
Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review

 ,
,  , and
, and
Abstract
:1. Introduction
2. Methodology
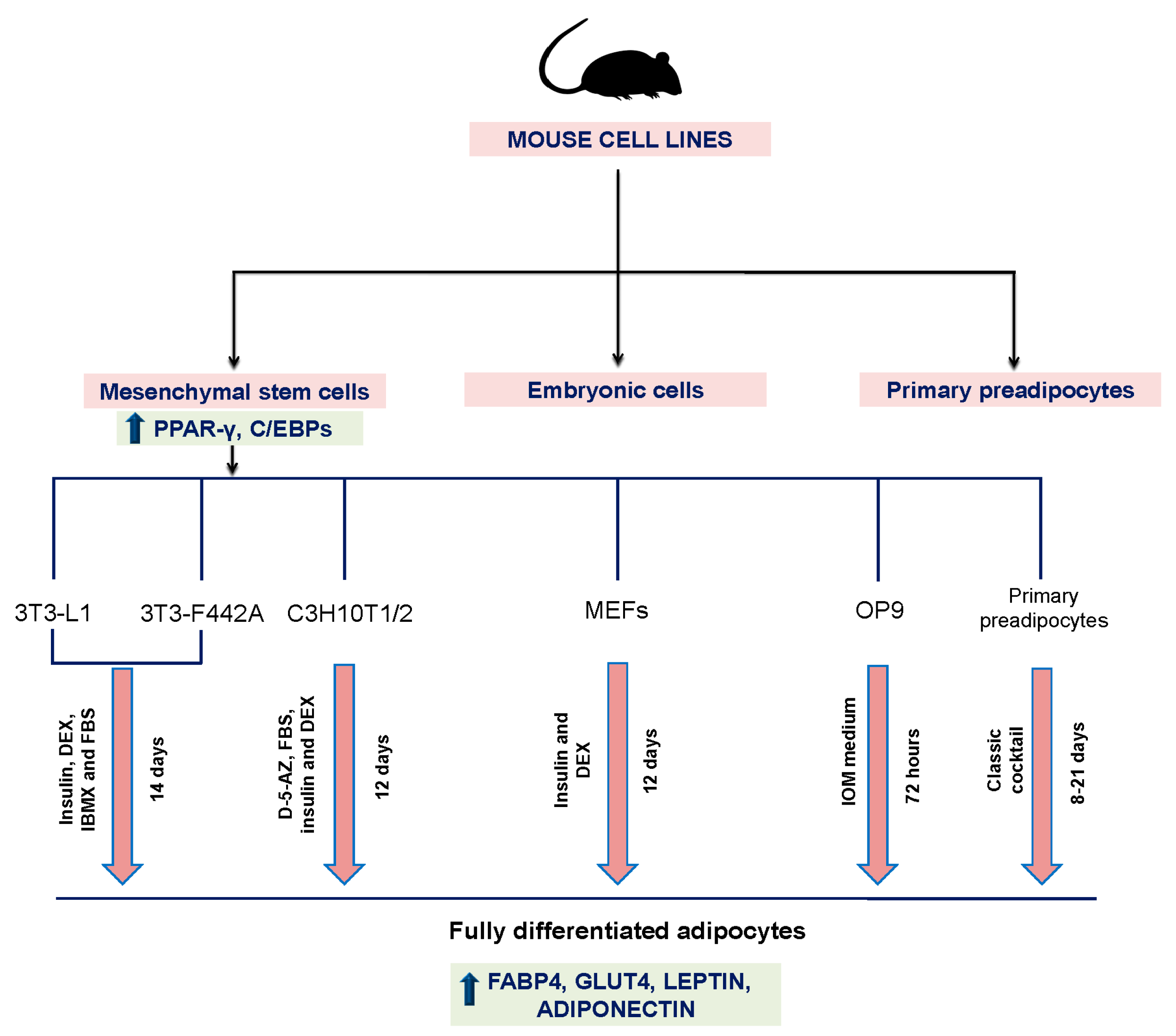
3. Animal Cell Models
3.1. 3T3-L1 Mouse Cell Line
3.2. 3T3-F442A Mouse Cell Line
3.3. OP9 Mouse Cell Line
3.4. C3H10T1/2 Mouse Cell Line
3.5. Primary Mouse Embryonic Fibroblasts (MEFs)
3.6. Porcine Primary Preadipocytes
3.7. Feline Primary Preadipocytes
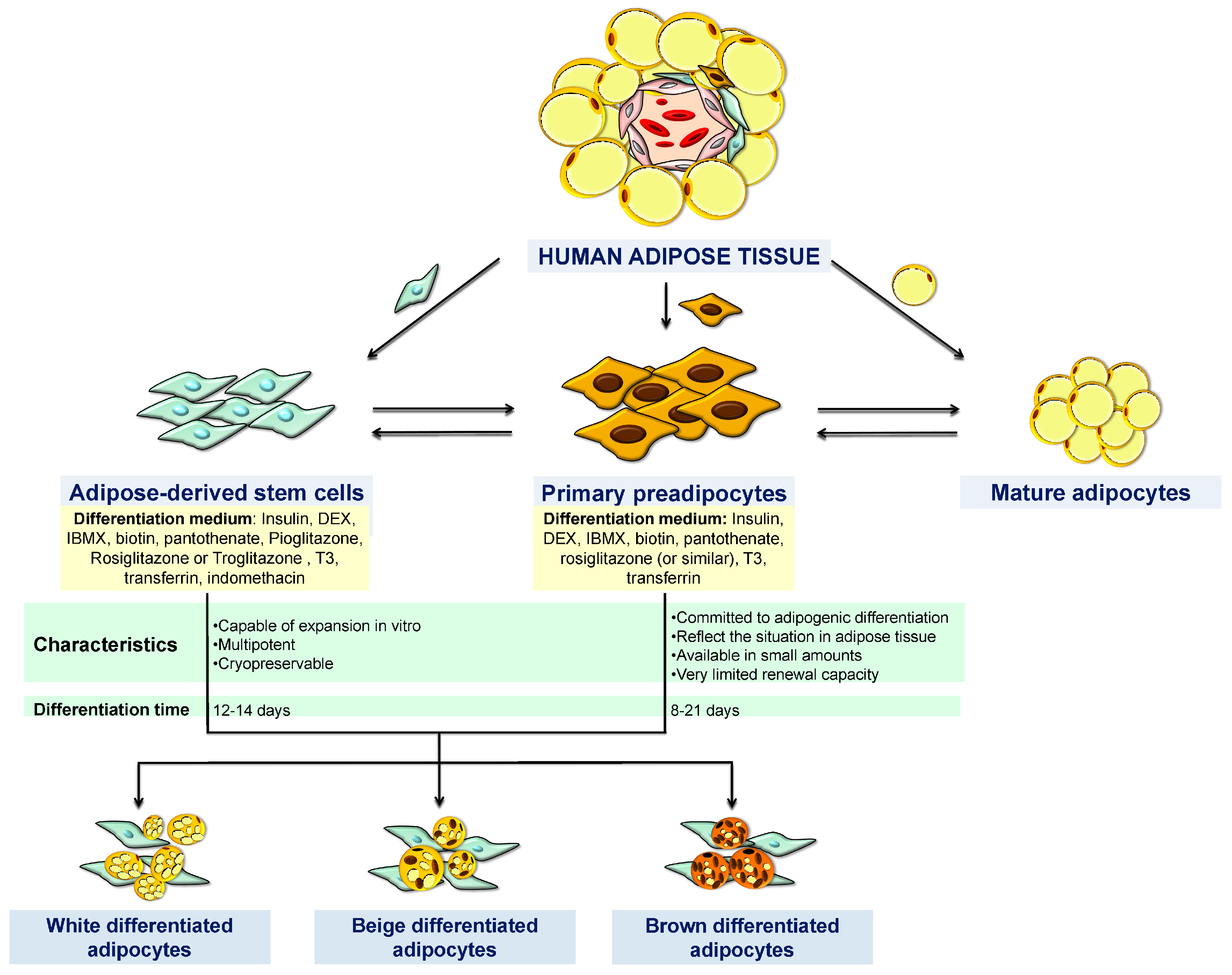
4. Human Cell Models
4.1. Adipose-Derived Stem Cells (ASCs)
4.2. Primary Preadipocytes
5. Brown/Beige Adipose Cell Lines
5.1. Primary Cell Models of Browning
5.2. Brown/Beige Differentiated Adipocytes
6. Cell Lines Representative of Various Diseases
6.1. Simpson-Golabi-Behemil Syndrome (SGBS) Cells
6.2. LiSa-2 Cells
7. Co-Cultures and Three-Dimensional Cultures (3D)
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, Q.A.; Scherer, P.E.; Gupta, R.K. Improved methodologies for the study of adipose biology: Insights gained and opportunities ahead. J. Lipid Res. 2014, 55, 605–624. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yao, W.; Jiang, H. Short-chain fatty acids enhance adipocyte differentiation in the stromal vascular fraction of porcine adipose tissue. J. Nutr. 2014, 144, 1887–1895. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Baraban, E.; Chavakis, T.; Hamilton, B.S.; Sales, S.; Wabitsch, M.; Bornstein, S.R.; Ehrhart-Bornstein, M. Anti-inflammatory properties of bone morphogenetic protein 4 in human adipocytes. Int. J. Obes. 2016, 40, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Wensveen, F.M.; Valentić, S.; Šestan, M.; TurkWensveen, T.; Polić, B. The “Big Bang” in obese fat: Events initiating obesity-induced adipose tissue inflammation. Eur. J. Immunol. 2015, 45, 2446–2456. [Google Scholar] [CrossRef] [PubMed]
- Boyer, W.R.; Johnson, T.M.; Fitzhugh, E.C.; Richardson, M.R.; Churilla, J.R. The associations between increasing degrees of homeostatic model assessment for insulin resistance and muscular strengthening activities among euglycaemic US adults. Diabetes Vasc. Dis. Res. 2015, 12, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Chen, P.; Sun, L. Regulatory networks of non-coding RNAs in brown/beige adipogenesis. Biosci. Rep. 2015, 35. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [PubMed]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Cell 2009, 20, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Saraf, N.; Sharma, P.K.; Mondal, S.C.; Vipin, K.G.; Singh, A.K. Role of PPARg2 transcription factor inthiazolidinedione-induced insulin sensitization. J. Pharm. Pharmacol. 2012, 64, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Fasshauer, M.; Klein, H.H.; Benito, M.; Kahn, C.R. Novel adipocyte lines from brown fat: A model system for the study of differentiation, energy metabolism, and insulin action. Bioessays 2002, 24, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Sorisky, A. From preadipocyte to adipocyte: Differentiation-directed signals of insulin from the cell surface to the nucleus. Crit. Rev. Clin. Lab. Sci. 1999, 36, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Dodson, M.V.; Vierck, J.L.; Hossner, K.L.; Byrne, K.; McNamara, J.P. The development and utility of a defined muscle and fat co-culture system. Tissue Cell 1997, 29, 517–524. [Google Scholar] [CrossRef]
- Armani, A.; Mammi, C.; Marzolla, V.; Calanchini, M.; Antelmi, A.; Rosano, G.M.; Fabbri, A.; Caprio, M. Cellular models for understanding adipogenesis, adipose dysfunction, and obesity. J. Cell. Biochem. 2010, 110, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Poulos, S.P.; Dodson, M.V.; Hausman, G.J. C ell line models for differentiation: Preadipocytes and adipocytes. Exp. Biol. Med. 2010, 35, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Fried, S.K. Optimal Protocol for the Differentiation and Metabolic Analysis of Human Adipose Stromal Cells. Methods Enzymol. 2014, 538, 49–65. [Google Scholar] [PubMed]
- Cristancho, A.G.; Lazar, M.A. Forming functional fat: A growing understanding of adipocyte differentiation. Nat. Rev. Mol. Cell Biol. 2011, 12, 722–734. [Google Scholar] [CrossRef] [PubMed]
- Hausman, G.J.; Basu, U.; Wei, S.; Hausman, D.B.; Dodson, M.V. Preadipocyte and Adipose Tissue Differentiation in Meat Animals: Influence of Species and Anatomical Location Annu. Rev. Anim. Biosci. 2014, 2, 323–351. [Google Scholar] [CrossRef] [PubMed]
- Wolins, N.E.; Quaynor, B.K.; Skinne, J.R. OP9 mouse stromal cells rapidly differentiate into adipocytes: Characterization of a useful new model of adipogenesis. J. Lipid Res. 2006, 47, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Green, H.; Meuth, M. An established pre-adipose cell line and its differentiation in culture. Cell 1974, 3, 127–133. [Google Scholar] [CrossRef]
- Green, H.; Kehinde, O. Spontaneous heritable changes leading to increased adipose conversion in 3T3 cells. Cell 1976, 7, 105–113. [Google Scholar] [CrossRef]
- Vishwanath, D.; Srinivasan, H.; Patil, M.S.; Seetarama, S.; Kumar, S.A.; Dixit, M.N. Novel method to differentiate 3T3 L1 cells in vitro to produce highly sensitive adipocytes for a GLUT4 mediated glucose uptake using fluorescent glucose analog. J. Cell Commun. Signal. 2013, 7, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Zebisch, K.; Voigt, V.; Wabitsch, M.; Brandsch, M. Protocol for effective differentiation of 3T3-L1 cells to adipocytes. Anal. Biochem. 2012, 425, 88–90. [Google Scholar] [CrossRef] [PubMed]
- Reznikoff, C.A.; Brankow, D.W.; Heidelberger, C. Establishment and characterization of a cloned line of C3Hmouseembryo cells sensitive to post confluence inhibition of division. Cancer Res. 1973, 33, 3231–3238. [Google Scholar] [PubMed]
