
Exogenous R-Spondin1 Induces Precocious Telogen-to-Anagen Transition in Mouse Hair Follicles


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
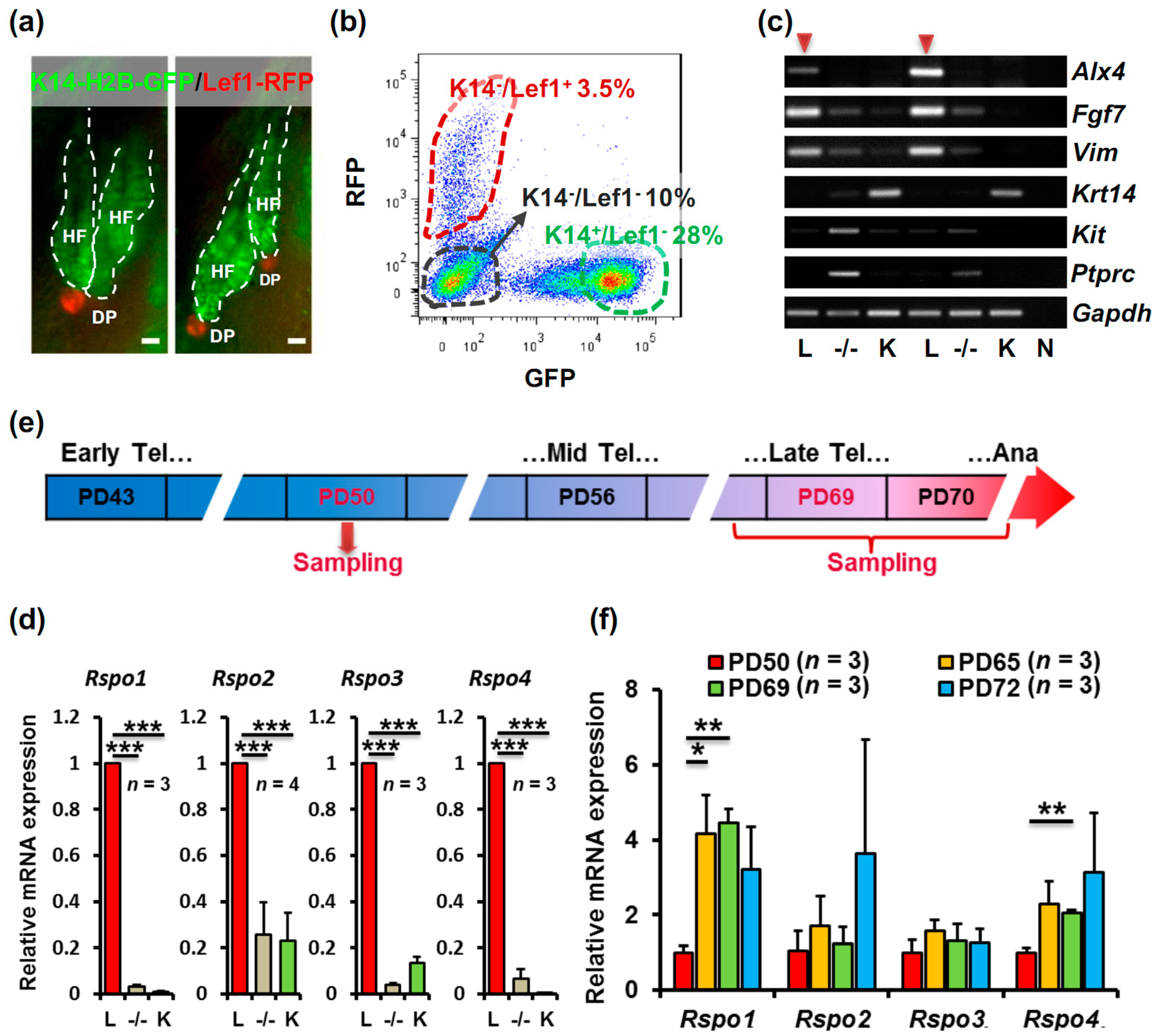
2.1. R-Spondin1-4 Expression Is Enriched in the Telogen Hair Follicle Dermal Papilla, and R-Spondin1 Expression Is Upregulated before Anagen Entry
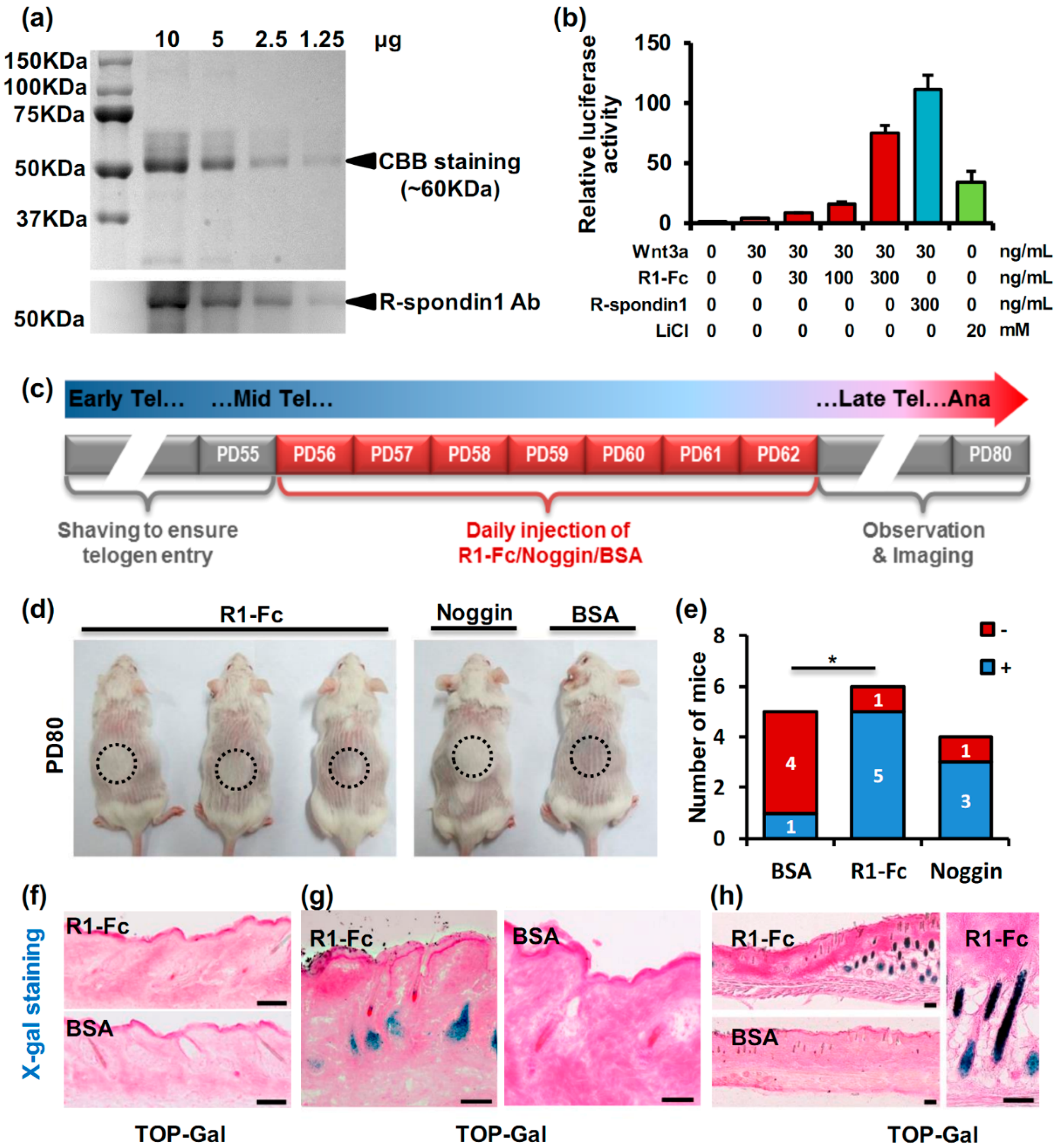
2.2. R-Spondin1 Injection Leads to Precocious Anagen Entry
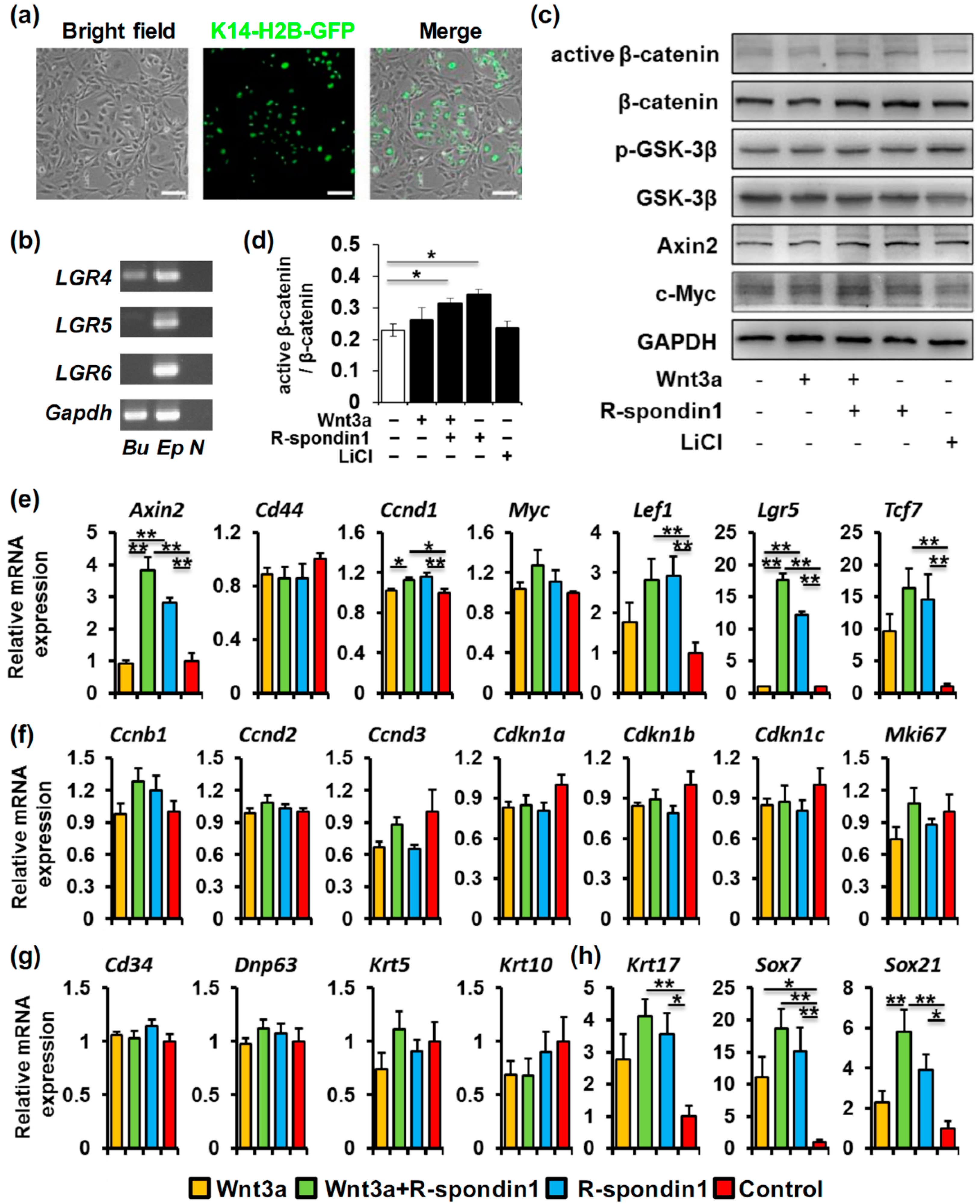
2.3. R-Spondin1 Activates Wnt/β-Catenin Pathway in Bulge Stem Cells and Regulate HFSC Fate Determination in Vitro
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Fluorescence Activated Cell Sorting (FACS)
4.3. Immunostaining
4.4. Alkaline Phosphatase (AP) Staining
4.5. R-Spondin1-Fc Protein Production and Verification
4.6. TCF-Luciferase Assay
4.7. R-Spondin1-Fc Protein Injection
4.8. Skin Harvest and β-Galactosidase (LacZ) Staining
4.9. In Vitro Bulge Stem Cell Culture and Treatment
4.10. MTS Cell Proliferation Assay
4.11. RNA Extraction and PCR
4.12. Protein Extraction and Western Blotting
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lim, X.; Nusse, R. Wnt signaling in skin development, homeostasis, and disease. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, H.; Duan, E. Epidermal development in mammals: Key regulators, signals from beneath, and stem cells. Int. J. Mol. Sci. 2013, 14, 10869–10895. [Google Scholar] [CrossRef] [PubMed]
- Morgan, B.A. The dermal papilla: An instructive niche for epithelial stem and progenitor cells in development and regeneration of the hair follicle. Cold Spring Harb. Perspect. Med. 2014, 4, a015180. [Google Scholar] [CrossRef] [PubMed]
- Carmon, K.S.; Gong, X.; Lin, Q.; Thomas, A.; Liu, Q. R-spondins function as ligands of the orphan receptors LGR4 and LGR5 to regulate Wnt/β-catenin signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 11452–11457. [Google Scholar] [CrossRef] [PubMed]
