
Astroglial Activation by an Enriched Environment after Transplantation of Mesenchymal Stem Cells Enhances Angiogenesis after Hypoxic-Ischemic Brain Injury

 ,
, 

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. The Combination of an Enriched Environment (EE) and Mesenchymal Stem Cells (MSCs) Induces Astrocyte Activation
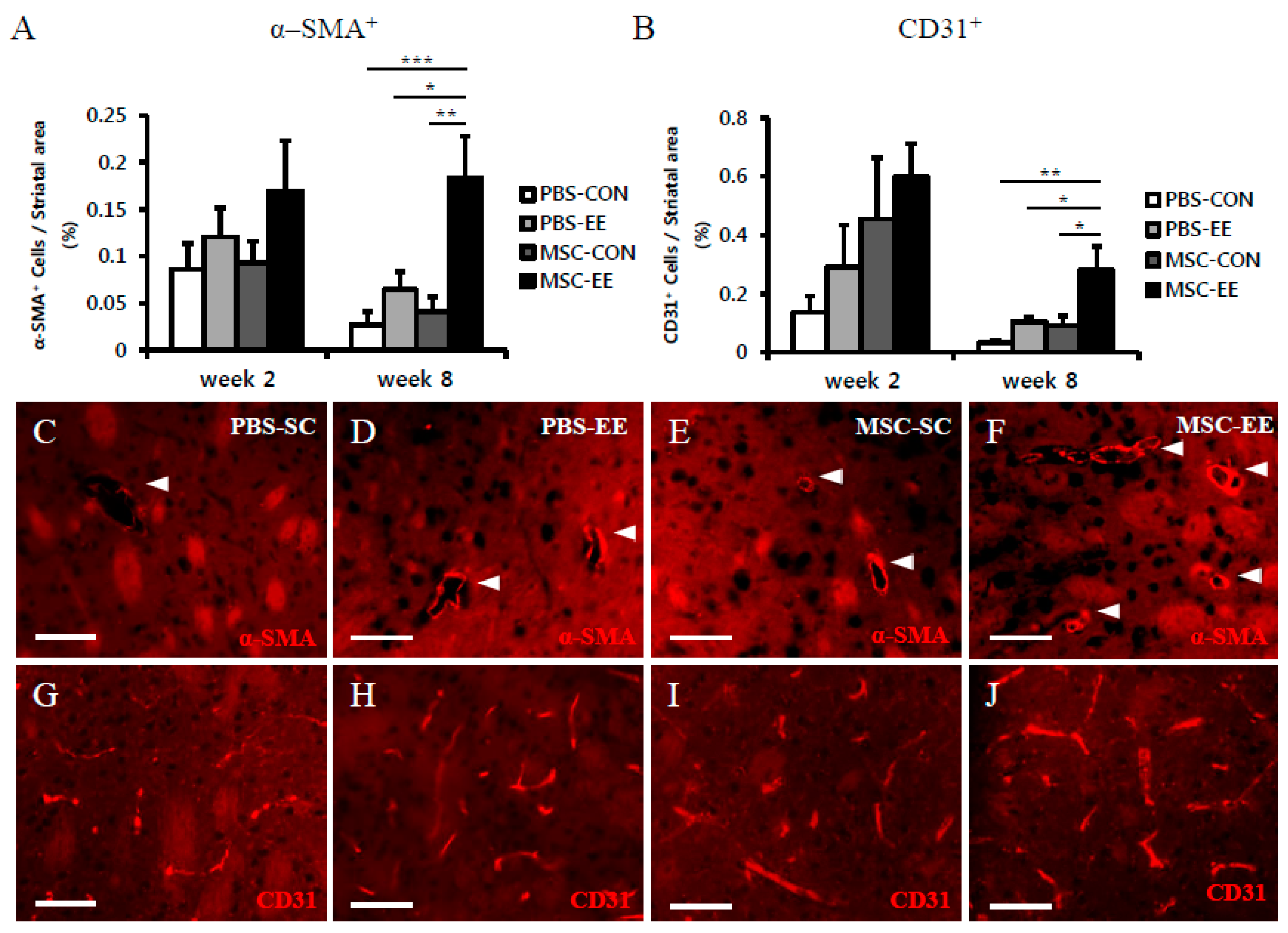
2.2. The Combination of an EE and MSCs Enhances Endogenous Angiogenesis
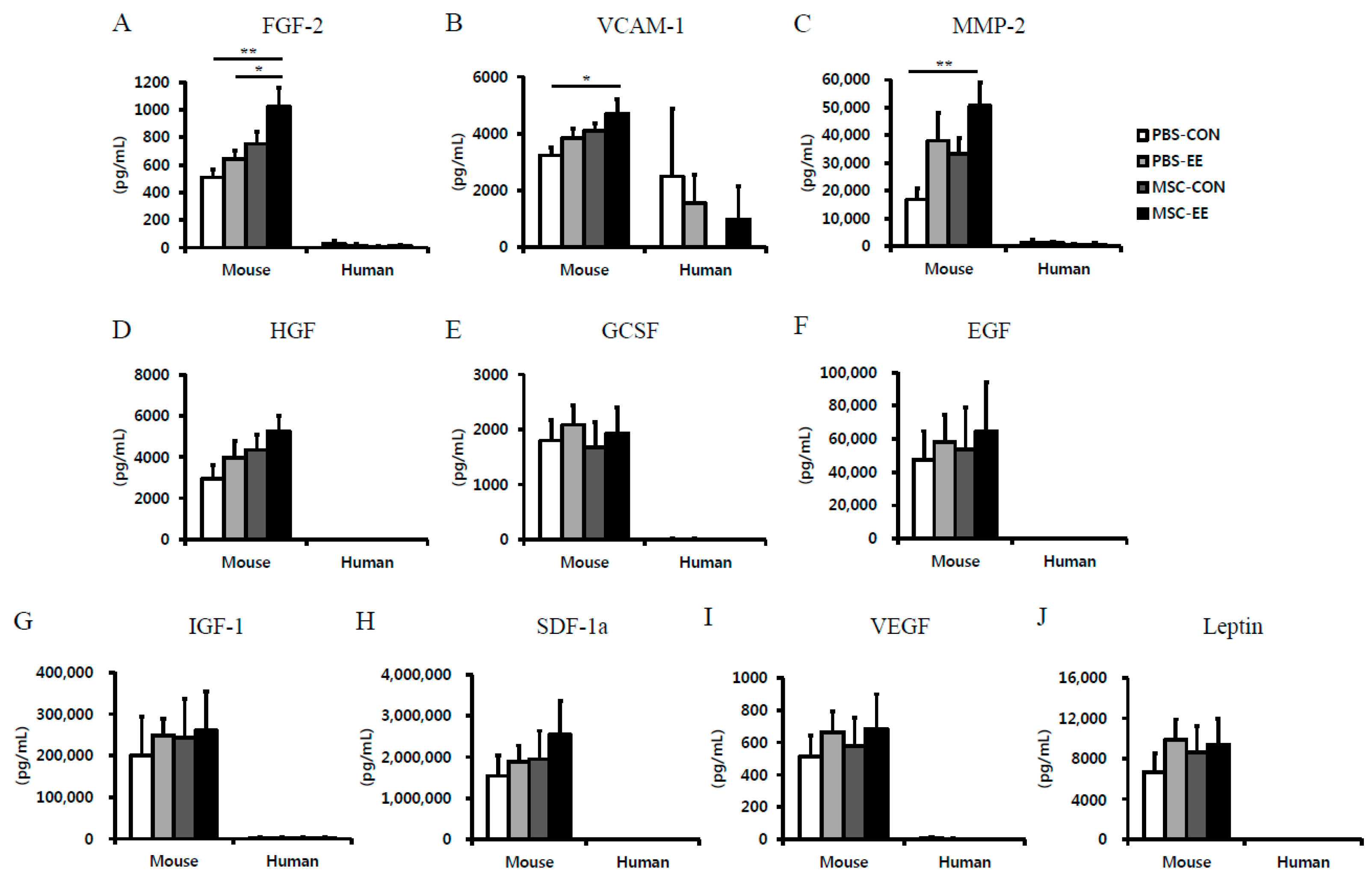
2.3. The Synergistic Effects of MSCs and an EE Upregulate Angiogenic Factors
2.4. Astrocytes Co-Cultured with MSCs Secrete Angiogenic Factors
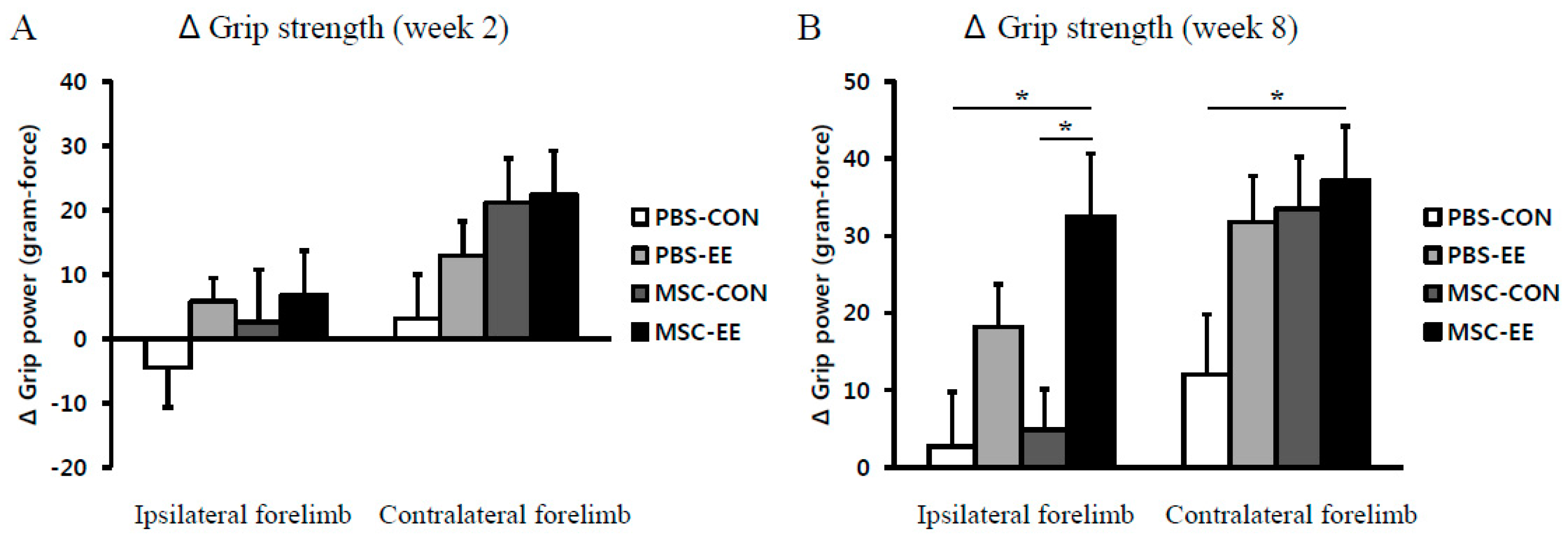
2.5. Rotarod Performance and Grip Strength Results Indicate that an EE Synergistically Improves Neurobehavioral Function after MSC Transplantation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Neonatal Hypoxic-Ischemic Brain Injury
4.3. Cell Transplantation
4.4. Grouping and Experimental Housing
4.5. Behavioral Assessment
4.6. Cell Culture: Co-Culture System
4.7. Immunohistochemistry
4.8. Assessment of Growth Factors in the Striatum
4.9. Western Blotting
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yu, J.H.; Seo, J.H.; Lee, J.Y.; Lee, M.Y.; Cho, S.R. Induction of neurorestoration from endogenous stem cells. Cell Transplant. 2016, 25, 863–882. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, M.R.; Bennett, E.L.; Hebert, M.; Morimoto, H. Social grouping cannot account for cerebral effects of enriched environments. Brain Res. 1978, 153, 563–576. [Google Scholar] [CrossRef]
- Seo, J.H.; Kim, H.; Park, E.S.; Lee, J.E.; Kim, D.W.; Kim, H.O.; Im, S.H.; Yu, J.H.; Kim, J.Y.; Lee, M.Y.; et al. Environmental enrichment synergistically improves functional recovery by transplanted adipose stem cells in chronic hypoxic-ischemic brain injury. Cell Transplant. 2013, 22, 1553–1568. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Yu, J.H.; Suh, H.; Kim, M.S.; Cho, S.R. Fibroblast growth factor-2 induced by enriched environment enhances angiogenesis and motor function in chronic hypoxic-ischemic brain injury. PLoS ONE 2013, 8, e74405. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Yu, J.H.; Kim, J.Y.; Seo, J.H.; Park, E.S.; Kim, C.H.; Kim, H.; Cho, S.R. Alteration of synaptic activity-regulating genes underlying functional improvement by long-term exposure to an enriched environment in the adult brain. Neurorehabilit. Neural Repair 2013, 27, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Schaffler, A.; Buchler, C. Concise review: Adipose tissue-derived stromal cells—Basic and clinical implications for novel cell-based therapies. Stem Cells 2007, 25, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Lv, B.; Zeng, H.; Shi, D.; Liu, Y.; Chen, F.; Li, F.; Liu, X.; Zhu, R.; Yu, L.; et al. Paracrine factors secreted by MSCs promote astrocyte survival associated with GFAP downregulation after ischemic Stroke via p38 MAPK and JNK. J. Cell. Physiol. 2015, 230, 2461–2475. [Google Scholar] [CrossRef] [PubMed]
- Sherman, L.S.; Munoz, J.; Patel, S.A.; Dave, M.A.; Paige, I.; Rameshwar, P. Moving from the laboratory bench to patients’ bedside: Considerations for effective therapy with stem cells. Clin. Transl. Sci. 2011, 4, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Dulamea, A.O. The potential use of mesenchymal stem cells in stroke therapy—From bench to bedside. J. Neurol. Sci. 2015, 352, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kurozumi, K.; Nakamura, K.; Tamiya, T.; Kawano, Y.; Kobune, M.; Hirai, S.; Uchida, H.; Sasaki, K.; Ito, Y.; Kato, K.; et al. BDNF gene-modified mesenchymal stem cells promote functional recovery and reduce infarct size in the rat middle cerebral artery occlusion model. Mol. Ther.: J. Am. Soc. Gene Ther. 2004, 9, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Ikegame, Y.; Yamashita, K.; Nakashima, S.; Nomura, Y.; Yonezawa, S.; Asano, Y.; Shinoda, J.; Hara, H.; Iwama, T. Fate of graft cells: What should be clarified for development of mesenchymal stem cell therapy for ischemic stroke? Front. Cell. Neurosci. 2014, 8, 322. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.C.; Shyu, W.C.; Chiang, M.F.; Lin, S.Z.; Chang, Y.C.; Wang, H.J.; Su, C.Y.; Li, H. Enhancement of neuroplasticity through upregulation of β1-integrin in human umbilical cord-derived stromal cell implanted stroke model. Neurobiol. Dis. 2007, 27, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.R.; Duan, W.M.; Reyes, M.; Keene, C.D.; Verfaillie, C.M.; Low, W.C. Human bone marrow stem cells exhibit neural phenotypes and ameliorate neurological deficits after grafting into the ischemic brain of rats. Exp. Neurol. 