- Fei, Z.; Bera, T.K.; Liu, X.; Xiang, L.; Pastan, I. Ankrd26 Gene Disruption Enhances Adipogenesis of Mouse Embryonic Fibroblasts. J. Biol. Chem. 2011, 286, 27761–27768. [Google Scholar] [CrossRef] [PubMed]
- Garfield, A.S. Derivation of Primary Mouse Embryonic Fibroblast (PMEF) Cultures. Methods Mol. Biol. 2010, 633, 19–27. [Google Scholar] [PubMed]
- Caprio, M.; Fève, B.; Claës, A.; Viengchareun, S.; Lombès, M.; Zennaro, M.C. Pivotal role of the mineralocorticoid receptor in corticosteroid-induced adipogenesis. FASEB J. 2007, 21, 2185–2194. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.S.; Chen, Y.Y.; Lee, P.S.; Kalyanam, N.; Ho, C.T.; Liou, W.S. Bisdemethoxycurcumin Inhibits Adipogenesis in 3T3-L1 Preadipocytes and Suppresses Obesity in High-Fat Diet-Fed C57BL/6 Mice. J. Agric. Food Chem. 2016, 64, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.C.; Kang, N.; Ko, S.C.; Kim, Y.B.; Jeon, Y.J. Anti-obesity effects of seaweeds of Jeju Island on the differentiation of 3T3-L1 preadipocytes and obese mice fed a high-fat diet. Food Chem. Toxicol. 2016, 90, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Tutino, V.; Orlando, A.; Russo, F.; Notarnicola, M. Hydroxytyrosol Inhibits Cannabinoid CB1 Receptor Gene Expression in 3T3-L1Preadipocyte Cell Line. J. Cell. Physiol. 2016, 231, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Okabe, Y.; Shimada, T.; Horikawa, T.; Kinoshita, K.; Koyama, K.; Ichinose, K.; Aburada, M.; Takahashi, K. Suppression of adipocyte hypertrophy by polymethoxyflavonoids isolated from Kaempferia parviflora. Phytomedicine 2014, 21, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Eseberri, I.; Miranda, J.; Lasa, A.; Churruca, I.; Portillo, M.P. Doses of Quercetin in the Range of Serum Concentrations Exert Delipidating Effects in 3T3-L1 Preadipocytes by Acting on Different Stages of Adipogenesis, but Not in Mature Adipocytes. Oxid. Med. Cell. Longev. 2015, 2015, 480943. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Apostolatos, A.; Carter, G.; Ajmo, J.; Gali, M.; Cooper, D.R.; You, M.; Bisht, K.S.; Patel, N.A. Protein kinase C δ (PKCδ) splice variants modulate apoptosis pathway in 3T3L1cells during adipogenesis: Identification of PKCδII inhibitor. J. Biol. Chem. 2013, 288, 26834–26846. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Lin, K.Y.; Peng, K.Y.; Day, Y.J.; Hung, L.M. Resveratrol exerts anti-obesity effects in high-fat diet obese mice and displays differential dosage effects on cytotoxicity, differentiation, and lipolysis in 3T3-L1 cells. Endocr. J. 2015, 63, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Tanaka, G.; Masuda, S.; Ogasawara, J.; Sakurai, T.; Kizaki, T. Melatonin promotes adipogenesis and mitochondrial biogenesis in 3T3-L1preadipocytes. J. Pineal. Res. 2015, 59, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Calzadilla, P.; Gómez-Serrano, M.; García-Santos, E.; Schiappacasse, A.; Abalde, Y.; Calvo, J.C.; Peral, B.; Guerra, L.N. N-Acetylcysteine affects obesity-related protein expression in 3T3-L1 adipocytes. Redox. Rep. 2013, 18, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, H.; Kondo, Y.; Kawasaki, T.; Imamura, N. Cineromycin B isolated from Streptomyces cinerochromogenes inhibits adipocyte differentiation of 3T3-L1 cells via Krüppel-like factors 2 and 3. Life Sci. 2015, 135, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Artaza, J.N.; Taylor, W.E.; Gonzalez-Cadavid, N.F.; Bhasin, S. Androgens stimulate myogenic differentiation and inhibit adipogenesis in C3H10T1/2 pluripotent cells through an androgen receptor-mediated pathway. Endocrinology 2003, 144, 5081–5088. [Google Scholar] [CrossRef] [PubMed]
- Mammi, C.; Marzolla, V.; Armani, A.; Feraco, A.; Antelmi, A.; Maslak, E. A novel combined glucocorticoid-mineralocorticoid receptor selective modulator markedly prevents weight gain and fat mass expansion in mice fed a high-fat diet. Int. J. Obes. 2016. [Google Scholar] [CrossRef] [PubMed]
- Regnier, S.M.; El-Hashani, E.; Kamau, W.; Zhang, X.; Massad, N.L.; Sargis, R.M. Tributyltin differentially promotes development of a phenotypically distinct adipocyte. Obesity 2015, 23, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Abdesselem, H.; Madani, A.; Hani, A.; Al-Noubi, M.; Goswami, N.; Ben Hamidane, H.; Billing, A.M.; Pasquier, J.; Bonkowski, M.S.; Halabi, N.; et al. SIRT1 Limits Adipocyte Hyperplasia through c-Myc Inhibition. J. Biol. Chem. 2016, 291, 2119–2135. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Ding, W.; Wang, J.; Wu, G.; Zhang, H.; Yin, J.; Zhou, L.; Li, D. LOC66273 isoform 2, a novel protein highly expressed in white adipose tissue, induces adipogenesis in 3T3-L1 cells. J. Nutr. 2012, 142, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Lien, C.C.; Jiang, J.L.; Jian, D.Y.; Kwok, C.F.; Ho, L.T.; Juan, C.C. Chronic endothelin-1 infusion causes adipocyte hyperplasia in rats. Obesity 2016, 24, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Lee da, S.; Choi, H.; Han, B.S.; Kim, W.K.; Lee, S.C.; Oh, K.J. c-Jun regulates adipocyte differentiation via the KLF15-mediated mode. Biochem. Biophys. Res. Commun. 2016, 469, 552–558. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Pang, Y.W.; Gao, H.M.; Tao, L.; Miao, K.; Wu, Z.H.; Tian, J.H. Heterologous expression of C. elegans Fat-1 decreases the N-6/N-3 fatty acid ratio and inhibits adipogenesis in 3T3-L1 cells. Biochem. Biophys. Res. Commun. 2012, 428, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Yun, U.J.; Song, N.J.; Yang, D.K.; Kwon, S.M.; Kim, K.; Kim, S. miR-195a inhibits adipocyte differentiation by targeting the preadipogenic determinator Zfp423. J. Cell. Biochem. 2015, 116, 2589–2597. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Chang, W.T.; Wu, S.J.; Xu, P.Y.; Ting, N.C.; Liou, C.J. Phloretin and phlorizin promote lipolysis and inhibit inflammation in mouse 3T3-L1 cells and in macrophage-adipocyte co-cultures. Mol. Nutr. Food Res. 2013, 57, 1803–1813. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.A.; Tang, Y.; Weiss, S.J.; Janorkar, A.V. Three-dimensional spheroid cell model of in vitro adipocyte inflammation. Tissue Eng. Part A 2015, 21, 1837–1847. [Google Scholar] [CrossRef] [PubMed]
