
Pathways Regulating Spheroid Formation of Human Follicular Thyroid Cancer Cells under Simulated Microgravity Conditions: A Genetic Approach

 ,
, 
Abstract
:
1. Introduction
2. Results and Discussion
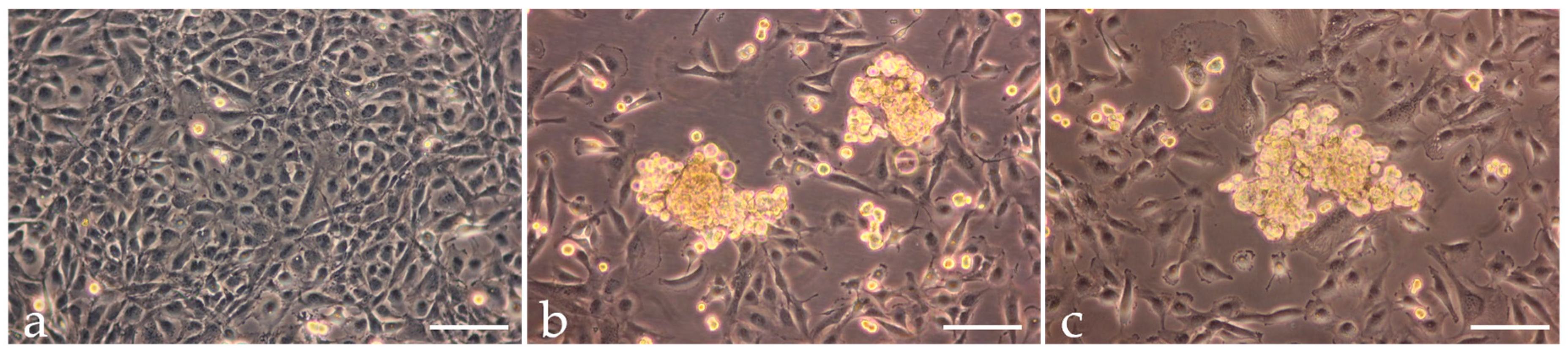
2.1. Spheroid Formation
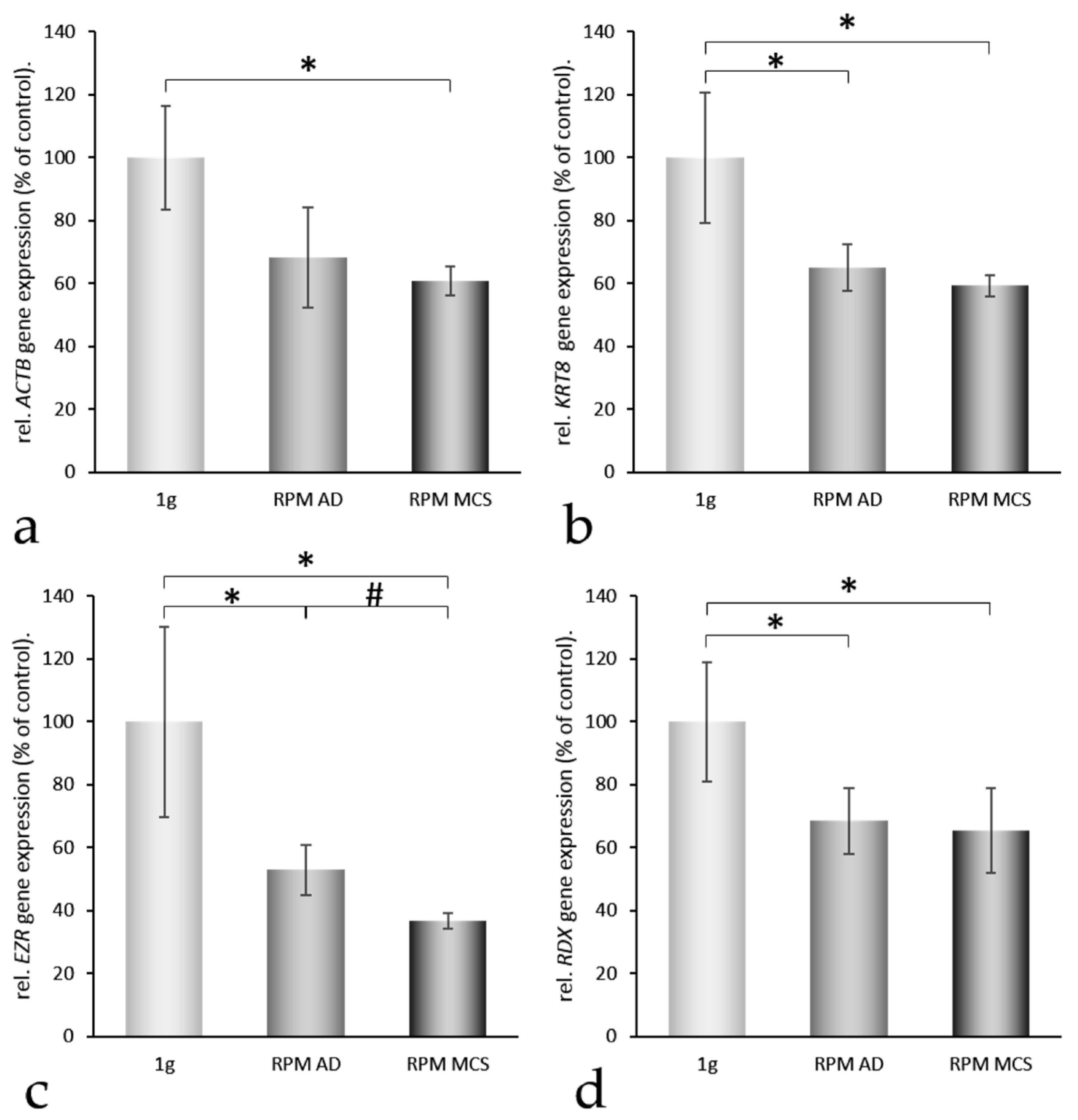
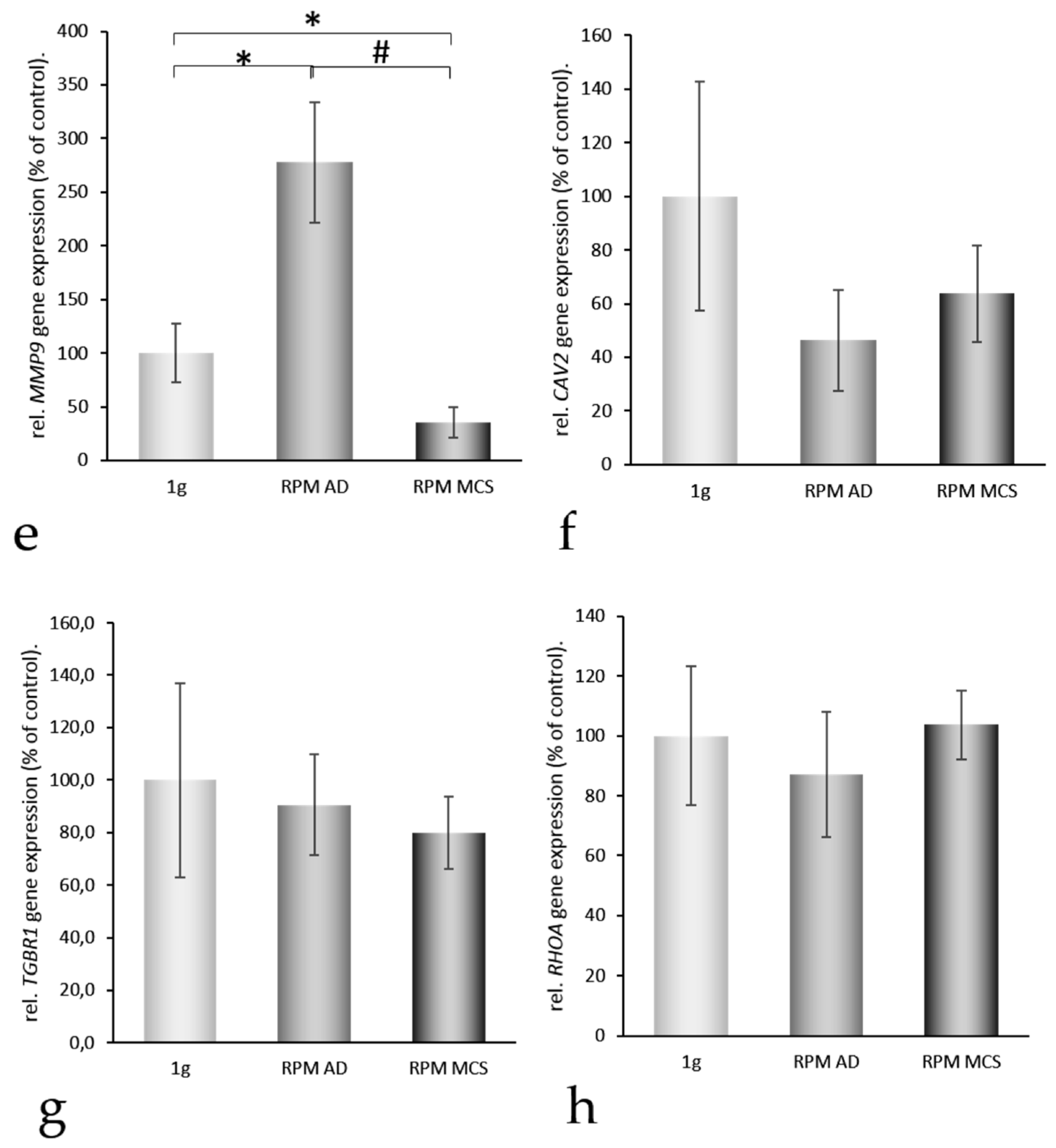
2.2. Impact of Simulated Microgravity on the Activation of Genes Coding for Selected Proteins
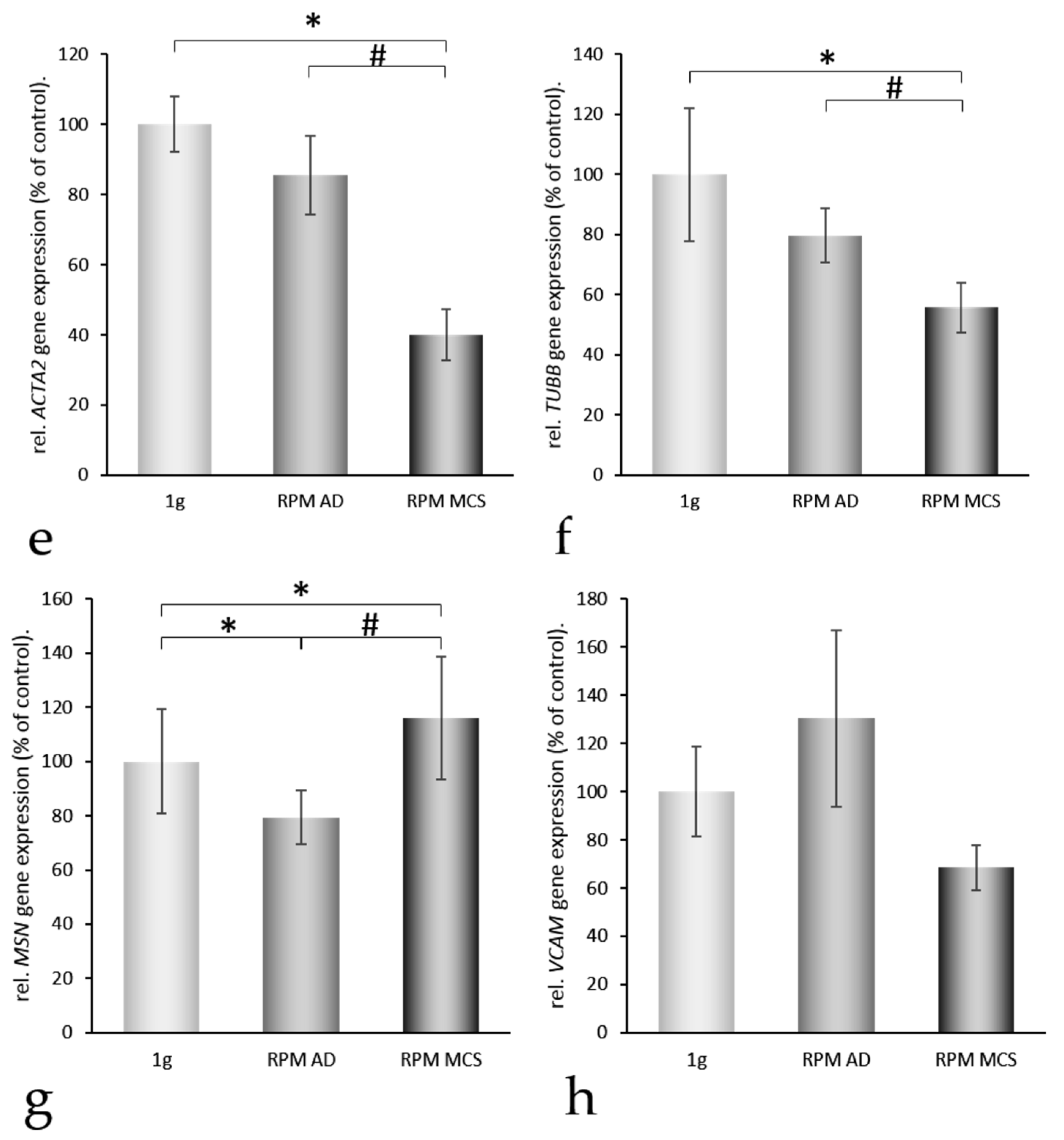
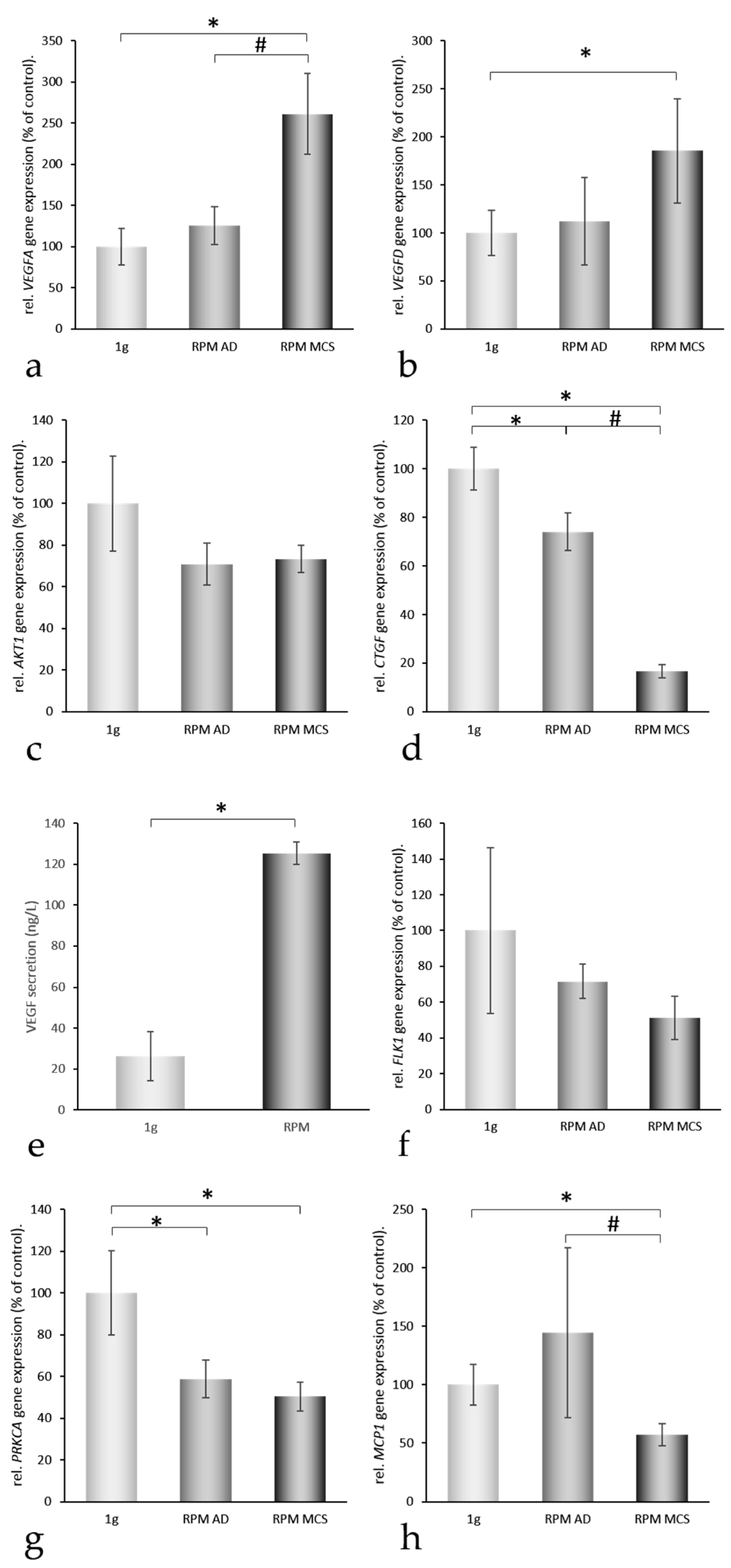
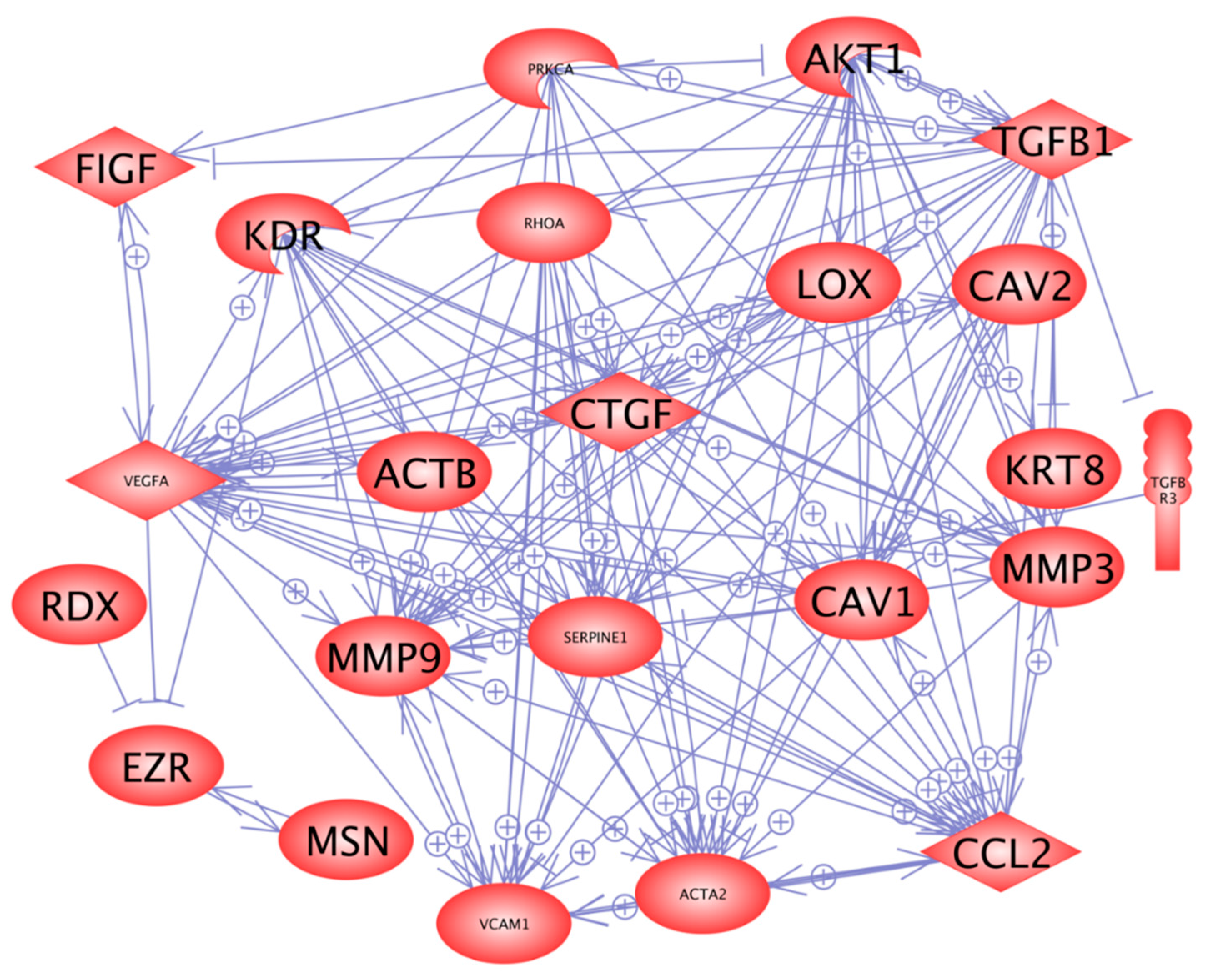
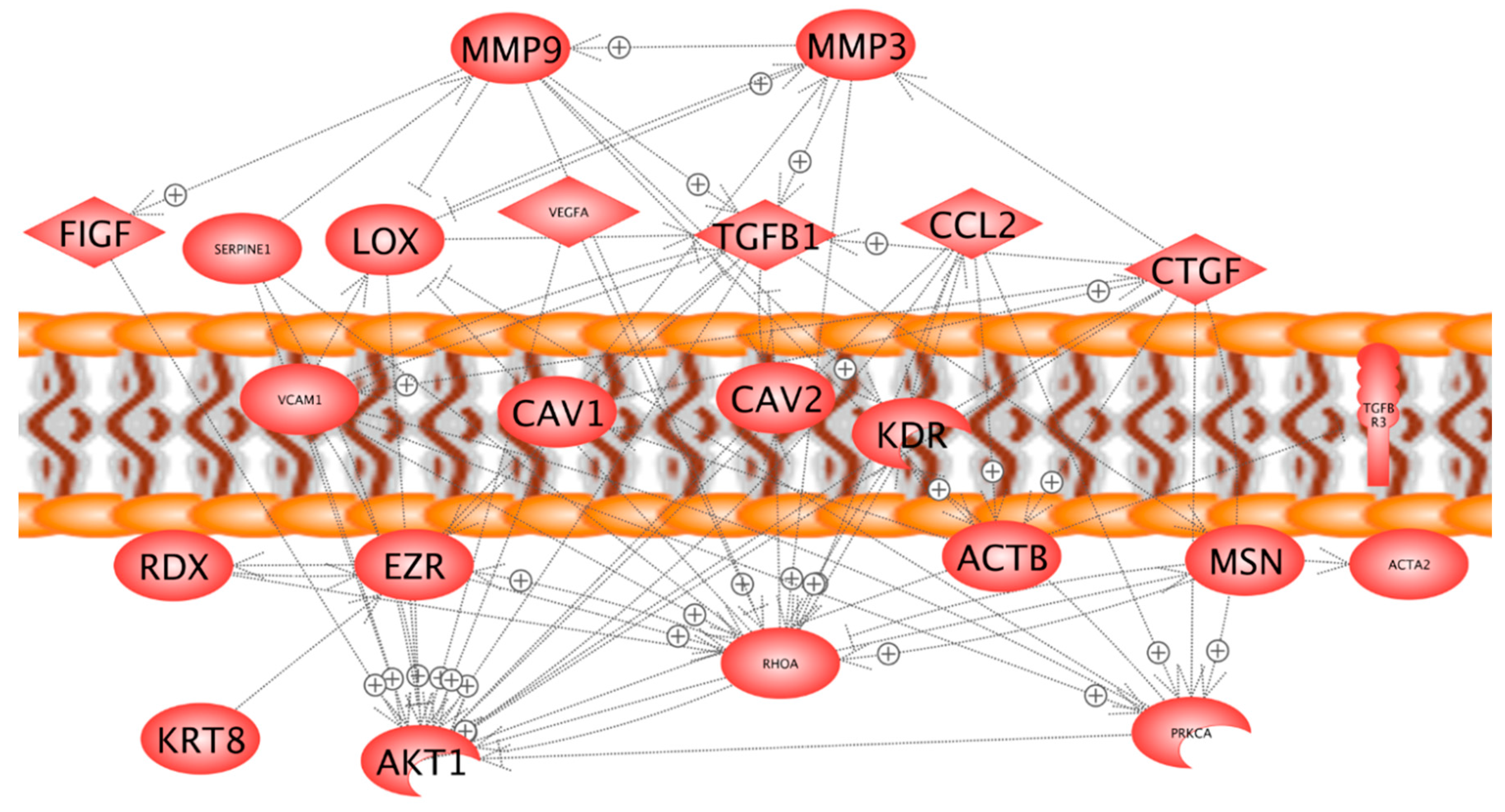
2.3. Three-Dimensional Growth Signaling Pathways
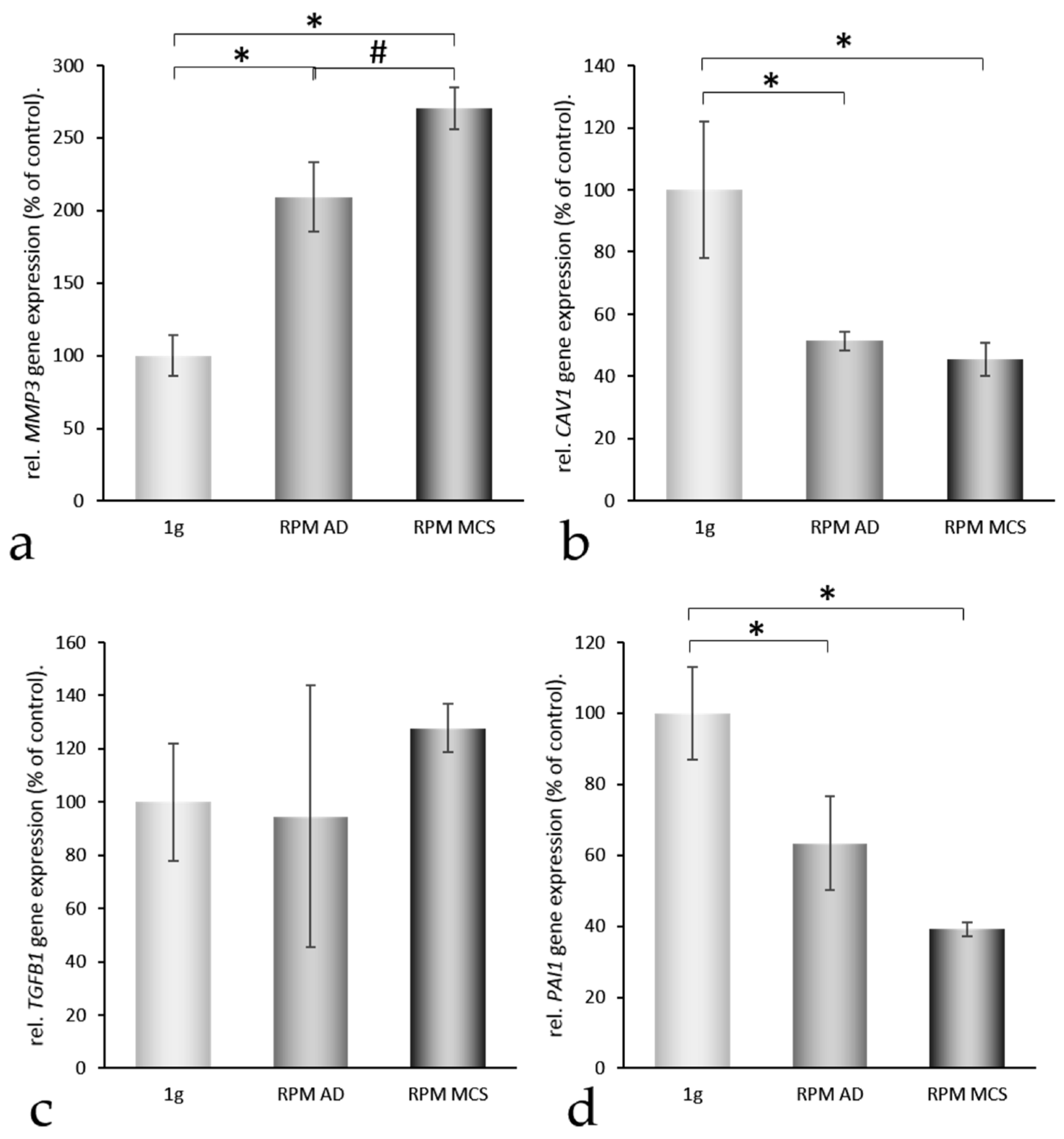
2.4. Factors Regulating the Amount of Extracellular Proteins
2.5. In Silico Search for Mutual Gene Regulation
3. Experimental Section
3.1. Cells
3.2. Random Positioning Machine
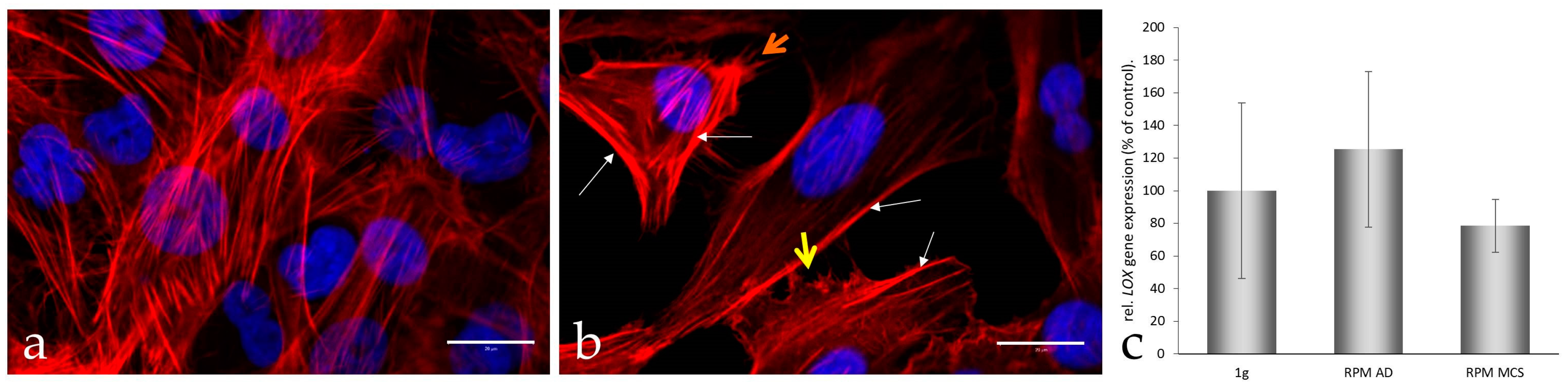
3.3. F-Actin Staining
3.4. RNA Isolation
3.5. Quantitative Real Time PCR
3.6. Pathway Studio Analysis
3.7. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviation
| ACTA2 | Actin aortic smooth muscle (P62736) |
| ACTB | Actin cytoplasmic 1 (P60709) |
| AD | Adherent |
| AKT1 | RAC-α serine/threonine-protein kinase (P31749) |
| CAV1 | Caveolin-1 (Q03135) |
| CAV2 | Caveolin-2 (P51636) |
| CCL2 | C–C motif chemokine 2 (P13500) |
| CTGF | Connective tissue growth factor (P29279) |
| EZR | Ezrin (P15311) |
| FIGF or VEGFD | Vascular endothelial growth factor D (O43915) |
| FLK1 or KDR | Vascular endothelial growth factor receptor 2 (P35968) |
| KRT8 | Keratin type II cytoskeletal 8 (P05787) |
| LOX | Protein-lysine 6-oxidase (P28300) |
| MCS | Multicellular spheroids |
| MCTS | Multicellular tumor spheroid |
| MMP-3 | Matrix metalloproteinase-3 or Stromelysin-1 (P08254) |
| MMP-9 | Matrix metalloproteinase-9 (P14780) |
| MSN | Moesin (P26038) |
| PRKCA | Protein kinase C alpha type (P17252) |
| qPCR | Quantitative real-time PCR |
| RDX | Radixin (P35241) |
| RHOA | protein: Ras homolog gene family, member A; gene: Transforming protein RhoA (P61586) |
| RPM | Random Positioning Machine |
| RWV | Rotating Wall Vessel |
| SERPINE1 | Plasminigen activator inhibitor 1 (P05121) |
| TGFB1 | Transforming growth factor β-1 (P01137) |
| TGFBR1 | Transforming growth factor beta receptor type 1 (P36897) |
| TGFBR3 | Transforming growth factor beta receptor type 3 (Q03167) |
| TUBB | Tubulin β (P07437) |
| VCAM1 | Vascular cellular adhesion protein 1 (P19320) |
| VEGFA | Vascular endothelial growth factor A (P15692) |
References
- Pietsch, J.; Kussian, R.; Sickmann, A.; Bauer, J.; Weber, G.; Nissum, M.; Westphal, K.; Egli, M.; Grosse, J.; Schonberger, J.; et al. Application of free-flow ief to identify protein candidates changing under microgravity conditions. Proteomics 2010, 10, 904–913. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, J.; Sickmann, A.; Weber, G.; Bauer, J.; Egli, M.; Wildgruber, R.; Infanger, M.; Grimm, D. A proteomic approach to analysing spheroid formation of two human thyroid cell lines cultured on a random positioning machine. Proteomics 2011, 11, 2095–2104. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Bauer, J.; Kossmehl, P.; Shakibaei, M.; Schoberger, J.; Pickenhahn, H.; Schulze-Tanzil, G.; Vetter, R.; Eilles, C.; Paul, M.; et al. Simulated microgravity alters differentiation and increases apoptosis in human follicular thyroid carcinoma cells. FASEB J. 2002, 16, 604–606. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Pietsch, J.; Wehland, M.; Schulz, H.; Saar, K.; Hubner, N.; Bauer, J.; Braun, M.; Schwarzwalder, A.; Segerer, J.; et al. Differential gene expression profile and altered cytokine secretion of thyroid cancer cells in space. FASEB J. 2014, 28, 813–835. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Infanger, M.; Westphal, K.; Ulbrich, C.; Pietsch, J.; Kossmehl, P.; Vadrucci, S.; Baatout, S.; Flick, B.; Paul, M.; et al. A delayed type of three-dimensional growth of human endothelial cells under simulated weightlessness. Tissue Eng. A 2009, 15, 2267–2275. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Wehland, M.; Pietsch, J.; Aleshcheva, G.; Wise, P.; van Loon, J.; Ulbrich, C.; Magnusson, N.E.; Infanger, M.; Bauer, J. Growing tissues in real and simulated microgravity: New methods for tissue engineering. Tissue Eng. B 2014, 20, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wehland, M.; Schulz, H.; Saar, K.; Hubner, N.; Infanger, M.; Bauer, J.; Grimm, D. Genomic approach to identify factors that drive the formation of three-dimensional structures by EA.hy926 endothelial cells. PLoS ONE 2013, 8, e64402. [Google Scholar] [CrossRef] [PubMed]
- Kopp, S.; Warnke, E.; Wehland, M.; Aleshcheva, G.; Magnusson, N.E.; Hemmersbach, R.; Corydon, T.J.; Bauer, J.; Infanger, M.; Grimm, D. Mechanisms of three-dimensional growth of thyroid cells during long-term simulated microgravity. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wehland, M.; Aleshcheva, G.; Hauslage, J.; Waßer, K.; Hemmersbach, R.; Infanger, M.; Bauer, J.; Grimm, D. Interleukin-6 expression under gravitational stress due to vibration and hypergravity in follicular thyroid cancer cells. PLoS ONE 2013, 8, e68140. [Google Scholar] [CrossRef] [PubMed]
- Grosse, J.; Wehland, M.; Pietsch, J.; Schulz, H.; Saar, K.; Hubner, N.; Eilles, C.; Bauer, J.; Abou-El-Ardat, K.; Baatout, S.; et al. Gravity-sensitive signaling drives 3-dimensional formation of multicellular thyroid cancer spheroids. FASEB J. 2012, 26, 5124–5140. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, J.; Ma, X.; Wehland, M.; Aleshcheva, G.; Schwarzwälder, A.; Segerer, J.; Birlem, M.; Horn, A.; Bauer, J.; Infanger, M.; et al. Spheroid formation of human thyroid cancer cells in an automated culturing system during the shenzhou-8 space mission. Biomaterials 2013, 34, 7694–7705. [Google Scholar] [PubMed]
- Svejgaard, B.; Wehland, M.; Ma, X.; Kopp, S.; Sahana, J.; Warnke, E.; Aleshcheva, G.; Hemmersbach, R.; Hauslage, J.; Grosse, J.; et al. Common effects on cancer cells exerted by a random positioning machine and a 2d clinostat. PLoS ONE 2015, 10, e0135157. [Google Scholar] [CrossRef] [PubMed]
- Ulbrich, C.; Pietsch, J.; Grosse, J.; Wehland, M.; Schulz, H.; Saar, K.; Hübner, N.; Hauslage, J.; Hemmersbach, R.; Braun, M.; et al. Differential gene regulation under altered gravity conditions in follicular thyroid cancer cells: Relationship between the extracellular matrix and the cytoskeleton. Cell. Physiol. Biochem. 2011, 28, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Warnke, E.; Kopp, S.; Wehland, M.; Hemmersbach, R.; Bauer, J.; Pietsch, J.; Infanger, M.; Grimm, D. Thyroid cells exposed to simutalted microgravity conditions—Comparison of the fast roating clinostat and the random position maschine. Microgravity Sci. Technol. 2015. [Google Scholar] [CrossRef]
- Faute, M.A.D.; Laurent, L.; Ploton, D.; Poupon, M.-F.; Jardillier, J.-C.; Bobichon, H. Distinctive alterations of invasiveness, drug resistance and cell-cell organization in 3D-cultures of MCF-7, a human breast cancer cell line, and its multidrug resistant variant. Clin. Exp. Metastasis 2002, 19, 161–168. [Google Scholar] [CrossRef]
