
Meta-Analysis on Associations of RGS1 and IL12A Polymorphisms with Celiac Disease Risk

Abstract
:1. Introduction
2. Results
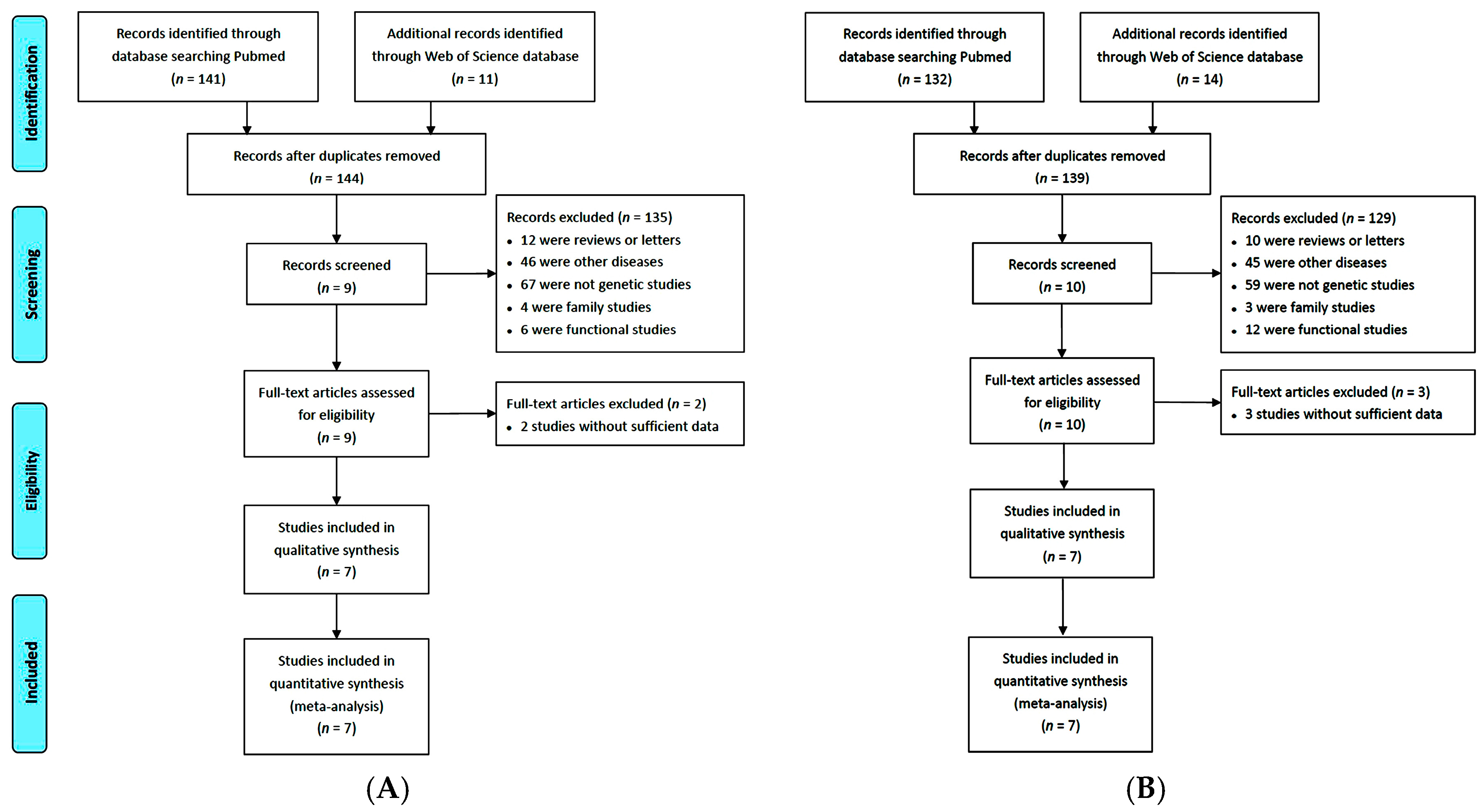
2.1. Eligible Studies and Characteristics
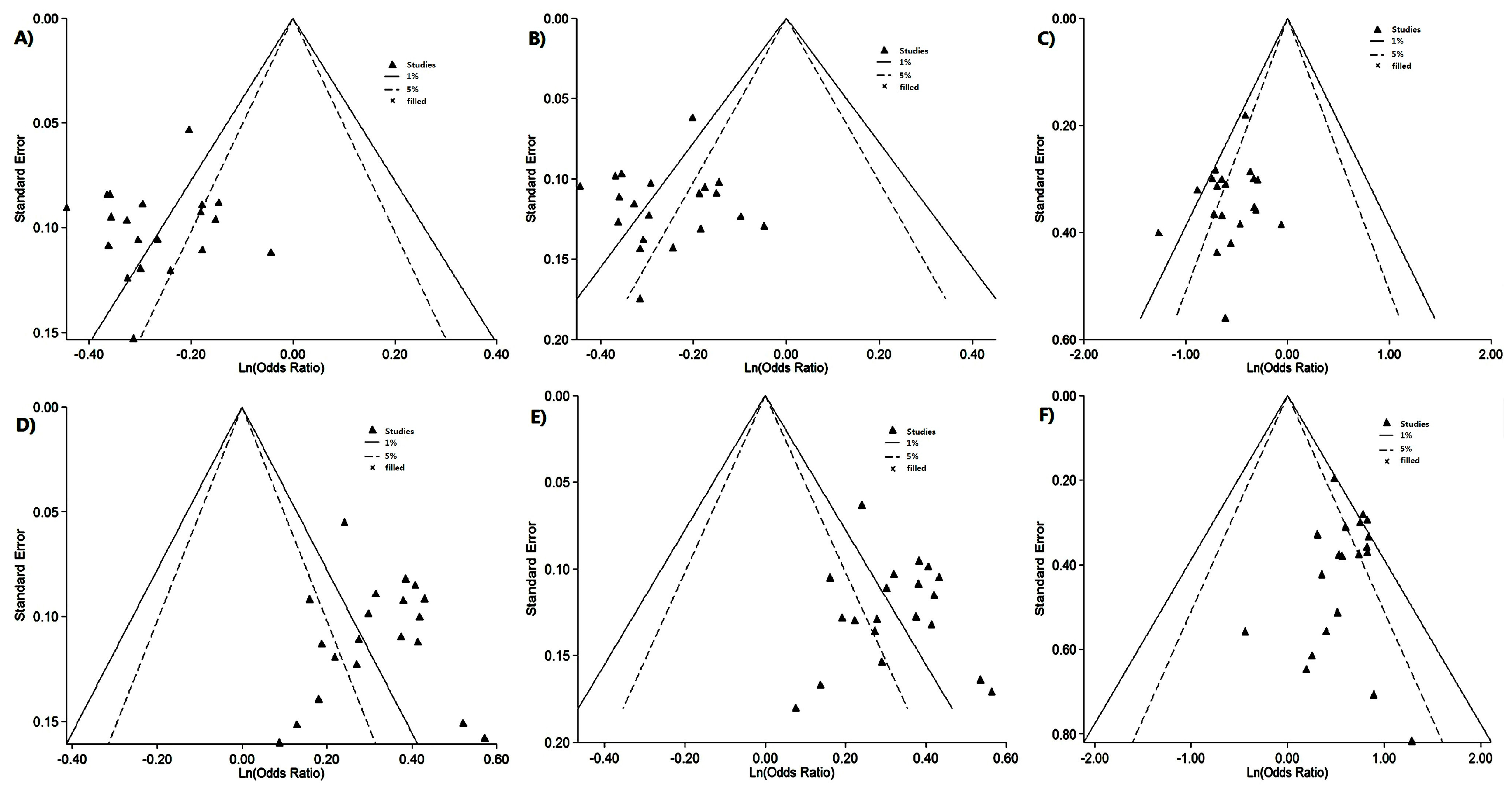
2.2. Risk of Bias Assessment
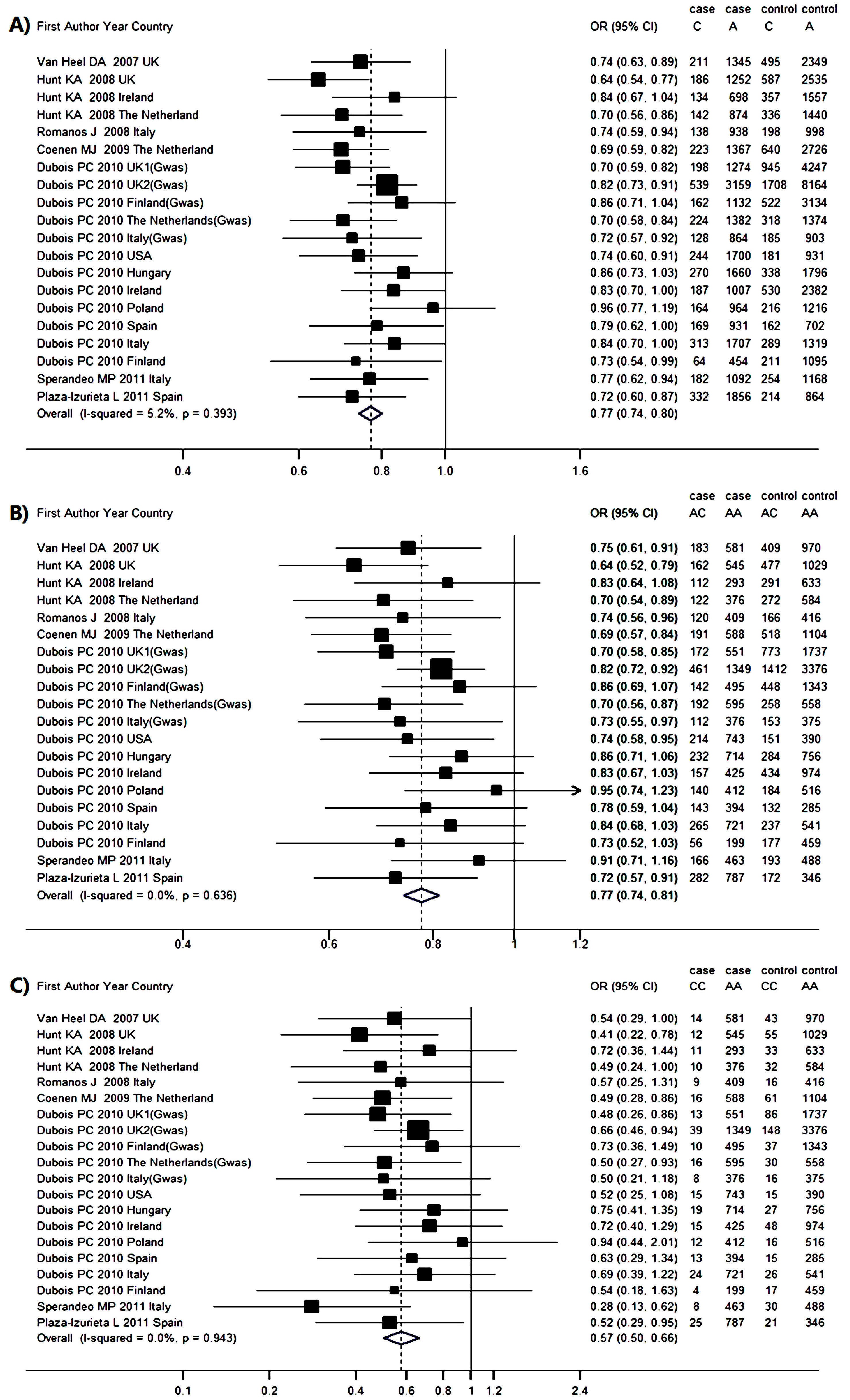
2.3. Genetic Association between RGS1 rs2816316 and CD Risk
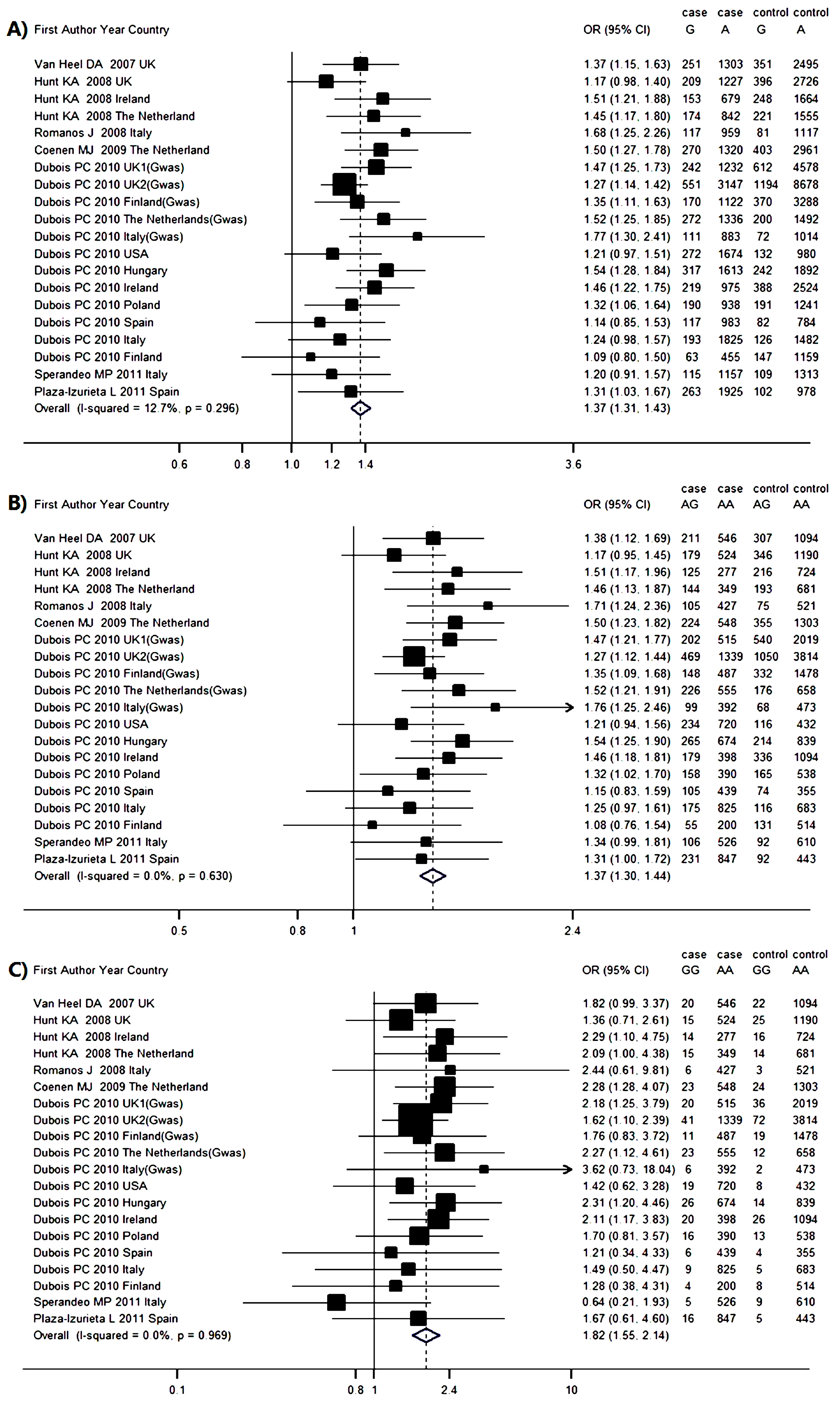
2.4. Genetic Association between IL12A rs17810546 and CD Risk
3. Discussion
4. Experimental Section
4.1. Search Strategy
4.2. Inclusion Criteria
4.3. Data Extraction
4.4. Risk of Bias Assessment
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Meresse, B.; Malamut, G.; Cerf-Bensussan, N. Celiac disease: An immunological jigsaw. Immunity 2012, 36, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.P.; Bai, J.C.; Liu, E.; Leffler, D.A. Advances in diagnosis and management of celiac disease. Gastroenterology 2015, 148, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Sollid, L.M.; Markussen, G.; Ek, J.; Gjerde, H.; Vartdal, F.; Thorsby, E. Evidence for a primary association of celiac disease to a particular HLA-DQ α/β heterodimer. J. Exp. Med. 1989, 169, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Petronzelli, F.; Bonamico, M.; Ferrante, P.; Grillo, R.; Mora, B.; Mariani, P.; Apollonio, I.; Gemme, G.; Mazzilli, M.C. Genetic contribution of the HLA region to the familial clustering of coeliac disease. Ann. Hum. Genet. 1997, 61, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Wolters, V.M.; Wijmenga, C. Genetic background of celiac disease and its clinical implications. Am. J. Gastroenterol. 2008, 103, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.A.; Zhernakova, A.; Turner, G.; Heap, G.A.; Franke, L.; Bruinenberg, M.; Romanos, J.; Dinesen, L.C.; Ryan, A.W.; Panesar, D.; et al. Newly identified genetic risk variants for celiac disease related to the immune response. Nat. Genet. 2008, 40, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Izurieta, L.; Castellanos-Rubio, A.; Irastorza, I.; Fernandez-Jimenez, N.; Gutierrez, G.; Bilbao, J.R.; CEGEC. Revisiting genome wide association studies (GWAS) in coeliac disease: Replication study in spanish population and expression analysis of candidate genes. J. Med. Genet. 