- Student, A.K.; Hsu, R.Y.; Lane, M.D. Induction of fatty acid synthetase synthesis in differentiating 3T3-L1 preadipocytes. J. Biol. Chem. 1980, 255, 4745–4750. [Google Scholar] [PubMed]
- Hernández-Mosqueira, C.; Velez-delValle, C.; Kuri-Harcuch, W. Tissue alkaline phosphatase is involved in lipid metabolism and gene expression and secretion of adipokines in adipocytes. Biochim. Biophys. Acta 2015, 1850, 2485–2496. [Google Scholar] [CrossRef] [PubMed]
- Desarzens, S.; Liao, W.H.; Mammi, C.; Caprio, M.; Faresse, N. Hsp90 blockers inhibit adipocyte differentiation and fat mass accumulation. PLoS ONE 2014, 9, e94127. [Google Scholar] [CrossRef] [PubMed]
- Scroyen, I.; Bauters, D.; Vranckx, C.; Lijnen, H.R. The Anti-Adipogenic Potential of COUP-TFII is Mediated by down regulation of the Notch Target Gene Hey1. PLoS ONE 2015, 10, e0145608. [Google Scholar] [CrossRef] [PubMed]
- Lane, J.M.; Doyle, J.R.; Fortin, J.P.; Kopin, A.S.; Ordovás, J.M. Development of an OP9 derived cell line as a robust model to rapidly study adipocyte differentiation. PLoS ONE 2014, 9, e112123. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Kodama, H.; Honjo, T. Generation of lymphohematopoietic cells from embryonic stem cells in culture. Science 1994, 265, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Gerhard Vogel, H. Drug Discovery and Evaluation Pharmacological Assays, 3rd ed.; Springer Verlag Berlin: Heidelberg, Germany, 2008; pp. 1379–1381. [Google Scholar]
- Seo, Y.S.; Kang, O.H.; Kim, S.B.; Mun, S.H.; Kang, D.H.; Yang, D.W.; Choi, J.G.; Lee, Y.M.; Kang, D.K.; Lee, H.S.; et al. Quercetin prevents adipogenesis by regulation of transcriptional factors and lipases in OP9 cells. Int. J. Mol. Med. 2015, 35, 1779–1785. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Kim, J.M.; Kim, M.S.; Hwang, J.K.; Yang, S.H.; Kim, H.J.; Lee, D.S.; Oh, H.; Kim, Y.C.; Ryu, D.G.; et al. Inhibitory effects of Pericarpium zanthoxyli extract on adipocyte differentiation. Int. J. Mol. Med. 2014, 33, 1140–1146. [Google Scholar] [CrossRef] [PubMed]
- Rahman, F.; Al Frouh, F.; Bordignon, B.; Fraterno, M.; Landrier, J.F.; Peiretti, F.; Fontes, M. Ascorbic acid is a dose-dependent inhibitor of adipocyte differentiation, probably by reducing cAMP pool. Front. Cell Dev. Biol. 2014, 7, 2–29. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Aoshima, H.; Saitoh, Y.; Miwa, N. Highly hydroxylated fullerene localizes at the cytoskeleton and inhibits oxidative stress in adipocytes and a subcutaneous adipose-tissue equivalent. Free. Radic. Biol. Med. 2011, 51, 1376–1389. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, Y.; Mizuno, H.; Xiao, L.; Hyoudou, S.; Kokubo, K.; Miwa, N. Polyhydroxylated fullerene C₆₀(OH)₄₄ suppresses intracellular lipid accumulation together with repression of intracellular superoxide anion radicals and subsequent PPARγ2 expression during spontaneous differentiation of OP9preadipocytes into adipocytes. Mol. Cell. Biochem. 2012, 366, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Kim, I.; Park, S.; Han, D.; Ha, S.; Kwon, M. Creatine inhibits adipogenesis by downregulating insulin-induced activation of the phosphatidylinositol 3-kinase signaling pathway. Stem Cells Dev. 2015, 24, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Beg, M.; Chauhan, P.; Varshney, S.; Shankar, K.; Rajan, S.; Saini, D. A withanolide coagulin-L inhibits adipogenesis modulating Wnt/β-catenin pathway and cell cycle in mitotic clonal expansion. Phytomedicine 2014, 21, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Biemann, R.; Fischer, B.; Blüher, M.; Navarrete Santos, A. Tributyltin affects adipogenic cell fate commitment in mesenchymal stem cells by a PPARγ independent mechanism. Chem. Biol. Interact. 2014, 214, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Bhasin, S.; Braga, M.; Artaza, J.N.; Pervin, S.; Taylor, W.E.; Krishnan, V.; Sinha, S.K.; Rajavashisth, T.B.; Jasuja, R. Regulation of myogenic differentiation by androgens: Cross talk between androgen receptor/β-catenin and follistatin/transforming growth factor-β signaling pathways. Endocrinology 2009, 150, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Chatterjee, S.; Li, L.; Kim, J.M.; Lee, J.; Yechoor, VK. The clock gene, brain and muscle Arnt-like 1, regulates adipogenesis via Wnt signaling pathway. FASEB J. 2012, 26, 3453–3463. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Wan, Y.; Zhang, S.; Zhang, Q.; Ye, H.; Li, Y. Role of bone morphogenetic protein 4 in the differentiation of brown fat-like adipocytes. Am. J. Physiol. Endocrinol. Metab. 2014, 306, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Rosen, ED.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Merkestein, M.; Laber, S.; McMurray, F.; Andrew, D.; Sachse, G.; Sanderson, J.; Li, M.; Usher, S.; Sellayah, D.; Ashcroft, F.M.; et al. FTO influences adipogenesis by regulating mitotic clonal expansion. Nat. Commun. 2015, 6, 6792. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Murthy, R.; Wood, B.; Song, B.; Wang, S.; Sun, B.; Malhi, H.; Kaufman, R.J. ER stress signaling through eIF2α and CHOP, but not IRE1α, attenuates adipogenesis in mice. Diabetologia 2013, 56, 911–924. [Google Scholar] [CrossRef] [PubMed]