- Kogai, T.; Curcio, F.; Hyman, S.; Cornford, E.M.; Brent, G.A.; Hershman, J.M. Induction of follicle formation in long-term cultured normal human thyroid cells treated with thyrotropin stimulates iodide uptake but not sodium/iodide symporter messenger rna and protein expression. J. Endocrinol. 2000, 167, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Albi, E.; Ambesi-Impiombato, F.S.; Peverini, M.; Damaskopoulou, E.; Fontanini, E.; Lazzarini, R.; Curcio, F.; Perrella, G. Thyrotropin receptor and membrane interactions in FRTL-5 thyroid cell strain in microgravity. Astrobiology 2011, 11, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Pietsch, J.; Wehland, M.; Richter, P.; Strauch, S.M.; Lebert, M.; Magnusson, N.E.; Wise, P.; Bauer, J. The impact of microgravity-based proteomics research. Expert Rev. Proteom. 2014, 11, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Wise, P.; Lebert, M.; Richter, P.; Baatout, S. How and why does the proteome respond to microgravity? Expert Rev. Proteom. 2011, 8, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, J.; Bauer, J.; Egli, M.; Infanger, M.; Wise, P.; Ulbrich, C.; Grimm, D. The effects of weightlessness on the human organism and mammalian cells. Curr. Mol. Med. 2011, 11, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Riwaldt, S.; Bauer, J.; Pietsch, J.; Braun, M.; Segerer, J.; Schwarzwälder, A.; Corydon, T.; Infanger, M.; Grimm, D. The importance of caveolin-1 as key-regulator of three-dimensional growth in thyroid cancer cells cultured under real and simulated microgravity conditions. Int. J. Mol. Sci. 2015, 16, 28296–28310. [Google Scholar] [CrossRef] [PubMed]
- Riwaldt, S.; Pietsch, J.; Sickmann, A.; Bauer, J.; Braun, M.; Segerer, J.; Schwarzwälder, A.; Aleshcheva, G.; Corydon, T.J.; Infanger, M.; et al. Identification of proteins involved in inhibition of spheroid formation under microgravity. Proteomics 2015, 15, 2945–2952. [Google Scholar] [CrossRef] [PubMed]
- Kanbe, N.; Tanaka, A.; Kanbe, M.; Itakura, A.; Kurosawa, M.; Matsuda, H. Human mast cells produce matrix metalloproteinase 9. Eur J. Immunol. 1999, 29, 2645–2649. [Google Scholar] [CrossRef]
- Kim, E.M.; Hwang, O. Role of matrix metalloproteinase-3 in neurodegeneration. J. Neurochem. 2011, 116, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Plow, E.F.; Herren, T.; Redlitz, A.; Miles, L.A.; Hoover-Plow, J.L. The cell biology of the plasminogen system. FASEB J. 1995, 9, 939–945. [Google Scholar] [PubMed]
- Bolat, F.; Gumurdulu, D.; Erkanli, S.; Kayaselcuk, F.; Zeren, H.; Ali Vardar, M.; Kuscu, E. Maspin overexpression correlates with increased expression of vascular endothelial growth factors A, C, and D in human ovarian carcinoma. Pathol.-Res. Pract. 2008, 204, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Butler, S.M.; Abrassart, J.M.; Hubbell, M.C.; Adeoye, O.; Semotiuk, A.; Williams, J.M.; Mata-Greenwood, E.; Khorram, O.; Pearce, W.J. Contributions of VEGF to age-dependent transmural gradients in contractile protein expression in ovine carotid arteries. Am. J. Physiol. Cell Physiol. 2011, 301, C653–C666. [Google Scholar] [CrossRef] [PubMed]
- Wisniewska-Kruk, J.; Hoeben, K.A.; Vogels, I.M.C.; Gaillard, P.J.; Van Noorden, C.J.F.; Schlingemann, R.O.; Klaassen, I. A novel co-culture model of the blood-retinal barrier based on primary retinal endothelial cells, pericytes and astrocytes. Exp. Eye Res. 2012, 96, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, L.; Li, Z.; Gong, G.; Chen, P.; Liu, H.; Wang, J.; Liu, Y.; Wu, X. The role of miR-205 in the VEGF-mediated promotion of human ovarian cancer cell invasion. Gynecol. Oncol. 2015, 137, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.; Stathopoulou, M.G.; Dade, S.; Ndiaye, N.C.; Azimi-Nezhad, M.; Murray, H.; Masson, C.; Lamont, J.; Fitzgerald, P.; Visvikis-Siest, S. Angiogenesis related genes NOS3, CD14, MMP3 and IL4R are associated to VEGF gene expression and circulating levels in healthy adults. BMC Med. Genet. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, S.; Nakamura, K.; Iwai, S.; Murakami, M.; Itoh, T.; Kijima, H.; Shipley, J.M.; Senior, R.M.; Shibuya, M. MMP9 induction by vascular endothelial growth factor receptor-1 is involved in lung-specific metastasis. Cancer Cell 2002, 2, 289–300. [Google Scholar] [CrossRef]