2011, 48, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Han, S.B.; Moratz, C.; Huang, N.N.; Kelsall, B.; Cho, H.; Shi, C.S.; Schwartz, O.; Kehrl, J.H. Rgs1 and Gnai2 regulate the entrance of B lymphocytes into lymph nodes and B cell motility within lymph node follicles. Immunity 2005, 22, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Moratz, C.; Kang, V.H.; Druey, K.M.; Shi, C.S.; Scheschonka, A.; Murphy, P.M.; Kozasa, T.; Kehrl, J.H. Regulator of G protein signaling 1 (RGS1) markedly impairs gi alpha signaling responses of B lymphocytes. J. Immunol. 2000, 164, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Di Sabatino, A.; Corazza, G.R. Coeliac disease. Lancet 2009, 373, 1480–1493. [Google Scholar] [CrossRef]
- Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 2003, 3, 133–146. [Google Scholar] [CrossRef] [PubMed]
- International Multiple Sclerosis Genetics Conssortium. IL12A, MPHOSPH9/CDK2AP1 and RGS1 are novel multiple sclerosis susceptibility loci. Genes Immun. 2010, 11, 397–405. [Google Scholar]
- Lahdenpera, A.; Ludvigsson, J.; Falth-Magnusson, K.; Hogberg, L.; Vaarala, O. The effect of gluten-free diet on Th1-Th2-Th3-associated intestinal immune responses in celiac disease. Scand. J. Gastroenterol. 2011, 46, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Coenen, M.J.; Trynka, G.; Heskamp, S.; Franke, B.; van Diemen, C.C.; Smolonska, J.; van Leeuwen, M.; Brouwer, E.; Boezen, M.H.; Postma, D.S.; et al. Common and different genetic background for rheumatoid arthritis and coeliac disease. Hum. Mol. Genet. 2009, 18, 4195–4203. [Google Scholar] [CrossRef] [PubMed]
- Dubois, P.C.; Trynka, G.; Franke, L.; Hunt, K.A.; Romanos, J.; Curtotti, A.; Zhernakova, A.; Heap, G.A.; Adany, R.; Aromaa, A.; et al. Multiple common variants for celiac disease influencing immune gene expression. Nat. Genet. 2010, 42, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Romanos, J.; Barisani, D.; Trynka, G.; Zhernakova, A.; Bardella, M.T.; Wijmenga, C. Six new coeliac disease loci replicated in an Italian population confirm association with coeliac disease. J. Med. Genet. 2009, 46, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Sperandeo, M.P.; Tosco, A.; Izzo, V.; Tucci, F.; Troncone, R.; Auricchio, R.; Romanos, J.; Trynka, G.; Auricchio, S.; Jabri, B.; et al. Potential celiac patients: A model of celiac disease pathogenesis. PLoS ONE 2011, 6, e21281. [Google Scholar] [CrossRef] [PubMed]