- Hee, S.W.; Tsai, S.H.; Chang, Y.C.; Chang, C.J.; Yu, I.S.; Lee, P.C.; Lee, W.J.; Yun-Chia Chang, E.; Chuang, L.M. The role of nocturnin in early adipogenesis and modulation of systemic insulin resistance in human. Obesity 2012, 20, 1558–1565. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, K.W.; Lee, E.W.; Jang, W.S.; Seo, J.; Shin, S.; Hwang, K.A.; Song, J. Suppression of PPARγ through MKRN1-mediated ubiquitination and degradation prevents adipocyte differentiation. Cell. Death Differ. 2014, 21, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Braga, M.; Reddy, S.T.; Vergnes, L.; Pervin, S.; Grijalva, V.; Stout, D.; David, J.; Li, X.; Tomasian, V.; Reid, C.B. Follistatin promotes adipocyte differentiation, browning, and energy metabolism. J. Lipid Res. 2014, 55, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.; Wang, Y.; Wei, N.; Xu, R.; Xiong, Y.; Wang, P.; Shen, Q.; Yang, G. Sirt1 inhibits Akt2-mediated porcine adipogenesis potentially by direct protein-protein interaction. PLoS ONE 2013, 8, e71576. [Google Scholar] [CrossRef] [PubMed]
- Shu, G.; Lu, N.S.; Zhu, X.T.; Xu, Y.; Du, M.Q.; Xie, Q.P.; Zhu, C.J.; Xu, Q.; Wang, S.B.; Wang, L.N.; et al. Phloretin promotes adipocyte differentiation in vitro and improves glucose homeostasis in vivo. J. Nutr. Biochem. 2014, 25, 1296–1308. [Google Scholar] [CrossRef] [PubMed]
- Bohan, A.E.; Purvis, K.N.; Bartosh, J.L.; Brandebourg, T.D. The proliferation and differentiation of primary pig preadipocytes is suppressed when cultures are incubated at 37° Celsius compared to euthermic conditions in pigs. Adipocyte 2014, 3, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Song, Z.Y.; Pu, L.; Yang, H.; Zheng, J.M.; Zhang, Z.Y.; Shi, X.E.; Yang, G.S. Retinol binding protein 4 affects the adipogenesis of porcine preadipocytes through insulin signaling pathways. Biochem. Cell Biol. 2013, 91, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.J.; Wei, N.; Wang, Y.; Xiong, Y.; Chen, F.F.; Wu, W.J.; Zhao, C.Z.; Sun, S.D.; Yang, G.S. Obese and lean porcine difference of FoxO1 and its regulation through C/EBPβ and PI3K/GSK3β signaling pathway. J. Anim. Sci. 2014, 92, 1968–1979. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.L.; Song, C.C.; Li, Y.F.; He, J.J.; Li, Y.L.; Zheng, X.L.; Yang, G.S. miR-125a inhibits porcine preadipocytes differentiation by targeting ERRα. Mol. Cell. Biochem. 2014, 395, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.E.; Li, Y.F.; Jia, L.; Ji, H.L.; Song, Z.Y.; Cheng, J.; Wu, G.F.; Song, C.C.; Zhang, Q.L.; Zhu, J.Y.; et al. MicroRNA-199a-5p affects porcine preadipocyte proliferation and differentiation. Int. J. Mol. Sci. 2014, 15, 8526–8538. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, X.; Guan, L.; Qi, Q.; Shu, G.; Jiang, Q.; Yuan, L.; Xi, Q.; Zhang, Y. MiRNA-181a regulates adipogenesis by targeting tumor necrosis factor-α (TNF-α) in the porcine model. PLoS ONE 2013, 8, e71568. [Google Scholar]
- Mai, Y.; Zhang, Z.; Yang, H.; Dong, P.; Chu, G.; Yang, G.; Sun, S. BMP and activin membrane-bound inhibitor (BAMBI) inhibits the adipogenesis of porcine preadipocytes through Wnt/β-catenin signaling pathway. Biochem. Cell Biol. 2014, 92, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Riedel, J.; Badewien-Rentzsch, B.; Kohn, B.; Hoeke, L.; Einspanier, R. Characterization of key genes of the renin–angiotensin system in mature feline adipocytes and during in vitro adipogenesis. J. Anim. Physiol. Anim. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Esteve Ràfols, M. Adipose tissue: Cell heterogeneity and functional diversity. Endocrinol. Nutr. 2014, 61, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Cawthorn, W.P.; Scheller, E.L.; MacDougald, O.A. Adipose tissue stem cells meet preadipocyte commitment: Going back to the future. J. Lipid Res. 2012, 53, 227–246. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.R.; Clark, E.L. Microscopic studies of the new formation of fat in living adult rabbits. Am. J. Anat. 1940, 67, 255–285. [Google Scholar] [CrossRef]
- Huang, X.; Ordemann, J.; Müller, J.M.; Dubiel, W. The COP9 signalosome, cullin 3 and Keap1 super complex regulates CHOP stability and adipogenesis. Biol. Open 2012, 1, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived Stem Cells: Isolation, Expansion and Differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Yong, K.W.; Pingguan-Murphy, B.; Xu, F.; Abas, W.A.; Choi, J.R.; Omar, S.Z.; Azmi, M.A.; Chua, K.H.; Wan Safwani, W.K. Phenotypic and functional characterization of long-term cryopreserved human adipose-derived stem cells. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Diascro, D.D., Jr.; Vogel, R.L.; Johnson, T.E.; Witherup, K.M.; Pitzenberger, S.M.; Rutledge, S.J.; Prescott, D.J.; Rodan, G.A.; Schmidt, A. High fatty acid content in rabbit serum is responsible for the differentiation of osteoblasts into adipocyte-like cells. J. Bone Miner. Res. 1998, 13, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Madsen, L.; Petersen, R.K.; Techer, N.; Kopperud, R.; Ma, T.; Døskeland, S.O.; Ailhaud, G.; Wang, J.; Amri, E.Z.; et al. Activation of protein kinase A and exchange protein directly activated by cAMP promotes adipocyte differentiation of human mesenchymal stem cells. PLoS ONE 2012, 7, e34114. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.; Kim, Y.; Tomás-Barberán, F.A.; Espín, J.C.; Chung, S. 1-Urolithin A, C and D, but not iso-Urolithin A and Urolithin B, attenuate triglyceride accumulation in human cultures of adipocytes and hepatocytes. Mol. Nutr. Food Res. 2016, 60, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Yagiz, Y.; Xu, C.; Lu, J.; Chung, S.; Marshall, M.R. Muscadine grape seed oil as a novel source of tocotrienols to reduce adipogenesis and adipocyte inflammation. Food Funct. 2015, 6, 2293–2302. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Gu, Y.; Liu, G.; Zhang, F.; Li, J.; Liu, F.; Zhang, Z.; Ye, J.; Li, Q. Cidec promotes the differentiation of human adipocytes by degradation of AMPKα through ubiquitin-proteasome pathway. Biochim. Biophys. Acta 2015, 1850, 2552–2562. [Google Scholar] [CrossRef] [PubMed]