2002, 174, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Meirelles Lda, S.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.R.; Kim, Y.R.; Kang, H.S.; Yim, S.H.; Park, C.I.; Min, Y.H.; Lee, B.H.; Shin, J.C.; Lim, J.B. Functional recovery after the transplantation of neurally differentiated mesenchymal stem cells derived from bone barrow in a rat model of spinal cord injury. Cell Transplant. 2009, 18, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Hardy, S.A.; Maltman, D.J.; Przyborski, S.A. Mesenchymal stem cells as mediators of neural differentiation. Curr. Stem Cell Res. Ther. 2008, 3, 43–52. [Google Scholar] [PubMed]
- Park, H.J.; Shin, J.Y.; Kim, H.N.; Oh, S.H.; Song, S.K.; Lee, P.H. Mesenchymal stem cells stabilize the blood-brain barrier through regulation of astrocytes. Stem Cell Res. Ther. 2015, 6, 187. [Google Scholar] [CrossRef] [PubMed]
- Mani, N.; Khaibullina, A.; Krum, J.M.; Rosenstein, J.M. Astrocyte growth effects of vascular endothelial growth factor (VEGF) application to perinatal neocortical explants: Receptor mediation and signal transduction pathways. Exp. Neurol. 2005, 192, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Ridet, J.L.; Malhotra, S.K.; Privat, A.; Gage, F.H. Reactive astrocytes: Cellular and molecular cues to biological function. Trends Neurosci. 1997, 20, 570–577. [Google Scholar] [CrossRef]
- Walton, M.; Connor, B.; Lawlor, P.; Young, D.; Sirimanne, E.; Gluckman, P.; Cole, G.; Dragunow, M. Neuronal death and survival in two models of hypoxic-ischemic brain damage. Brain Res. Rev. 1999, 29, 137–168. [Google Scholar] [CrossRef]