- Tang, C.-H.; Tsai, C.-C. CCL2 increases MMP-9 expression and cell motility in human chondrosarcoma cells via the Ras/Raf/MEK/ERK/NF-κB signaling pathway. Biochem. Pharmacol. 2012, 83, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Nishimura, Y.; Shigematsu, S.; Takeuchi, Y.; Nakamura, J.; Aizawa, T.; Hashizume, K. Vascular endothelial cell growth factor attenuates actions of transforming growth factor-β in human endothelial cells. J. Biol. Chem. 2004, 279, 55104–55108. [Google Scholar] [CrossRef] [PubMed]
- Liekens, S.; de Clercq, E.; Neyts, J. Angiogenesis: Regulators and clinical applications. Biochem. Pharmacol. 2001, 61, 253–270. [Google Scholar] [CrossRef]
- Hata, Y.; Rook, S.L.; Aiello, L.P. Basic fibroblast growth factor induces expression of VEGF receptor KDR through a protein kinase C and p44/p42 mitogen-activated protein kinase-dependent pathway. Diabetes 1999, 48, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, P.; di Benedetto, P.; Capece, D.; Zazzeroni, F.; Liakouli, V.; Ruscitti, P.; Pantano, I.; Berardicurti, O.; Carubbi, F.; Alesse, E.; et al. Impaired Cav-1 expression in SSc mesenchymal cells upregulates VEGF signaling: A link between vascular involvement and fibrosis. Fibrogen. Tissue Repair 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Fearnley, G.W.; Odell, A.F.; Latham, A.M.; Mughal, N.A.; Bruns, A.F.; Burgoyne, N.J.; Homer-Vanniasinkam, S.; Zachary, I.C.; Hollstein, M.C.; Wheatcroft, S.B.; et al. VEGF-A isoforms differentially regulate ATF-2-dependent VCAM-1 gene expression and endothelial-leukocyte interactions. Mol. Biol. Cell 2014, 25, 2509–2521. [Google Scholar] [CrossRef] [PubMed]
- Coral, K.; Madhavan, J.; Pukhraj, R.; Angayarkanni, N. High glucose induced differential expression of lysyl oxidase and its isoform in ARPE-19 cells. Curr. Eye Res. 2013, 38, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.; Wehland, M.; Pietsch, J.; Sickmann, A.; Weber, G.; Grimm, D. Annotated gene and proteome data support recognition of interconnections between the results of different experiments in space research. Microgravity Sci. Technol. 2015. [Google Scholar] [CrossRef]
- Blom, I.E.; Goldschmeding, R.; Leask, A. Gene regulation of connective tissue growth factor: New targets for antifibrotic therapy? Matrix Biol. 2002, 21, 473–482. [Google Scholar] [CrossRef]
- Marumo, T.; Schini-Kerth, V.B.; Busse, R. Vascular endothelial growth factor activates nuclear factor-κB and induces monocyte chemoattractant protein-1 in bovine retinal endothelial cells. Diabetes 1999, 48, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Masiello, M.G.; Cucina, A.; Proietti, S.; Palombo, A.; Coluccia, P.; D’Anselmi, F.; Dinicola, S.; Pasqualato, A.; Morini, V.; Bizzarri, M. Phenotypic switch induced by simulated microgravity on MDA-MB-231 breast cancer cells. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Testa, F.; Palombo, A.; Dinicola, S.; D’Anselmi, F.; Proietti, S.; Pasqualato, A.; Masiello, M.G.; Coluccia, P.; Cucina, A.; Bizzarri, M. Fractal analysis of shape changes in murine osteoblasts cultured under simulated microgravity. Rend. Fis. Acc. Lincei 2014, 25, 39–47. [Google Scholar] [CrossRef]