- Van Heel, D.A.; Franke, L.; Hunt, K.A.; Gwilliam, R.; Zhernakova, A.; Inouye, M.; Wapenaar, M.C.; Barnardo, M.C.; Bethel, G.; Holmes, G.K.; et al. A genome-wide association study for celiac disease identifies risk variants in the region harboring IL2 and IL21. Nat. Genet. 2007, 39, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Trynka, G.; Wijmenga, C.; van Heel, D.A. A genetic perspective on coeliac disease. Trends Mol. Med. 2010, 16, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Al-Toma, A.; Verbeek, W.H.; Hadithi, M.; von Blomberg, B.M.; Mulder, C.J. Survival in refractory coeliac disease and enteropathy-associated T-cell lymphoma: Retrospective evaluation of single-centre experience. Gut 2007, 56, 1373–1378. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Xiong, K.; Issekutz, T.B. Regulation of chemokine-induced transendothelial migration of T lymphocytes by endothelial activation: Differential effects on naive and memory T cells. J. Leukoc. Biol. 2000, 67, 825–833. [Google Scholar] [PubMed]
- Smyth, D.J.; Plagnol, V.; Walker, N.M.; Cooper, J.D.; Downes, K.; Yang, J.H.; Howson, J.M.; Stevens, H.; McManus, R.; Wijmenga, C.; et al. Shared and distinct genetic variants in type 1 diabetes and celiac disease. N. Engl. J. Med. 2008, 359, 2767–2777. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, D.L.; Abeler-Dorner, L.; Raine, T.; Hwang, I.Y.; Jandke, A.; Wencker, M.; Deban, L.; Rudd, C.E.; Irving, P.M.; Kehrl, J.H.; et al. Cutting edge: Regulator of g protein signaling-1 selectively regulates gut T cell trafficking and colitic potential. J. Immunol. 2011, 187, 2067–2071. [Google Scholar] [CrossRef] [PubMed]