- De Lau, W.; Barker, N.; Low, T.Y.; Koo, B.K.; Li, V.S.; Teunissen, H.; Kujala, P.; Haegebarth, A.; Peters, P.J.; van de Wetering, M.; et al. LGR5 homologues associate with Wnt receptors and mediate R-spondin signalling. Nature 2011, 476, 293–297. [Google Scholar] [CrossRef] [PubMed]
- De Lau, W.; Peng, W.C.; Gros, P.; Clevers, H. The R-spondin/LGR5/RNF43 module: Regulator of Wnt signal strength. Genes Dev. 2014, 28, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Glinka, A.; Dolde, C.; Kirsch, N.; Huang, Y.L.; Kazanskaya, O.; Ingelfinger, D.; Boutros, M.; Cruciat, C.M.; Niehrs, C. LGR4 and LGR5 are R-spondin receptors mediating Wnt/β-catenin and Wnt/PCP signalling. EMBO Rep. 2011, 12, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Ruffner, H.; Sprunger, J.; Charlat, O.; Leighton-Davies, J.; Grosshans, B.; Salathe, A.; Zietzling, S.; Beck, V.; Therier, M.; Isken, A.; et al. R-spondin potentiates Wnt/β-catenin signaling through orphan receptors LGR4 and LGR5. PLoS ONE 2012, 7, e40976. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.X.; Xie, Y.; Zhang, Y.; Charlat, O.; Oster, E.; Avello, M.; Lei, H.; Mickanin, C.; Liu, D.; Ruffner, H.; et al. ZNRF3 promotes Wnt receptor turnover in an R-spondin-sensitive manner. Nature 2012, 485, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Yu, Q.C.; Jiang, W.; Liu, W.; Song, W.; Yu, H.; Zhang, L.; Yang, Y.; Zeng, Y.A. R-spondin1 is a novel hormone mediator for mammary stem cell self-renewal. Genes Dev. 2014, 28, 2205–2218. [Google Scholar] [CrossRef] [PubMed]
- Chadi, S.; Buscara, L.; Pechoux, C.; Costa, J.; Laubier, J.; Chaboissier, M.C.; Pailhoux, E.; Vilotte, J.L.; Chanat, E.; Le Provost, F. R-spondin1 is required for normal epithelial morphogenesis during mammary gland development. Biochem. Biophys. Res. Commun. 2009, 390, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.A.; Kakitani, M.; Zhao, J.; Oshima, T.; Tang, T.; Binnerts, M.; Liu, Y.; Boyle, B.; Park, E.; Emtage, P.; et al. Mitogenic influence of human R-spondin1 on the intestinal epithelium. Science 2005, 309, 1256–1259. [Google Scholar] [CrossRef] [PubMed]