- Infanger, M.; Kossmehl, P.; Shakibaei, M.; Baatout, S.; Witzing, A.; Grosse, J.; Bauer, J.; Cogoli, A.; Faramarzi, S.; Derradji, H.; et al. Induction of three-dimensional assembly and increase in apoptosis of human endothelial cells by simulated microgravity: Impact of vascular endothelial growth factor. Apoptosis 2006, 11, 749–764. [Google Scholar] [CrossRef] [PubMed]
- Kondepudi, D.K.; Storm, P.B. Gravity detection through bifurcation. Adv. Space Res. 1992, 12, 7–14. [Google Scholar] [CrossRef]
- Mesland, D.A. Possible actions of gravity on the cellular machinery. Adv. Space Res. 1992, 12, 15–25. [Google Scholar] [CrossRef]
- Tabony, J. Morphological bifurcations involving reaction-diffusion processes during microtubule formation. Science 1994, 264, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Junker, J.P.; van Oudenaarden, A. Every cell is special: Genome-wide studies add a new dimension to single-cell biology. Cell 2014, 157, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Schonberger, J.; Bauer, J.; Spruss, T.; Weber, G.; Chahoud, I.; Eilles, C.; Grimm, D. Establishment and characterization of the follicular thyroid carcinoma cell line ML-1. J. Mol. Med. 2000, 78, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.; Grimm, D.; Bauer, J. Application of binary buffer systems to free flow cell electrophoresis. Electrophoresis 2000, 21, 325–328. [Google Scholar] [CrossRef]
- Pietsch, J.; Sickmann, A.; Weber, G.; Bauer, J.; Egli, M.; Wildgruber, R.; Infanger, M.; Grimm, D. Metabolic enzyme diversity in different human thyroid cell lines and their sensitivity to gravitational forces. Proteomics 2012, 12, 2539–2546. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, S.; Yonemura, S. Cortical actin organization: Lessons from ERM (ezrin/radixin/moesin) proteins. J. Biol. Chem. 1999, 274, 34507–34510. [Google Scholar] [CrossRef] [PubMed]
- Corydon, T.J.; Kopp, S.; Wehland, M.; Braun, M.; Schutte, A.; Mayer, T.; Hulsing, T.; Oltmann, H.; Schmitz, B.; Hemmersbach, R.; et al. Alterations of the cytoskeleton in human cells in space proved by life-cell imaging. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Haynes, J.; Srivastava, J.; Madson, N.; Wittmann, T.; Barber, D.L. Dynamic actin remodeling during epithelial-mesenchymal transition depends on increased moesin expression. Mol. Biol. Cell 2011, 22, 4750–4764. [Google Scholar] [CrossRef] [PubMed]
- Neisch, A.L.; Fehon, R.G. Ezrin, radixin and moesin: Key regulators of membrane–cortex interactions and signaling. Curr. Opin. Cell Biol. 2011, 23, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-S.; Hinds, P.W. Increased ezrin expression and activation by CDK5 coincident with acquisition of the senescent phenotype. Mol. Cell 2003, 11, 1163–1176. [Google Scholar] [CrossRef]
- Inoki, I. Connective tissue growth factor binds vascular endothelial growth factor (VEGF) and inhibits VEGF-induced angiogenesis. FASEB J. 2001, 16, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Bao, P.; Kodra, A.; Tomic-Canic, M.; Golinko, M.S.; Ehrlich, H.P.; Brem, H. The role of vascular endothelial growth factor in wound healing. J. Surg. Res. 2009, 153, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Bauer, J.; Schoenberger, J. Blockade of neoangiogenesis, a new and promising technique to control the growth of malignant tumors and their metastases. Curr. Vasc. Pharmacol. 2009, 7, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Bass, M.B.; Sherman, S.I.; Schlumberger, M.J.; Davis, M.T.; Kivman, L.; Khoo, H.-M.; Notari, K.H.; Peach, M.; Hei, Y.-J.; Patterson, S.D. Biomarkers as predictors of response to treatment with motesanib in patients with progressive advanced thyroid cancer. J. Clin. Endocrinol. Metab. 2010, 95, 5018–5027. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Bauer, J.; Ulbrich, C.; Westphal, K.; Wehland, M.; Infanger, M.; Aleshcheva, G.; Pietsch, J.; Ghardi, M.; Beck, M.; et al. Different responsiveness of endothelial cells to vascular endothelial growth factor and basic fibroblast growth factor added to culture media under gravity and simulated microgravity. Tissue Eng. A 2010, 16, 1559–1573. [Google Scholar] [CrossRef] [PubMed]