- Cosnes, J.; Cellier, C.; Viola, S.; Colombel, J.F.; Michaud, L.; Sarles, J.; Hugot, J.P.; Ginies, J.L.; Dabadie, A.; Mouterde, O.; et al. Incidence of autoimmune diseases in celiac disease: Protective effect of the gluten-free diet. Clin. Gastroenterol. Hepatol. 2008, 6, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Mowry, E.M.; Carey, R.F.; Blasco, M.R.; Pelletier, J.; Duquette, P.; Villoslada, P.; Malikova, I.; Roger, E.; Kinkel, R.P.; McDonald, J.; et al. Association of multiple sclerosis susceptibility variants and early attack location in the CNS. PLoS ONE 2013, 8, e75565. [Google Scholar]
- Kappen, J.H.; Medina-Gomez, C.; van Hagen, P.M.; Stolk, L.; Estrada, K.; Rivadeneira, F.; Uitterlinden, A.G.; Stanford, M.R.; Ben-Chetrit, E.; Wallace, G.R.; et al. Genome-wide association study in an admixed case series reveals IL12A as a new candidate in behcet disease. PLoS ONE 2015, 10, e0119085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Sabatino, A.; Pickard, K.M.; Gordon, J.N.; Salvati, V.; Mazzarella, G.; Beattie, R.M.; Vossenkaemper, A.; Rovedatti, L.; Leakey, N.A.; Croft, N.M.; et al. Evidence for the role of interferon-α production by dendritic cells in the Th1 response in celiac disease. Gastroenterology 2007, 133, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Paajanen, L.; Kokkonen, J.; Karttunen, T.J.; Tuure, T.; Korpela, R.; Vaarala, O. Intestinal cytokine mRNA expression in delayed-type cow’s milk allergy. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Thakkinstian, A.; McKay, G.J.; McEvoy, M.; Chakravarthy, U.; Chakrabarti, S.; Silvestri, G.; Kaur, I.; Li, X.; Attia, J. Systematic review and meta-analysis of the association between complement component 3 and age-related macular degeneration: A huge review and meta-analysis. Am. J. Epidemiol. 2011, 173, 1365–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.P.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossman, M.D.; Thompson, B.; Frederick, M.; Maliarik, M.; Iannuzzi, M.C.; Rybicki, B.A.; Pandey, J.P.; Newman, L.S.; Magira, E.; Beznik-Cizman, B.; et al. HLA-DRB1*1101: A significant risk factor for sarcoidosis in blacks and whites. Am. J. Hum. Genet. 2003, 73, 720–735. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.L.; Sutton, A.J.; Jones, D.R.; Abrams, K.R.; Rushton, L. Contour-enhanced meta-analysis funnel plots help distinguish publication bias from other causes of asymmetry. J. Clin. Epidemiol. 2008, 61, 991–996. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Year | Country | Ethnicity | Genotyping Method | MAF | Sample Size | ||