- Shen, L.H.; Li, Y.; Chopp, M. Astrocytic endogenous glial cell derived neurotrophic factor production is enhanced by bone marrow stromal cell transplantation in the ischemic boundary zone after stroke in adult rats. Glia 2010, 58, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Z.; Xin, H.; Chopp, M. The role of astrocytes in mediating exogenous cell-based restorative therapy for stroke. Glia 2014, 62, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, Z.G.; Li, Y.; Wang, L.; Xu, Y.X.; Gautam, S.C.; Lu, M.; Zhu, Z.; Chopp, M. Intravenous administration of human bone marrow stromal cells induces angiogenesis in the ischemic boundary zone after stroke in rats. Circ. Res. 2003, 92, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Fernandez, M.; Rodriguez-Frutos, B.; Ramos-Cejudo, J.; Teresa Vallejo-Cremades, M.; Fuentes, B.; Cerdan, S.; Diez-Tejedor, E. Effects of intravenous administration of allogenic bone marrow- and adipose tissue-derived mesenchymal stem cells on functional recovery and brain repair markers in experimental ischemic stroke. Stem Cell Res. Ther. 2013, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Li, Y.; Katakowski, M.; Chen, X.; Wang, L.; Lu, D.; Lu, M.; Gautam, S.C.; Chopp, M. Intravenous bone marrow stromal cell therapy reduces apoptosis and promotes endogenous cell proliferation after stroke in female rat. J. Neurosci. Res. 2003, 73, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Alder, J.; Kramer, B.C.; Hoskin, C.; Thakker-Varia, S. Brain-derived neurotrophic factor produced by human umbilical tissue-derived cells is required for its effect on hippocampal dendritic differentiation. Dev. Neurobiol. 2012, 72, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Johnston, M.V.; Fatemi, A.; Wilson, M.A.; Northington, F. Treatment advances in neonatal neuroprotection and neurointensive care. Lancet Neurol. 2011, 10, 372–382. [Google Scholar] [CrossRef]
- Kidani, Y.; Miki, Y.; Nomimura, N.; Minakawa, S.; Tanaka, N.; Miyoshi, H.; Wakabayashi, K.; Kudo, Y. The therapeutic effect of CD133+ cells derived from human umbilical cord blood on neonatal mouse hypoxic-ischemic encephalopathy model. Life Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Shinoyama, M.; Ideguchi, M.; Kida, H.; Kajiwara, K.; Kagawa, Y.; Maeda, Y.; Nomura, S.; Suzuki, M. Cortical region-specific engraftment of embryonic stem cell-derived neural progenitor cells restores axonal sprouting to a subcortical target and achieves motor functional recovery in a mouse model of neonatal hypoxic-ischemic brain injury. Front. Cell. Neurosci. 