- George, M.L.; Tutton, M.G.; Janssen, F.; Arnaout, A.; Abulafi, A.M.; Eccles, S.A.; Swift, R.I. VEGF-A, VEGF-C, and VEGF-D in colorectal cancer progression. Neoplasia 2001, 3, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Nersita, R.; Matrone, A.; Klain, M.; Scavuzzo, F.; Vitolo, G.; Abbondanza, C.; Carlino, M.V.; Giacco, V.; Amato, G.; Carella, C. Decreased serum vascular endothelial growth factor-D levels in metastatic patients with differentiated thyroid carcinoma. Clin. Endocrinol. 2011, 76, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Edgell, C.J.; McDonald, C.C.; Graham, J.B. Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proc. Natl. Acad. Sci. USA 1983, 80, 3734–3737. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.J.; Ren, Y.Q.; Wang, G.P.; Song, Q.; Li, M.; Jiang, S.S.; Ning, T.; Guan, Y.S.; Yang, J.L.; Luo, F. Biological behaviors and proteomics analysis of hybrid cell line Eahy926 and its parent cell line A549. J. Exp. Clin. Cancer Res. 2009, 28. [Google Scholar] [CrossRef] [PubMed]
- Mannello, F.; Medda, V. Nuclear localization of matrix metalloproteinases. Prog. Histochem. Cytochem. 2012, 47, 27–58. [Google Scholar] [CrossRef] [PubMed]
- Werle, M.J.; VanSaun, M. Activity dependent removal of agrin from synaptic basal lamina by matrix metalloproteinase 3. J. Neurocytol. 2003, 32, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Warnke, E.; Pietsch, J.; Wehland, M.; Bauer, J.; Infanger, M.; Gorog, M.; Hemmersbach, R.; Braun, M.; Ma, X.; Sahana, J.; et al. Spheroid formation of human thyroid cancer cells under simulated microgravity: A possible role of ctgf and cav1. Cell Commun. Signal. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.-Y.; Chang, C.-C.; Prakash, E.; Kuo, M.-L. Connective tissue growth factor (CTGF) and cancer progression. J. Biomed. Sci. 2008, 15, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Zhang, Q.; Mao, Z.; Chen, J.; Wang, X.; Qu, J.; Zhang, J.; Jin, D. CTGF is overexpressed in papillary thyroid carcinoma and promotes the growth of papillary thyroid cancer cells. Tumor Biol. 2011, 32, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Molè, D.; Gentilin, E.; Gagliano, T.; Tagliati, F.; Bondanelli, M.; Pelizzo, M.R.; Rossi, M.; Filieri, C.; Pansini, G.; degli Uberti, E.C.; et al. Protein kinase C: A putative new target for the control of human medullary thyroid carcinoma cell proliferation in vitro. Endocrinology 2012, 153, 2088–2098. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Way, K.J.; Arikawa, E.; Chou, E.; Opland, D.M.; Clermont, A.; Isshiki, K.; Ma, R.C.W.; Scott, J.A.; Schoen, F.J.; et al. Differential regulation of angiotensin II-induced expression of connective tissue growth factor by protein kinase C isoforms in the myocardium. J. Biol. Chem. 2005, 280, 15719–15726. [Google Scholar] [CrossRef] [PubMed]
- Tourkina, E.; Gooz, P.; Pannu, J.; Bonner, M.; Scholz, D.; Hacker, S.; Silver, R.M.; Trojanowska, M.; Hoffman, S. Opposing effects of protein kinase C and protein kinase C on collagen expression by human lung fibroblasts are mediated via MEK/ERK and caveolin-1 signaling. J. Biol. Chem. 2005, 280, 13879–13887. [Google Scholar] [CrossRef] [PubMed]