- Cadieu, E.; Neff, M.W.; Quignon, P.; Walsh, K.; Chase, K.; Parker, H.G.; Vonholdt, B.M.; Rhue, A.; Boyko, A.; Byers, A.; et al. Coat variation in the domestic dog is governed by variants in three genes. Science 2009, 326, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Snippert, H.J.; Haegebarth, A.; Kasper, M.; Jaks, V.; van Es, J.H.; Barker, N.; van de Wetering, M.; van den Born, M.; Begthel, H.; Vries, R.G.; et al. LGR6 marks stem cells in the hair follicle that generate all cell lineages of the skin. Science 2010, 327, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- Jaks, V.; Barker, N.; Kasper, M.; van Es, J.H.; Snippert, H.J.; Clevers, H.; Toftgard, R. LGR5 marks cycling, yet long-lived, hair follicle stem cells. Nat. Genet. 2008, 40, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Greco, V.; Chen, T.; Rendl, M.; Schober, M.; Pasolli, H.A.; Stokes, N.; Dela Cruz-Racelis, J.; Fuchs, E. A two-step mechanism for stem cell activation during hair regeneration. Cell Stem Cell 2009, 4, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Rendl, M.; Lewis, L.; Fuchs, E. Molecular dissection of mesenchymal-epithelial interactions in the hair follicle. PLoS Biol. 2005, 3, e331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Kawamura, K.; Takae, S.; Deguchi, M.; Yang, Q.; Kuo, C.; Hsueh, A.J. Oocyte-derived R-spondin2 promotes ovarian follicle development. FASEB J. 2013, 27, 2175–2184. [Google Scholar] [CrossRef] [PubMed]