- Roca-Rodríguez, M.M.; El Bekay, R.; Garrido-Sanchez, L.; Gómez-Serrano, M.; Coin-Aragüez, L.; Oliva-Olivera, W.; Lhamyani, S.; Clemente-Postigo, M.; García-Santos, E.; de Luna Diaz, R.; et al. Parathyroid Hormone-Related Protein, Human Adipose-Derived Stem Cells Adipogenic Capacity and Healthy Obesity. J. Clin. Endocrinol. Metab. 2015, 100, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Narvaez, C.J.; Simmons, K.M.; Brunton, J.; Salinero, A.; Chittur, S.V.; Welsh, J.E. Induction of STEAP4 correlates with 1,25-dihydroxyvitamin D3 stimulation of adipogenesis in mesenchymal progenitor cells derived from human adipose tissue. J. Cell. Physiol. 2013, 228, 2024–2036. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Dusting, G.J.; Peshavariya, H.; Jiang, F.; Hsiao, S.T.; Chan, E.C.; Liu, G.S. Differentiation of human adipose-derived stem cells into fat involves reactive oxygen species and Forkhead box O1 mediated upregulation of antioxidant enzymes. Stem Cells Dev. 2013, 22, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ojeda, F.J.; Gomez-Llorente, C.; Aguilera, C.M.; Gil, A.; Rupérez, A.I. Impact of 3 Amino-1,2,4-Triazole (3-AT)-Derived Increase in Hydrogen Peroxide Levels on Inflammation and Metabolism in Human Differentiated Adipocytes. PLoS ONE 2016, 11, e0152550. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ojeda, F.J.; Aguilera, C.M.; Rupérez, A.I.; Gil, Á.; Gomez-Llorente, C. An analogue of atrial natriuretic peptide (C-ANP4–23) modulates glucose metabolism in human differentiated adipocytes. Mol. Cell. Endocrinol. 2016, 431, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Shi, C.M.; Chen, L.; Pang, L.X.; Xu, G.F.; Gu, N.; Zhu, L.J.; Guo, X.R.; Ni, Y.H.; Ji, C.B. The biological effects of hsa-miR-1908 in human adipocytes. Mol. Biol. Rep. 2015, 42, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Pisani, D.F.; Djedaini, M.; Beranger, G.E.; Elabd, C.; Scheideler, M.; Ailhaud, G.; Amri, E.Z. Differentiation of Human Adipose-Derived Stem Cells into “Brite” (Brown-in-White) Adipocytes. Front. Endocrinol. 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Molchadsky, A.; Ezra, O.; Amendola, P.G.; Krantz, D.; Kogan-Sakin, I.; Buganim, Y.; Rivlin, N.; Goldfinger, N.; Folgiero, V.; Falcioni, R.; et al. p53 is required for brown adipogenic differentiation and has a protective role against diet-induced obesity. Cell Death. Differ. 2013, 20, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Lessard, J.; Laforest, S.; Pelletier, M.; Leboeuf, M.; Blackburn, L.; Tchernof, A. Low abdominal subcutaneous preadipocyte adipogenesis is associated with visceral obesity, visceral adipocyte hypertrophy, and a dysmetabolic state. Adipocyte 2014, 3, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Michaud, A.; Lacroix-Pépin, N.; Pelletier, M.; Daris, M.; Biertho, L.; Fortier, M.A.; Tchernof, A. Expression of genes related to prostaglandin synthesis or signaling in human subcutaneous and omental adipose tissue: Depot differences and modulation by adipogenesis. Mediators Inflamm. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Park, H.T.; Lee, E.S.; Cheon, Y.P.; Lee, D.R.; Yang, K.S.; Kim, Y.T.; Hur, J.Y.; Kim, S.H.; Lee, K.W.; Kim, T. The relationship between fat depot-specific preadipocyte differentiation and metabolic syndrome in obese women. Clin. Endocrinol. 2012, 76, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.L.; Vorlander, C.; Reddy-Luthmoodoo, M.; Natoli, A.K.; Formosa, M.F.; Bertovic, D.A.; Anderson, M.J.; Duffy, S.J.; Kingwell, B.A. Reduced UCP-1 content in in vitro differentiated beige/brite adipocytes derived from preadipocytes of human subcutaneous white adipose tissues in obesity. PLoS ONE 2014, 9, e91997. [Google Scholar] [CrossRef] [PubMed]
- Rossmeislová, L.; Malisová, L.; Kracmerová, J.; Tencerová, M.; Kovácová, Z.; Koc, M.; Siklová-Vítková, M.; Viquerie, N.; Langin, D.; Stich, V. Weight loss improves the adipogenic capacity of human preadipocytes and modulates their secretory profile. Diabetes 2013, 62, 1990–1995. [Google Scholar] [CrossRef] [PubMed]
- Darimont, C.; Macé, K. Immortalization of human preadipocytes. Biochimie 2003, 85, 1231–1233. [Google Scholar] [CrossRef] [PubMed]
- Church, C.; Brown, M.; Rodeheffer, M.S. Conditional immortalization of primary adipocyte precursor cells. Adipocyte 2015, 4, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Wu, Y.; Fried, S.K. A modified protocol to maximize differentiation of human preadipocytes and improve metabolic phenotypes. Obesity 2012, 20, 2334–2340. [Google Scholar] [CrossRef] [PubMed]
- Divoux, A.; Karastergiou, K.; Xie, H.; Guo, W.; Perera, R.J. Fried, S.K.; Smith, S.R. Identification of a novel lncRNA in gluteal adipose tissue and evidence for its positive effect on preadipocyte differentiation. Obesity 2014, 22, 1781–175. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; Gavrilovic, J.; Turner, J. Effect of tissue inhibitor of metalloproteinases 3 on DLK1 shedding in cultured human pre-adipocytes and implications for adipose tissue remodelling. Lancet 2015, 385, S35. [Google Scholar] [CrossRef]
- Gustafson, B.; Hammarstedt, A.; Hedjazifar, S.; Hoffmann, J.M.; Svensson, P.A.; Grimsby, J.; Rondinone, C.; Smith, U. BMP4 and BMP Antagonists Regulate Human White and Beige Adipogenesis. Diabetes 2015, 64, 1670–1681. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Pickering, R.T.; Puri, V. Prolonged efficiency of siRNA-mediated gene silencing in primary cultures of human preadipocytes and adipocytes. Obesity 2014, 22, 1064–1069. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Serrano, M.; Sabater, M.; Ortega, F.; Serino, M.; Pueyo, N.; Luche, E.; Waget, A.; Rodriguez-Hermosa, J.I.; Ricart, W. Study of lactoferrin gene expression in human and mouse adipose tissue, human preadipocytes and mouse 3T3-L1 fibroblasts. Association with adipogenic and inflammatory markers. J. Nutr. Biochem. 2013, 24, 1266–1275. [Google Scholar] [CrossRef] [PubMed]