- Estour, B.; Herle, A.J.; Juillard, G.J.F.; Totanes, T.L.; Sparkes, R.S.; Giuliano, A.E.; Klandorf, H. Characterization of a human follicular thyroid carcinoma cell line (UCLA RO 82 W-1). Virchows Arch. B Cell Pathol. Mol. Pathol. 1989, 57, 167–174. [Google Scholar] [CrossRef]
- Aleshcheva, G.; Sahana, J.; Ma, X.; Hauslage, J.; Hemmersbach, R.; Egli, M.; Infanger, M.; Bauer, J.; Grimm, D. Changes in morphology, gene expression and protein content in chondrocytes cultured on a random positioning machine. PLoS ONE 2013, 8, e79057. [Google Scholar] [CrossRef] [PubMed]
- Aleshcheva, G.; Wehland, M.; Sahana, J.; Bauer, J.; Corydon, T.J.; Hemmersbach, R.; Frett, T.; Egli, M.; Infanger, M.; Grosse, J.; et al. Moderate alterations of the cytoskeleton in human chondrocytes after short-term microgravity produced by parabolic flight maneuvers could be prevented by upregulation of BMP-2 and SOX-9. FASEB J. 2015, 29, 2303–2314. [Google Scholar] [CrossRef] [PubMed]
- Wehland, M.; Aleshcheva, G.; Schulz, H.; Saar, K.; Hubner, N.; Hemmersbach, R.; Braun, M.; Ma, X.; Frett, T.; Warnke, E.; et al. Differential gene expression of human chondrocytes cultured under short-term altered gravity conditions during parabolic flight maneuvers. Cell Commun. Signal. 2015, 13. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Bonchev, D. A survey of current software for network analysis in molecular biology. Hum. Genom. 2010, 4, 353–360. [Google Scholar] [CrossRef]
- Meli, A.; Perrella, G.; Curcio, F.; Hemmersbach, R.; Neubert, J.; Impiombato, F.A. Response to thyrotropin of normal thyroid follicular cell strain FRTL5 in hypergravity. Biochimie 1999, 81, 281–285. [Google Scholar] [CrossRef]
- Meli, A.; Perrella, G.; Curcio, F.; Impiombato, F.A. Response to hypergravity of normal in vitro cultured follicular cells from thyroid. Acta Astronaut. 1998, 42, 465–472. [Google Scholar] [CrossRef]
- Albi, E.; Curcio, F.; Spelat, R.; Lazzarini, A.; Lazzarini, R.; Cataldi, S.; Loreti, E.; Ferri, I.; Ambesi-Impiombato, F.S. Loss of parafollicular cells during gravitational changes (microgravity, hypergravity) and the secret effect of pleiotrophin. PLoS ONE 2012, 7, e48518. [Google Scholar] [CrossRef] [PubMed]
- Albi, E.; Curcio, F.; Spelat, R.; Lazzarini, A.; Lazzarini, R.; Loreti, E.; Ferri, I.; Ambesi-Impiombato, F.S. Observing the mouse thyroid sphingomyelin under space conditions: A case study from the mds mission in comparison with hypergravity conditions. Astrobiology 2012, 12, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Zhou, A.; Gordon, R.E.; Henderson, S.C.; Schwartz, A.E.; Friedman, E.W.; Davies, T.F. Thyroid organoid formation in simulated microgravity: Influence of keratinocyte growth factor. Thyroid 2000, 10, 481–487. [Google Scholar] [PubMed]
- Masini, M.A.; Albi, E.; Barmo, C.; Bonfiglio, T.; Bruni, L.; Canesi, L.; Cataldi, S.; Curcio, F.; D’Amora, M.; Ferri, I.; et al. The impact of long-term exposure to space environment on adult mammalian organisms: A study on mouse thyroid and testis. PLoS ONE 2012, 7, e35418. [Google Scholar] [CrossRef] [PubMed]
- Albi, E.; Curcio, F.; Lazzarini, A.; Floridi, A.; Cataldi, S.; Lazzarini, R.; Loreti, E.; Ferri, I.; Ambesi-Impiombato, F.S. How microgravity changes galectin-3 in thyroid follicles. Biomed Res. Int. 2014, 2014, 652863. [Google Scholar] [CrossRef] [PubMed]
- Albi, E.; Curcio, F.; Lazzarini, A.; Floridi, A.; Cataldi, S.; Lazzarini, R.; Loreti, E.; Ferri, I.; Ambesi-Impiombato, F.S. A firmer understanding of the effect of hypergravity on thyroid tissue: Cholesterol and thyrotropin receptor. PLoS ONE 2014, 9, e98250. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.L.; Souza, G.R. Using space-based investigations to inform cancer research on earth. Nat. Rev. Cancer 2013, 13, 315–327. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | F-Primer | Sequence | R-Primer | Sequence |