|---|---|---|---|---|---|---|---|---|
| Control_1 | Control_2 | Case | Control | |||||
| Van Heel [18] | 2007 | UK | European | The Illumina Infinium II assay | 0.174 | 0.123 | 778 | 1422 |
| Hunt [6] | 2008 | UK | European | Infinium assay | 0.188 | 0.127 | 719 | 1561 |
| – | – | Ireland | European | GoldenGate assay | 0.187 | 0.130 | 416 | 957 |
| – | – | Netherlands | European | GoldenGate assay | 0.189 | 0.124 | 508 | 888 |
| Romanos [16] | 2009 | Italy | European | Taqman probe | 0.166 | 0.067 | 538 | 598 |
| Coenen [14] | 2009 | Netherlands | European | Taqman probe | 0.190 | 0.120 | 795 | 1683 |
| Dubois [15] | 2010 | UK1 | European | Illumina Hap300v1-1 | 0.182 | 0.118 | 737 | 2596 |
| – | – | UK2 | European | Illumina 670-QuadCustom_v1 | 0.173 | 0.121 | 1849 | 4936 |
| – | – | Finland | European | Illumina 670-QuadCustom_v1 | 0.143 | 0.101 | 647 | 1829 |
| – | – | Netherlands | European | Illumina 670-QuadCustom_v1 | 0.188 | 0.118 | 803 | 846 |
| – | – | Italy | European | Illumina 670-QuadCustom_v1 | 0.169 | 0.067 | 497 | 543 |
| – | – | USA | American | Illumina GoldenGate | 0.162 | 0.118 | 973 | 555 |
| – | – | Hungary | European | Illumina GoldenGate | 0.158 | 0.113 | 965 | 1067 |
| – | – | Ireland | European | Illumina GoldenGate | 0.182 | 0.133 | 597 | 1456 |
| – | – | Poland | European | Illumina GoldenGate | 0.151 | 0.133 | 564 | 716 |
| – | – | Spain | European | Illumina GoldenGate | 0.188 | 0.095 | 550 | 433 |
| – | – | Italy | European | Illumina GoldenGate | 0.180 | 0.078 | 1010 | 804 |
| – | – | Finland | European | Illumina GoldenGate | 0.162 | 0.113 | 259 | 653 |
| Sperandeo [17] | 2011 | Italy | European | Taqman probe | 0.178 | 0.077 | 637 | 711 |
| Plaza-Izurieta [7] | 2011 | Spain | European | TaqMan probe | 0.199 | 0.094 | 1094 | 540 |
| Author and Year | CD Group | Control Group | C vs. A | AC vs. AA | CC vs. AA | HWE p Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AA | AC | CC | AA | AC | CC | OR | 95% CI | OR | 95% CI | OR | 95% CI | ||
| Van Heel 2007 UK [18] | 581 | 183 | 14 | 970 | 409 | 43 | 0.74 | 0.63–0.89 | 0.75 | 0.61–0.91 | 0.54 | 0.29–1.00 | 0.988 |