- Söhle, J.; Machuy, N.; Smailbegovic, E.; Holtzmann, U.; Grönniger, E.; Wenck, H.; Stäb, F.; Winnefeld, M. Identification of new genes involved in human adipogenesis and fat storage. PLoS ONE 2012, 7, e31193. [Google Scholar] [CrossRef] [PubMed]
- Veilleux, A.; Côté, J.A.; Blouin, K.; Nadeau, M.; Pelletier, M.; Marceau, P.; Laberge, P.Y.; Luu-The, V.; Tchernof, A. Glucocorticoid-induced androgen inactivation by aldo-keto reductase 1C2 promotes adipogenesis in human preadipocytes. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E941–E949. [Google Scholar] [CrossRef] [PubMed]
- Berti, L.; Irmler, M.; Zdichavsky, M.; Meile, T.; Böhm, A.; Stefan, N.; Fritsche, A.; Beckers, J.; Königsrainer, A.; Häring, H.U.; et al. Fibroblast growth factor 21 is elevated in metabolically unhealthy obesity and affects lipid deposition, adipogenesis, and adipokine secretion of human abdominal subcutaneous adipocytes. Mol. Metab. 2015, 4, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Famulla, S.; Schlich, R.; Sell, H.; Eckel, J. Differentiation of human adipocytes at physiological oxygen levels results in increased adiponectin secretion and isoproterenol-stimulated lipolysis. Adipocyte 2012, 1, 132–181. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Ji, C.; Song, G.; Shi, C.; Shen, Y.; Chen, L.; Yang, L.; Zhao, Y.; Guo, X. Obesity-associated microRNA-26b regulates the proliferation of human preadipocytes via arrest of the G1/S transition. Mol. Med. Rep. 2015, 12, 3648–3654. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Xu, G.; Ji, C.; Shi, C.; Shen, Y.; Chen, L.; Zhu, L.; Yang, L.; Zhao, Y.; Guo, X. The role of microRNA-26b in human adipocyte differentiation and proliferation. Gene 2014, 533, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Dai, Y.M.; Ji, C.B.; Yang, L.; Shi, C.M.; Xu, G.F.; Pang, L.X.; Huang, F.Y.; Zhang, C.M.; Guo, X.R. MiR-146b is a regulator of human visceral preadipocyte proliferation and differentiation and its expression is altered in human obesity. Mol. Cell. Endocrinol. 2014, 393, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, X.; Ding, X.; Wang, H.; Chen, X.; Zhao, H.; Jia, Y.; Liu, S.; Liu, Y. miR-27 inhibits adipocyte differentiation via suppressing CREB expression. Acta Biochim. Biophys. 2014, 46, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.G.; Boudreau, A.; Atlas, E. Bisphenol A induces differentiation of human preadipocytes in the absence of glucocorticoid and is inhibited by an estrogen-receptor antagonist. Nutr. Diabetes 2014, 4, e102. [Google Scholar] [CrossRef] [PubMed]
- Ellero-Simatos, S.; Claus, S.P.; Benelli, C.; Forest, C.; Letourneur, F.; Cagnard, N.; Beaune, P.H.; de Waziers, I. Combined transcriptomic-(1)H NMR metabonomic study reveals that monoethylhexyl phthalate stimulates adipogenesis and glyceroneogenesis in human adipocytes. J. Proteome Res. 2011, 10, 5493–5502. [Google Scholar] [CrossRef] [PubMed]
- You, J.S.; Lee, Y.J.; Kim, K.S.; Kim, S.H.; Chang, K.J. Anti-obesity and hypolipidaemic effects of Nelumbo nucifera seed ethanol extract in human pre-adipocytes and rats fed a high-fat diet. J. Sci. Food Agric. 2014, 94, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Leong, J.; Ye, W.; Xu, P.; Lin, S.H.; Liu, J.Y.; Lin, Y.C. (−)-Gossypol-enriched cottonseed oil inhibits proliferation and adipogenesis of human breast pre-adipocytes. Anticancer Res. 2013, 33, 949–955. [Google Scholar] [PubMed]
- Todoric, J.; Strobl, B.; Jais, A.; Boucheron, N.; Bayer, M.; Amann, S.; Lindroos, J.; Teperino, R.; Prager, G.; Bilban, M. Cross-talk between interferon-γ and hedgehog signaling regulates adipogenesis. Diabetes 2011, 60, 1668–1676. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, A.; Foster, C.; Landry, A.; Sorisky, A. The role of interleukin 1β in the anti-adipogenic action of macrophages on human preadipocytes. J. Endocrinol. 2013, 217, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Dewulf, E.M.; Ge, Q.; Bindels, L.B.; Sohet, F.M.; Cani, P.D.; Brichard, S.M.; Delzenne, N.M. Evaluation of the relationship between GPR43 and adiposity in human. Nutr. Metab. 2013, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Lidell, M.E.; Betz, M.J.; Dahlqvist Leinhard, O.; Heglind, M.; Elander, L.; Slawik, M.; Mussack, T.; Nilsson, D.; Romu, T.; Nuutila, P.; et al. Evidence for two types of brown adipose tissue in humans. Nat. Med. 2013, 19, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Pfannenberg, C.; Werner, M.K.; Ripkens, S.; Stef, I.; Deckert, A.; Schmadl, M.; Reimold, M.; Häring, H.U.; Claussen, C.D.; Stefan, N. Impact of age on the relationships of brown adipose tissue with sex and adiposity in humans. Diabetes 2010, 59, 1789–1793. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, J.; Bengtsson, T.; Cannon, B. Unexpected evidence for active brown adipose tissue in adult humans. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E444–E452. [Google Scholar] [CrossRef] [PubMed]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.D.; Boström, P.; O’Sullivan, J.F.; Schinzel, R.T.; Lewis, G.D.; Dejam, A.; Lee, Y.K.; Palma, M.J.; Calhoun, S.; Georgiadi, A.; et al. β-Aminoisobutyric acid induces browning of white fat and hepatic β-oxidation and is inversely correlated with cardiometabolic risk factors. Cell Metab. 2014, 19, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Elsen, M.; Raschke, S.; Eckel, J. Browning of white fat: Does irisin play a role in humans? J. Endocrinol. 2014, 222, R25–R38. [Google Scholar] [CrossRef] [PubMed]