- Ootani, A.; Li, X.; Sangiorgi, E.; Ho, Q.T.; Ueno, H.; Toda, S.; Sugihara, H.; Fujimoto, K.; Weissman, I.L.; Capecchi, M.R.; et al. Sustained in vitro intestinal epithelial culture within a Wnt-dependent stem cell niche. Nat. Med. 2009, 15, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Botchkarev, V.A.; Botchkareva, N.V.; Nakamura, M.; Huber, O.; Funa, K.; Lauster, R.; Paus, R.; Gilchrest, B.A. Noggin is required for induction of the hair follicle growth phase in postnatal skin. FASEB J. 2001, 15, 2205–2214. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Lei, X.; Zhang, X.; Zhang, H.; Liu, S.; Chen, Q.; Hu, H.; Wang, X.; Ning, L.; Cao, Y.; et al. Mtor signaling promotes stem cell activation via counterbalancing BMP-mediated suppression during hair regeneration. J. Mol. Cell Biol. 2015, 7, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Oshimori, N.; Fuchs, E. Paracrine TGF-β signaling counterbalances BMP-mediated repression in hair follicle stem cell activation. Cell Stem Cell 2012, 10, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Parma, P.; Radi, O.; Vidal, V.; Chaboissier, M.C.; Dellambra, E.; Valentini, S.; Guerra, L.; Schedl, A.; Camerino, G. R-spondin1 is essential in sex determination, skin differentiation and malignancy. Nat. Genet. 2006, 38, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Ishii, Y.; Wajid, M.; Bazzi, H.; Fantauzzo, K.A.; Barber, A.G.; Blaydon, D.C.; Nam, J.S.; Yoon, J.K.; Kelsell, D.P.; Christiano, A.M. Mutations in R-spondin 4 (RSPO4) underlie inherited anonychia. J. Investig. Dermatol. 2008, 128, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Blaydon, D.C.; Philpott, M.P.; Kelsell, D.P. R-spondins in cutaneous biology: Nails and cancer. Cell Cycle 2007, 6, 895–897. [Google Scholar] [CrossRef] [PubMed]
- Chua, A.W.; Ma, D.; Gan, S.U.; Fu, Z.; Han, H.C.; Song, C.; Sabapathy, K.; Phan, T.T. The role of R-spondin2 in keratinocyte proliferation and epidermal thickening in keloid scarring. J. Investig. Dermatol. 