| Hunt 2008 UK [6] | 545 | 162 | 12 | 1029 | 477 | 55 | 0.64 | 0.54–0.77 | 0.64 | 0.52–0.79 | 0.41 | 0.22–0.78 | 0.976 |
| Hunt 2008 Ireland [6] | 293 | 112 | 11 | 633 | 291 | 33 | 0.84 | 0.67–1.04 | 0.83 | 0.64–1.08 | 0.72 | 0.36–1.44 | 0.950 |
| Hunt 2008 Netherlands [6] | 376 | 122 | 10 | 584 | 272 | 32 | 0.70 | 0.56–0.86 | 0.70 | 0.54–0.89 | 0.49 | 0.24–1.00 | 0.962 |
| Romanos 2008 Italy [16] | 409 | 120 | 9 | 416 | 166 | 16 | 0.74 | 0.59–0.94 | 0.74 | 0.56–0.96 | 0.57 | 0.25–1.31 | 0.908 |
| Coenen 2009 Netherlands [14] | 588 | 191 | 16 | 1104 | 518 | 61 | 0.69 | 0.59–0.94 | 0.69 | 0.57–0.84 | 0.49 | 0.28–0.86 | 0.980 |
| Dubois 2010 UK1 [15] | 551 | 172 | 13 | 1737 | 773 | 86 | 0.70 | 0.59–0.82 | 0.70 | 0.58–0.85 | 0.48 | 0.26–0.86 | 0.999 |
| Dubois 2010 UK2 [15] | 1349 | 461 | 39 | 3376 | 1412 | 148 | 0.82 | 0.73–0.91 | 0.82 | 0.72–0.92 | 0.66 | 0.46–0.94 | 0.980 |
| Dubois 2010 Finland [15] | 495 | 142 | 10 | 1343 | 448 | 37 | 0.86 | 0.71–1.04 | 0.86 | 0.69–1.07 | 0.73 | 0.36–1.49 | 0.956 |
| Dubois 2010 Netherlands [15] | 595 | 192 | 16 | 558 | 258 | 30 | 0.70 | 0.58–0.84 | 0.70 | 0.56–0.87 | 0.50 | 0.27–0.93 | 0.978 |
| Dubois 2010 Italy [15] | 376 | 112 | 8 | 375 | 153 | 16 | 0.72 | 0.57–0.92 | 0.73 | 0.55–0.97 | 0.50 | 0.21–1.18 | 0.934 |
| Dubois 2010 USA [15] | 743 | 214 | 15 | 390 | 151 | 15 | 0.74 | 0.60–1.90 | 0.74 | 0.58–0.95 | 0.52 | 0.25–1.08 | 9.933 |
| Dubois 2010 Hungary [15] | 714 | 232 | 19 | 756 | 284 | 27 | 0.86 | 0.73–1.03 | 0.86 | 0.71–1.06 | 0.75 | 0.41–1.35 | 0.957 |
| Dubois 2010 Ireland [15] | 425 | 157 | 15 | 974 | 434 | 48 | 0.83 | 0.70–1.00 | 0.83 | 0.67–1.03 | 0.72 | 0.40–1.29 | 0.967 |
| Dubois 2010 Poland [15] | 412 | 140 | 12 | 516 | 184 | 16 | 0.96 | 0.77–1.19 | 0.95 | 0.74–1.23 | 0.94 | 0.44–2.01 | 0.932 |
| Dubois 2010 Spain [15] | 394 | 143 | 13 | 285 | 132 | 15 | 0.79 | 0.62–1.00 | 0.78 | 0.59–1.04 | 0.63 | 0.29–1.34 | 0.952 |
| Dubois 2010 Italy [15] | 721 | 265 | 24 | 541 | 237 | 26 | 0.84 | 0.70–1.00 | 0.84 | 0.68–1.03 | 0.69 | 0.39–1.22 | 0.994 |
| Dubois 2010 Finland [15] | 199 | 56 | 4 | 459 | 177 | 17 | 0.73 | 0.54–0.99 | 0.73 | 0.52–1.03 | 0.54 | 0.18–1.63 | 0.989 |
| Sperandeo 2011 Italy [17] | 463 | 166 | 8 | 488 | 193 | 30 | 0.77 | 0.62–0.94 | 0.91 | 0.71–1.16 | 0.28 | 0.13–0.62 | 0.054 |