- Than, A.; He, H.L.; Chua, S.H.; Xu, D.; Sun, L.; Leow, M.K.; Chen, P. Apelin enhances brown adipogenesis and browning of white adipocytes. J. Biol. Chem. 2015, 290, 14679–14691. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bolze, F.; Fromme, T.; Klingenspor, M. Intrisic differences in BRITE adipogenesis of primary adipocytes from two different mouse strains. Biochim. Biophys. Acta 2014, 1841, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Hiroki, A.; Yohei, K.; Satoshi, H.; Takayuki, N.; Ken, K.; Tohru, M.; Masayuki, F. Induction of Beige-Like Adipocytes in 3T3-L1 Cells. J. Vet. Med. Sci. 2014, 76, 57–64. [Google Scholar]
- Gburcik, V.; Cawthorn, W.P.; Nedergaard, J.; Timmons, J.A.; Cannon, B. An essential role for Tbx15 in the differentiation of brown and “brite” but not white adipocytes. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E1053–E1060. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.; Chatterjee, S.; Yin, H.; Liu, R.; Lee, J.; Yechoor, V.K.; Ma, K. Novel Function of Rev-erbα in Promoting Brown Adipogenesis. Sci. Rep. 2015, 5, 11239. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.Y.; Kim, H.L.; Park, J.; Jung, Y.; Youn, D.H.; Lee, J.H.; Jin, J.S.; So, H.S.; Park, R.; Kim, S.H.; et al. Rubi Fructus (Rubus coreanus) activates the expression of thermogenic genesin vivo and in vitro. Int. J. Obes. (Lond.) 2015, 39, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Federico, L.; Ren, H.; Mueller, P.A.; Wu, T.; Liu, S.; Popovic, J.; Blalock, E.M.; Sunkara, M.; Ovaa, H.; Albers, H.M.; et al. Autotaxin and its product lysophosphatidic acid suppress brown adipose differentiation and promote diet-induced obesity in mice. Mol. Endocrinol. 2012, 26, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Werner, C.D.; Kebebew, E.; Celi, F.S. Functional thermogenic beige adipogenesis is inducible in human neck fat. Int. J. Obes. 2014, 38, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.J.; Shen, X.D.; Wen, J.B.; Kong, Y.; Chu, J.J.; Yan, G.Q.; Li, T.; Liu, D.; Wu, M.Q.; Zeng, G.H.; et al. Osteopontin-induced brown adipogenesis from white preadipocytes through a PI3K-AKT dependent signaling. Biochem. Biophys. Res. Commun. 2015, 459, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Sjölund, J.; Pelorosso, F.G.; Quigley, D.A.; Del Rosario, R.; Balmain, A. Identification of Hipk2 as an essential regulator of white fat development. Proc. Natl. Acad. Sci. USA. 2014, 111, 7373–7378. [Google Scholar] [CrossRef] [PubMed]
- Baboota, R.K.; Singh, D.P.; Sarma, S.M.; Kaur, J.; Sandhir, R.; Boparai, R.K.; Kondepudi, K.K.; Bishnoi, M. Capsaicin induces "brite" phenotype in differentiating 3T3-L1 preadipocytes. PLoS ONE 2014, 9, e103093. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Hong, J.; Liu, R.; Chen, M.; Xu, M.; Gu, W.; Zhang, Y.; Ma, Q.; Wang, F.; Shi, J. SFRP5 acts as a mature adipocyte marker but not as a regulator in adipogenesis. J. Mol. Endocrinol. 2014, 53, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Sheyn, D.; Pelled, G.; Tawackoli, W.; Su, S.; Ben-David, S.; Gazit, D.; Gazit, Z. Transient overexpression of Pparγ2 and C/ebpα in mesenchymal stem cells induces brown adipose tissue formation. Regen. Med. 2013, 8, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Klaus, S.; Choy, L.; Cham, O. Characterization of the novel brown adipocyte cell line HIB 1B. Adrenergic pathways involved in regulation of uncoupling protein gene expression. J. Cell Sci. 1994, 107, 313–319. [Google Scholar] [PubMed]
- Kim, W.K.; Oh, K.J.; Choi, H.R.; Park, A.; Han, B.S.; Chi, S.W.; Kim, S.J.; Bae, K.H.; Lee, S.C. MAP kinase phosphatase 3 inhibits brown adipocyte differentiation via regulation of Erk phosphorylation. Mol. Cell. Endocrinol. 2015, 416, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Lynes, M.D.; Dreyfuss, J.M.; Shamsi, F.; Schulz, T.J.; Zhang, H.; Huang, T.L.; Townsend, K.L.; Li, Y.; Takahashi, H.; et al. Clonal analyses and gene profiling identify genetic biomarkers of the thermogenic potential of human brown and white preadipocytes. Nat. Med. 2015, 21, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.I.; Chen, Y.C.; Chen, L.H.; Juan, C.C.; Ku, H.H.; Wang, S.T.; Chiou, S.H.; Chiou, G.Y.; Chi, C.W.; Hsu, C.C. PGC-1α mediates differentiation of mesenchymal stem cells to brown adipose cells. J. Atheroscler. Thromb. 2011, 18, 966–980. [Google Scholar] [CrossRef] [PubMed]
- Golabi, M.; Leung, A.; Lopez, C. Simpson-Golabi-Behmel Syndrome Type 1. Gene. Reviews; University of Washington: Seattle, DC, USA, 2006; pp. 1993–2016. [Google Scholar]
- Wabitsch, M.; Brenner, R.E.; Melzner, I.; Braun, M.; Möller, P.; Heinze, E.; Debatin, K.M.; Hauner, H. Characterization of a human preadipocyte cell strain with high capacity for adipose differentiation. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Ludewig, A.H.; Klapper, M.; Wabitsch, M.; Döring, F.; Nitz, I. Differential expression of alternative Acyl-CoA binding protein (ACBP) transcripts in an inducible human preadipocyte cell line. Horm. Metab. Res. 2011, 43, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Fischer-Posovszky, P.; Newell, F.S.; Wabitsch, M.; Tornqvist, H.E. Human SGBS cells—A unique tool for studies of human fat cell biology. Obes. Facts 2008, 1, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Allott, E.H.; Oliver, E.; Lysaght, J.; Gray, S.G.; Reynolds, J.V.; Roche, H.M.; Pidgeon, G.P. The SGBS cell strain as a model for the in vitro study of obesity and cancer. Clin. Transl. Oncol. 2012, 14, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Doan-Xuan, Q.M.; Sarvari, A.K.; Fischer-Posovszky, P.; Wabitsch, M.; Balajthy, Z.; Fesus, L.; Bacso, Z. High content analysis of differentiation and cell death in human adipocytes. Cytometry A 2013, 83, 933–943. [Google Scholar] [PubMed]
