholar] [CrossRef] [PubMed]
- De Long, E.R.; De Long, D.M.; Clarke-Pearson, D.L. Comparing the areas under two or more correlated receiver operating characteristic curves: A nonparametric approach. Biometrics 1988, 44, 837–845. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Characteristics * | Cases (n = 250) | Controls (n = 252) |
|---|---|---|
| Males, n (%) | 163 (65.2) | 200 (79.4) |
| Females, n (%) | 87 (34.8) | 52 (20.6) |
| Age, (years) # | 52.3 ± 13.8 | 48.1 ± 7.9 |
| BMI, (kg/m2) | 33.6 ± 11.0 | 27.9 ± 4.0 |
| Hypertension, n (%) | 174 (69.6) | 0 (0) |
| Type 2 diabetes n, (%) | 146 (58.4) | 1 (0.4) |
| Dyslipidemia | 29 (11.6) | 0 (0) |
| Unstable angina | 4 (1.6) | 0 (0) |
| SNP ID | Assoc Allele | p-HW | ObsHET | PredHET | Odds Ratio (95% CI) | Case; Control Ratio | χ2 | p Value |
|---|---|---|---|---|---|---|---|---|
| rs523096 | G | 0.2009 | 0.312 | 0.314 | 1.0369 (0.7466–1.4399) | 99:401, 80:336 | 0.047 | 0.8287 |
| rs518394 | G | 0.5137 | 0.316 | 0.314 | 1.0112 (0.7294–1.4018) | 403:97, 341:83 | 0.005 | 0.9465 |
| rs564398 | C | 0.1935 | 0.257 | 0.261 | 1.4917 (1.0345–2.1511) | 89:411, 54:372 | 4.625 | 0.0315 * |
| rs7865618 | G | 0.1822 | 0.273 | 0.272 | 1.1249 (0.7906–1.6004) | 85:415, 65:357 | 0.429 | 0.5127 |
| rs10757272 | T | 0.2009 | 0.453 | 0.484 | 1.1612 (0.8882–1.5180) | 301:197, 225:169 | 1.011 | 0.3146 |
| rs4977574 | G | 0.1745 | 0.473 | 0.486 | 1.3515 (1.0462–1.7459) | 308:192, 233:193 | 4.515 | 0.0336 * |
| rs2891168 | G | 0.0698 | 0.529 | 0.500 | 2.1908 (1.6920–2.8368) | 301:197, 167:257 | 40.619 | 1.85 × 10−10 ** |
| rs1333042 | A | 0.9668 | 0.721 | 0.485 | 2.2012 (1.6932–2.8616) | 251:249, 133:293 | 34.137 | 5.14 × 10−9 ** |
| rs2383207 | A | 0.0827 | 0.284 | 0.312 | 1.0005 (0.7165–1.3971) | 96:400, 77:321 | 0 | 0.9976 |
| rs10757278 | G | 0.3587 | 0.445 | 0.499 | 1.1438 (0.8824–1.4827) | 249:251, 196:226 | 1.031 | 0.3099 |
| rs1333048 | C | 0.1595 | 0.441 | 0.474 | 1.2095 (0.9225–1.5859) | 317:183, 232:162 | 1.897 | 0.1684 |
| rs1333049 | C | 0. 4310 | 0.444 | 0.499 | 1.1034 (0.8523–1.4283) | 246:254, 200:228 | 0.564 | 0.4526 |
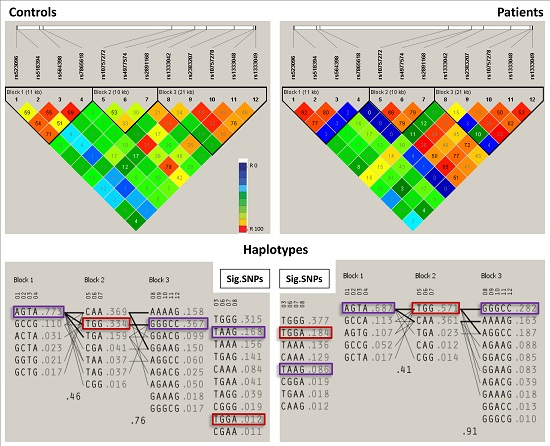
| Block | Haplotype | Frequency | Case, Control Frequency | Chi Square | p Value |
|---|---|---|---|---|---|
| Block 1 | AGTA * | 0.732 | 0.701, 0.768 | 5.245 | 0.0220 |
| Block 2 | TGG ** | 0.465 | 0.568, 0.345 | 46.38 | 9.74 × 10−12 |