2011, 131, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.S.; Turcotte, T.J.; Yoon, J.K. Dynamic expression of R-spondin family genes in mouse development. Gene Expr. Patterns 2007, 7, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.R.; Turcotte, T.J.; Crocker, A.L.; Han, X.H.; Yoon, J.K. The canonical wnt signaling activator, R-spondin2, regulates craniofacial patterning and morphogenesis within the branchial arch through ectodermal-mesenchymal interaction. Dev. Biol. 2011, 352, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, M.; Porntaveetus, T.; Kawasaki, K.; Oommen, S.; Otsuka-Tanaka, Y.; Hishinuma, M.; Nomoto, T.; Maeda, T.; Takubo, K.; Suda, T.; et al. R-spondins/LGRs expression in tooth development. Dev. Dyn. 2014, 243, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.M.; Schreiner, C.M.; Wert, S.E.; Mucenski, M.L.; Scott, W.J.; Whitsett, J.A. R-spondin 2 is required for normal laryngeal-tracheal, lung and limb morphogenesis. Development 2008, 135, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Enshell-Seijffers, D.; Lindon, C.; Kashiwagi, M.; Morgan, B.A. β-catenin activity in the dermal papilla regulates morphogenesis and regeneration of hair. Dev. Cell 2010, 18, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Andl, T.; Bagasra, A.; Lu, M.M.; Epstein, D.J.; Morrisey, E.E.; Millar, S.E. Characterization of Wnt gene expression in developing and postnatal hair follicles and identification of Wnt5A as a target of sonic hedgehog in hair follicle morphogenesis. Mech. Dev. 2001, 107, 69–82. [Google Scholar] [CrossRef]
- Plikus, M.V. New activators and inhibitors in the hair cycle clock: Targeting stem cells’ state of competence. J. Investig. Dermatol. 2012, 132, 1321–1324. [Google Scholar] [CrossRef] [PubMed]
- Plikus, M.V.; Mayer, J.A.; de la Cruz, D.; Baker, R.E.; Maini, P.K.; Maxson, R.; Chuong, C.M. Cyclic dermal bmp signalling regulates stem cell activation during hair regeneration. Nature 2008, 451, 340–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lien, W.H.; Fuchs, E. Wnt some lose some: Transcriptional governance of stem cells by Wnt/β-catenin signaling. Genes Dev. 2014, 28, 1517–1532. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Qiao, J.; Zhang, S.; Zhang, H.; Lei, X.; Wang, X.; Deng, Z.; Ning, L.; Cao, Y.; Guo, Y.; et al. GPR39 marks specific cells within the sebaceous gland and contributes to skin wound healing. Sci. Rep. 2015, 5, 7913. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, X.; Zhao, Q.; Liu, S.; Zhang, H.; Shi, J.; Li, N.; Lei, X.; Zhao, H.; Deng, Z.; et al. Senescence of human skin-derived precursors regulated by Akt-FOXO3-p27 (KIP(1))/p15 (INK(4)b) signaling. Cell. Mol. Life Sci. 2015, 72, 2949–2960. [Google Scholar] [CrossRef] [PubMed]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, N.; Liu, S.; Zhang, H.-S.; Deng, Z.-L.; Zhao, H.-S.; Zhao, Q.; Lei, X.-H.; Ning, L.-N.; Cao, Y.-J.; Wang, H.-B.; et al. Exogenous R-Spondin1 Induces Precocious Telogen-to-Anagen Transition in Mouse Hair Follicles. Int. J. Mol. Sci. 2016, 17, 582. https://doi.org/10.3390/ijms17040582
Li N, Liu S, Zhang H-S, Deng Z-L, Zhao H-S, Zhao Q, Lei X-H, Ning L-N, Cao Y-J, Wang H-B, et al. Exogenous R-Spondin1 Induces Precocious Telogen-to-Anagen Transition in Mouse Hair Follicles. International Journal of Molecular Sciences. 2016; 17(4):582. https://doi.org/10.3390/ijms17040582
Chicago/Turabian StyleLi, Na, Shu Liu, Hui-Shan Zhang, Zhi-Li Deng, Hua-Shan Zhao, Qian Zhao, Xiao-Hua Lei, Li-Na Ning, Yu-Jing Cao, Hai-Bin Wang, and et al. 2016. "Exogenous R-Spondin1 Induces Precocious Telogen-to-Anagen Transition in Mouse Hair Follicles" International Journal of Molecular Sciences 17, no. 4: 582. https://doi.org/10.3390/ijms17040582
APA StyleLi, N., Liu, S., Zhang, H. -S., Deng, Z. -L., Zhao, H. -S., Zhao, Q., Lei, X. -H., Ning, L. -N., Cao, Y. -J., Wang, H. -B., Liu, S., & Duan, E. -K. (2016). Exogenous R-Spondin1 Induces Precocious Telogen-to-Anagen Transition in Mouse Hair Follicles. International Journal of Molecular Sciences, 17(4), 582. https://doi.org/10.3390/ijms17040582