| Plaza-Izurieta 2011 Spain [7] | 787 | 282 | 25 | 346 | 172 | 21 | 0.72 | 0.60–0.87 | 0.72 | 0.57–0.91 | 0.52 | 0.29–0.95 | 0.947 |
| Overall OR | – | – | – | – | – | – | 0.77 | 0.74–0.80 | 0.77 | 0.74–0.81 | 0.57 | 0.50–0.67 | – |
| Author and Year | CD Group | Control Group | G vs. A | AG vs. AA | GG vs. AA | HWE p Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AA | AG | GG | AA | AG | GG | OR | 95% CI | OR | 95% CI | OR | 95% CI | ||
| Van Heel 2007 UK [18] | 546 | 211 | 20 | 1094 | 307 | 22 | 1.37 | 1.15–1.63 | 1.38 | 1.12–1.69 | 1.82 | 0.99–3.37 | 0.930 |
| Hunt 2008 UK [6] | 524 | 179 | 15 | 1190 | 346 | 25 | 1.17 | 0.98–1.40 | 1.17 | 0.95–1.45 | 1.36 | 0.71–2.61 | 0.979 |
| Hunt 2008 Ireland [6] | 277 | 125 | 14 | 724 | 216 | 16 | 1.51 | 1.21–1.88 | 1.51 | 1.17–1.96 | 2.29 | 1.10–4.75 | 0.980 |
| Hunt 2008 Netherlands [6] | 349 | 144 | 15 | 681 | 193 | 14 | 1.45 | 1.17–1.80 | 1.46 | 1.13–1.87 | 2.09 | 1.00–4.38 | 0.938 |
| Romanos 2008 Italy [16] | 427 | 105 | 6 | 521 | 75 | 3 | 1.68 | 1.25–2.26 | 1.71 | 1.24–2.36 | 2.44 | 0.61–9.81 | 0.865 |
| Coenen 2009 Netherlands [14] | 548 | 224 | 23 | 1303 | 355 | 24 | 1.50 | 1.27–1.78 | 1.50 | 1.23–1.82 | 2.28 | 1.28–4.07 | 0.974 |
| Dubois 2010 UK1 [15] | 515 | 202 | 20 | 2019 | 540 | 36 | 1.47 | 1.25–1.85 | 1.47 | 1.21–1.77 | 2.18 | 1.25–3.79 | 0.987 |
| Dubois 2010 UK2 [15] | 1339 | 469 | 41 | 3814 | 1050 | 72 | 1.27 | 1.14–1.42 | 1.27 | 1.12–1.44 | 1.62 | 1.10–2.39 | 0.977 |
| Dubois 2010 Finland [15] | 487 | 148 | 11 | 1478 | 332 | 19 | 1.35 | 1.11–1.63 | 1.35 | 1.09–1.68 | 1.76 | 0.83–3.72 | 0.941 |
| Dubois 2010 Netherlands [15] | 555 | 226 | 23 | 658 | 176 | 12 | 1.52 | 1.25–1.73 | 1.52 | 1.21–1.91 | 2.27 | 1.12–4.61 | 0.952 |
| Dubois 2010 Italy [15] | 392 | 99 | 6 | 473 | 68 | 2 | 1.77 | 1.30–2.41 | 1.76 | 1.25–2.46 | 3.62 | 0.73–18.04 | 0.788 |
| Dubois 2010 USA [15] | 720 | 234 | 19 | 432 | 116 | 8 | 1.21 | 0.97–1.51 | 1.21 | 0.94–1.56 | 1.42 | 0.62–3.28 | 0.946 |
| Dubois 2010 Hungary [15] | 674 | 265 | 26 | 839 | 214 | 14 | 1.54 | 1.28–1.84 | 1.54 | 1.25–2.45 | 2.31 | 1.20–4.46 | 0.932 |
| Dubois 2010 Ireland [15] | 398 | 179 | 20 | 1094 | 336 | 26 | 1.46 | 1.22–1.75 | 1.46 | 1.18–1.81 | 2.11 | 1.17–3.83 | 0.972 |