- Galhardo, M.; Sinkkonen, L.; Berninger, P.; Lin, J.; Sauter, T.; Heinäniemi. Integrated analysis of transcript-level regulation of metabolism revel as disease-relevant nodes of the human metabolic network. Nucleic Acids Res. 2014, 42, 1474–1496. [Google Scholar] [CrossRef] [PubMed]
- Wabitsch, M.; Brüderlein, S.; Melzner, I.; Braun, M.; Mechtersheimer, G.; Möller, P. LiSa-2, a novel human liposarcoma cell line with a high capacity for terminal adipose differentiation. Int. J. Cancer. 2000, 88, 889–894. [Google Scholar] [CrossRef]
- Van Beek, E.A.; Bakker, A.H.; Kruyt, P.M.; Vink, C.; Saris, W.H.; Franssen-van Hal, N.L.; Keijer, J. Comparative expression analysis of isolated human adipocytes and the human adipose cell lines LiSa-2 and PAZ6. Int. J. Obes. 2008, 32, 912–921. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Park, Y.; Namkoong, S.; Lee, J. Esculetin inhibits the inflammatory response by inducing heme oxigenase-1 in cocultured macrophages and adipocytes. Food Funct. 2014, 5, 2371–2377. [Google Scholar] [CrossRef] [PubMed]
- Keuper, M.; Dzyakanchuk, A.; Amrein, K.E.; Wabitsch, M.; Fischer-Posovszky, P. THP-1 Macrophages and SGBS Adipocytes—A New Human in vitro Model System of Inflamed Adipose Tissue. Front. Endocrinol. 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Chazenbalk, G.; Bertolotto, C.; Heneidi, S.; Jumabay, M.; Trivax, B.; Aronowitz, J.; Yoshimura, K.; Simmons, C.F.; Dumesic, D.A.; Azziz, R. Novel pathway of adipogenesis through cross-talk between adipose tissue macrophages, adipose stem cells and adipocytes: Evidence of cell plasticity. PLoS ONE 2011, 6, e17834. [Google Scholar] [CrossRef] [PubMed]
- Challa, T.D.; Straub, L.G.; Balaz, M.; Kiehlmann, E.; Donze, O.; Rudofsky, G.; Ukropec, J.; Ukropcova, B.; Wolfrum, C. Regulation of de novo Adipocyte Differentiation through Cross Talk between Adipocytes and Preadipocytes. Diabetes 2015, 64, 4075–4087. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.A.; Tang, Y.; Weiss, S.J.; Janorkar, A.V. Three-Dimensional Spheroid Cell Model of in vitro Adipocyte Inflammation. Tissue Eng. 2015, 21, 11–12. [Google Scholar]
- Unser, A.M.; Mooney, B.; Corr, D.T.; Tseng, Y.H.; Xie, Y. 3D brown adipogenesis to create Brown-Fat-in-Microstrands. Biomaterials. 2016, 75, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Brännmark, C.; Paul, A.; Ribeiro, D.; Magnusson, B.; Brolén, G.; Enejder, A.; Forslöw, A. Increased adipogenesis of human adipose-derived stem cells on polycaprolactone fiber matrices. PLoS ONE 2014, 9, e113620. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cell Model | Source | Differentiation Time | Characteristics | Differentiation Cocktail | Articles in the Last 5 Years |
|---|---|---|---|---|---|
| 3T3-L1 | Murine Swiss 3T3 cells from embryos | 14 days | Easier and less costly to use than freshly isolated cells Stand a high number of passages Homogenous response to treatments and experiments | Insulin, DEX, IBMX and FBS (Green and Meuth 1974 [
22]) DEX and troglitazone (Vishwanath et al. [24]) Rosiglitazone (Zebisch et al. [25]) | 392 |
| 3T3-F442A | Murine Swiss 3T3 cells | 14 days | More advanced commitment towards adipocyte differentiation than 3T3-L1 cells | Insulin, DEX, IBMX and FBS (Green and Kehinde [23]) | 7 |
| C3H10T1/2 | Mouse embryonic stem cell precursor | 12 days | Homogeneous population of multipotent cells More commitment of stem cells towards the adipocyte lineage | Demethylating agent 5-azacytidine and 10% FBS, insulin and DEX (Reznikoff et al. [26]) | 13 |
| OP9 | Mouse stromal cell | 72 h | Fast adipogenic differentiation (72 h) Confluent after many passages Long periods in culture suitable for high-throughput screening | IOM medium (Wolins et al. [21]) | 6 |
| MEFs | Mouse embryonic fibroblasts | 14–15 days | Unlimited, undifferentiated proliferation in vitro | Fei medium (Fei et al. [27]) Petrov medium and Bauters medium [28] | 9 |
| Elsen et al [138] | Than et al. [139] | Li et al. [140] | Pisani et al. [102] | Hiroki et al. [141] | Gburcik et al. [142] | |
|---|---|---|---|---|---|---|
| Cells | hASC (Subcutaneous AT) | Rat Primary Preadipocytes | Mice Primary Cultures of White and Brown Adipocytes | hMADS | 3T3-L1 | Brown Adipocytes (3129/Sv Strain of Mice) |
| Adipogenic induction | ||||||
| Insulin | 66 nM | 860 nM | 850 nM | 850 nM | 1.72 nM | 2.4 nM |
| Dexamethasone | 5.1 nM | 1 nM | 1 mM | 1 μM | 0.25 μM | |
| Indomethacin | 125 μM | 125 nM | ||||
| IBMX | 0.5 mM | 0.5 mM | 0.5 mM | 0.5 mM | 0.5 mM | |
| Rosiglitazone | 0.5 μM | 1 mM | 1 μM | 0.1 μM | 1 μM | 1 μM |
| FBS | 10% | 10% | 10% | |||
| T3 | 1 nM | 1 nM | 1 nM | 0.2 nM | 50 nM | |
| Cortisol | 100 nM | |||||
| Transferrin | 10 μg/mL | |||||
| Apo-transferrin | 10 mg/mL | |||||
| Gentamycin | 50 mg/mL | |||||
| Troglitazone | 5 μM | |||||
| Ascorbate | 25 g/mL | |||||
| l-Glutamine | 4 mM | |||||
| Differentiation Days | 14 | 8 | 7 | 14–16 | 8 | 6 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Ojeda, F.J.; Rupérez, A.I.; Gomez-Llorente, C.; Gil, A.; Aguilera, C.M. Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. Int. J. Mol. Sci. 2016, 17, 1040. https://doi.org/10.3390/ijms17071040
Ruiz-Ojeda FJ, Rupérez AI, Gomez-Llorente C, Gil A, Aguilera CM. Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. International Journal of Molecular Sciences. 2016; 17(7):1040. https://doi.org/10.3390/ijms17071040
Chicago/Turabian StyleRuiz-Ojeda, Francisco Javier, Azahara Iris Rupérez, Carolina Gomez-Llorente, Angel Gil, and Concepción María Aguilera. 2016. "Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review" International Journal of Molecular Sciences 17, no. 7: 1040. https://doi.org/10.3390/ijms17071040
APA StyleRuiz-Ojeda, F. J., Rupérez, A. I., Gomez-Llorente, C., Gil, A., & Aguilera, C. M. (2016). Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. International Journal of Molecular Sciences, 17(7), 1040. https://doi.org/10.3390/ijms17071040