| Block 3 | GGGCC * | 0.321 | 0.288, 0.360 | 5.647 | 0.0175 |
| Sig. SNPs | TAAG * | 0.147 | 0.106, 0.195 | 14.473 | 1.00 × 10−4 |
| TGGA ** | 0.147 | 0.214, 0.067 | 39.769 | 2.86 × 10−10 |
| Model | Variables | AUC (95% CI) | p Value | Difference in ACUs (95%CI): Model 2–Model 1 | p Value |
|---|---|---|---|---|---|
| Clinical variables only (Model 1) | Age, Gender, BMI | 0.79 (0.73–0.84) | 1× 10−10 | 0.08 (0.04–0.12) | 0.000236 |
| Clinical variables + SNPs (Model 2) | Age, Gender, BMI, rs1333042, rs2891168 | 0.87 (0.82–0.90) | 1× 10−10 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
AbdulAzeez, S.; Al-Nafie, A.N.; Al-Shehri, A.; Borgio, J.F.; Baranova, E.V.; Al-Madan, M.S.; Al-Ali, R.A.; Al-Muhanna, F.; Al-Ali, A.; Al-Mansori, M.; et al. Intronic Polymorphisms in the CDKN2B-AS1 Gene Are Strongly Associated with the Risk of Myocardial Infarction and Coronary Artery Disease in the Saudi Population. Int. J. Mol. Sci. 2016, 17, 395. https://doi.org/10.3390/ijms17030395
AbdulAzeez S, Al-Nafie AN, Al-Shehri A, Borgio JF, Baranova EV, Al-Madan MS, Al-Ali RA, Al-Muhanna F, Al-Ali A, Al-Mansori M, et al. Intronic Polymorphisms in the CDKN2B-AS1 Gene Are Strongly Associated with the Risk of Myocardial Infarction and Coronary Artery Disease in the Saudi Population. International Journal of Molecular Sciences. 2016; 17(3):395. https://doi.org/10.3390/ijms17030395
Chicago/Turabian StyleAbdulAzeez, Sayed, Awatif N. Al-Nafie, Abdullah Al-Shehri, J. Francis Borgio, Ekaterina V. Baranova, Mohammed S. Al-Madan, Rudaynah A. Al-Ali, Fahad Al-Muhanna, Abdullah Al-Ali, Mohammed Al-Mansori, and et al. 2016. "Intronic Polymorphisms in the CDKN2B-AS1 Gene Are Strongly Associated with the Risk of Myocardial Infarction and Coronary Artery Disease in the Saudi Population" International Journal of Molecular Sciences 17, no. 3: 395. https://doi.org/10.3390/ijms17030395
APA StyleAbdulAzeez, S., Al-Nafie, A. N., Al-Shehri, A., Borgio, J. F., Baranova, E. V., Al-Madan, M. S., Al-Ali, R. A., Al-Muhanna, F., Al-Ali, A., Al-Mansori, M., Ibrahim, M. F., Asselbergs, F. W., Keating, B., Koeleman, B. P. C., & Al-Ali, A. K. (2016). Intronic Polymorphisms in the CDKN2B-AS1 Gene Are Strongly Associated with the Risk of Myocardial Infarction and Coronary Artery Disease in the Saudi Population. International Journal of Molecular Sciences, 17(3), 395. https://doi.org/10.3390/ijms17030395