| Dubois 2010 Poland [15] | 390 | 158 | 16 | 538 | 165 | 13 | 1.32 | 1.06–1.64 | 1.32 | 1.02–1.70 | 1.70 | 0.81–3.57 | 0.932 |
| Dubois 2010 Spain [15] | 439 | 105 | 6 | 355 | 74 | 4 | 1.14 | 0.85–1.53 | 1.15 | 0.83–1.59 | 1.21 | 0.34–4.33 | 0.947 |
| Dubois 2010 Italy [15] | 825 | 175 | 9 | 683 | 116 | 5 | 1.24 | 0.98–1.57 | 1.25 | 0.97–1.61 | 1.49 | 0.50–4.47 | 0.975 |
| Dubois 2010 Finland [15] | 200 | 55 | 4 | 514 | 131 | 8 | 1.09 | 0.80–1.50 | 1.08 | 0.76–1.54 | 1.28 | 0.38–4.31 | 0.914 |
| Sperandeo 2011 Italy [17] | 526 | 106 | 5 | 610 | 92 | 9 | 1.20 | 0.91–1.57 | 1.34 | 0.99–1.81 | 0.64 | 0.21–1.93 | 0.029 |
| Plaza-Izurieta 2011 Spain [7] | 847 | 231 | 16 | 443 | 92 | 5 | 1.31 | 1.03–1.67 | 1.31 | 1.00–1.72 | 1.67 | 0.61–4.60 | 0.926 |
| Overall OR | – | – | – | – | – | – | 1.37 | 1.31–1.43 | 1.37 | 1.30–1.44 | 1.82 | 1.55–2.14 | – |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, C.-C.; Wang, M.; Cao, F.-D.; Huang, W.-H.; Xiao, D.; Ye, X.-G.; Ou, M.-L.; Zhang, N.; Zhang, B.-H.; Liu, Y.; et al. Meta-Analysis on Associations of RGS1 and IL12A Polymorphisms with Celiac Disease Risk. Int. J. Mol. Sci. 2016, 17, 457. https://doi.org/10.3390/ijms17040457
Guo C-C, Wang M, Cao F-D, Huang W-H, Xiao D, Ye X-G, Ou M-L, Zhang N, Zhang B-H, Liu Y, et al. Meta-Analysis on Associations of RGS1 and IL12A Polymorphisms with Celiac Disease Risk. International Journal of Molecular Sciences. 2016; 17(4):457. https://doi.org/10.3390/ijms17040457
Chicago/Turabian StyleGuo, Cong-Cong, Man Wang, Feng-Di Cao, Wei-Huang Huang, Di Xiao, Xing-Guang Ye, Mei-Ling Ou, Na Zhang, Bao-Huan Zhang, Yang Liu, and et al. 2016. "Meta-Analysis on Associations of RGS1 and IL12A Polymorphisms with Celiac Disease Risk" International Journal of Molecular Sciences 17, no. 4: 457. https://doi.org/10.3390/ijms17040457
APA StyleGuo, C. -C., Wang, M., Cao, F. -D., Huang, W. -H., Xiao, D., Ye, X. -G., Ou, M. -L., Zhang, N., Zhang, B. -H., Liu, Y., Yang, G., & Jing, C. -X. (2016). Meta-Analysis on Associations of RGS1 and IL12A Polymorphisms with Celiac Disease Risk. International Journal of Molecular Sciences, 17(4), 457. https://doi.org/10.3390/ijms17040457