|---|---|---|---|---|
| 18S rRNA | 18S-F | GGAGCCTGCGGCTTAATTT | 18S-R | CAACTAAGAACGGCCATGCA |
| ACTA2 | ACTA2-F | GAGCGTGGCTATTCCTTCGT | ACTA2-R | TTCAAAGTCCAGAGCTACATAACACAGT |
| ACTB | ACTB-F | TGCCGACAGGATGCAGAAG | ACTB-R | GCCGATCCACACGGAGTACT |
| AKT1 | AKT1-F | CTTCTATGGCGCTGAGATTGTG | AKT1-R | CAGCATGAGGTTCTCCAGCT |
| CAV1 | CAV1-F | CCTCCTCACAGTTTTCATCCA | CAV1-R | TGTAGATGTTGCCCTGTTCC |
| CAV2 | CAV2-F | GATCCCCACCGGCTCAAC | CAV2-R | CACCGGCTCTGCGATCA |
| CTGF | CTGF-F | ACAAGGGCCTCTTCTGTGACTT | CTGF-F | GGTACACCGTACCACCGAAGAT |
| EZR | EZR-F | GCAATCCAGCCAAATACAACTG | EZR-R | CCACATAGTGGAGGCCAAAGTAC |
| FLK1 | FLK1-F | TCTTCTGGCTACTTCTTGTCATCATC | FLK1-R | GATGGACAAGTAGCCTGTCTTCAGT |
| KRT8 | KRT8-F | GATCTCTGAGATGAACCGGAACA | KRT8-R | GCTCGGCATCTGCAATGG |
| LOX | LOX-F | TGGGAATGGCACAGTTGTCA | LOX-R | AGCCACTCTCCTCTGGGTGTT |
| MCP1 | MCP1-F | GCTATAGAAGAATCACCAGCAGCAA | MCP1-R | TGGAATCCTGAACCCACTTCTG |
| MMP3 | MMP3-F | ACAAAGGATACAACAGGGACCAA | MMP3-R | TAGAGTGGGTACATCAAAGCTTCAGT |
| MMP9 | MMP9-F | CCTGGAGACCTGAGAACCAATC | MMP9-R | TTCGACTCTCCACGCATCTCT |
| MSN | MSN-F | GAAATTTGTCATCAAGCCCATTG | MSN-R | CCATGCACAAGGCCAAGAT |
| PAI1 | PAI1-F | AGGCTGACTTCACGAGTCTTTCA | PAI1-R | CACTCTCGTTCACCTCGATCTTC |
| PRKCA | PRKCA-F | TGGGTCACTGCTCTATGGACTTATC | PRKCA-R | CGCCCCCTCTTCTCAGTGT |
| RDX | RDX-F | GAAAATGCCGAAACCAATCAA | RDX-R | GTATTGGGCTGAATGGCAAATT |
| RHOA | RHOA-F | CGTTAGTCCACGGTCTGGTC | RHOA-R | GCCATTGCTCAGGCAACGAA |
| TGFB1 | TGFB1-F | CACCCGCGTGCTAATGGT | TGFB1-R | AGAGCAACACGGGTTCAGGTA |
| TGFBR1 | TGFBR1-F | CGCACTGTCATTCACCATCG | TGFBR1-R | CACGGAACCACGAACGTTC |
| TUBB | TUBB-F | CTGGACCGCATCTCTGTGTACTAC | TUBB-R | GACCTGAGCGAACAGAGTCCAT |
| VCAM | VCAM-F | CATGGAATTCGAACCCAAACA | VCAM-R | GGCTGACCAAGACGGTTGTATC |
| VEGFA | VEGFA-F | GCGCTGATAGACATCCATGAAC | VEGFA-R | CTACCTCCACCATGCCAAGTG |
| VEGFD | VEGFD-F | TGCAGGAGGAAAATCCACTTG | VEGFD-R | CTCGCAACGATCTTCGTCAA |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riwaldt, S.; Bauer, J.; Wehland, M.; Slumstrup, L.; Kopp, S.; Warnke, E.; Dittrich, A.; Magnusson, N.E.; Pietsch, J.; Corydon, T.J.; et al. Pathways Regulating Spheroid Formation of Human Follicular Thyroid Cancer Cells under Simulated Microgravity Conditions: A Genetic Approach. Int. J. Mol. Sci. 2016, 17, 528. https://doi.org/10.3390/ijms17040528
Riwaldt S, Bauer J, Wehland M, Slumstrup L, Kopp S, Warnke E, Dittrich A, Magnusson NE, Pietsch J, Corydon TJ, et al. Pathways Regulating Spheroid Formation of Human Follicular Thyroid Cancer Cells under Simulated Microgravity Conditions: A Genetic Approach. International Journal of Molecular Sciences. 2016; 17(4):528. https://doi.org/10.3390/ijms17040528
Chicago/Turabian StyleRiwaldt, Stefan, Johann Bauer, Markus Wehland, Lasse Slumstrup, Sascha Kopp, Elisabeth Warnke, Anita Dittrich, Nils E. Magnusson, Jessica Pietsch, Thomas J. Corydon, and et al. 2016. "Pathways Regulating Spheroid Formation of Human Follicular Thyroid Cancer Cells under Simulated Microgravity Conditions: A Genetic Approach" International Journal of Molecular Sciences 17, no. 4: 528. https://doi.org/10.3390/ijms17040528
APA StyleRiwaldt, S., Bauer, J., Wehland, M., Slumstrup, L., Kopp, S., Warnke, E., Dittrich, A., Magnusson, N. E., Pietsch, J., Corydon, T. J., Infanger, M., & Grimm, D. (2016). Pathways Regulating Spheroid Formation of Human Follicular Thyroid Cancer Cells under Simulated Microgravity Conditions: A Genetic Approach. International Journal of Molecular Sciences, 17(4), 528. https://doi.org/10.3390/ijms17040528