2013, 7, 128. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, H.; Maeda, K.; Morishita, R.; Iguchi, S.; Nishikawa, T.; Takami, Y.; Kikuchi, Y.; Saito, Y.; Tamai, K.; Ogihara, T.; et al. Novel autologous cell therapy in ischemic limb disease through growth factor secretion by cultured adipose tissue-derived stromal cells. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2542–2547. [Google Scholar] [CrossRef] [PubMed]
- Sadat, S.; Gehmert, S.; Song, Y.H.; Yen, Y.; Bai, X.; Gaiser, S.; Klein, H.; Alt, E. The cardioprotective effect of mesenchymal stem cells is mediated by IGF-I and VEGF. Biochem. Biophys. Res. Commun. 2007, 363, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, J.; Zhang, C.L.; Wang, L.; Lu, D.; Katakowski, M.; Gao, Q.; Shen, L.H.; Zhang, J.; Lu, M.; et al. Gliosis and brain remodeling after treatment of stroke in rats with marrow stromal cells. Glia 2005, 49, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Jang, I.K.; Kim, H.; Yang, M.S.; Lee, J.E.; Kim, H.E.; Eom, Y.W.; Lee, D.H.; Yu, J.H.; Kim, J.Y.; et al. Early immunomodulation by intravenously transplanted mesenchymal stem cells promotes functional recovery in spinal cord injured rats. Cell Med. 2011, 2, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Delcroix, G.J.; Schiller, P.C.; Benoit, J.P.; Montero-Menei, C.N. Adult cell therapy for brain neuronal damages and the role of tissue engineering. Biomaterials 2010, 31, 2105–2120. [Google Scholar] [CrossRef] [PubMed]
- Gobbo, O.L.; O’Mara, S.M. Impact of enriched-environment housing on brain-derived neurotrophic factor and on cognitive performance after a transient global ischemia. Behav. Brain Res. 2004, 152, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Dahlqvist, P.; Zhao, L.; Johansson, I.M.; Mattsson, B.; Johansson, B.B.; Seckl, J.R.; Olsson, T. Environmental enrichment alters nerve growth factor-induced gene A and glucocorticoid receptor messenger RNA expression after middle cerebral artery occlusion in rats. Neuroscience 1999, 93, 527–535. [Google Scholar] [CrossRef]
- Young, D.; Lawlor, P.A.; Leone, P.; Dragunow, M.; During, M.J. Environmental enrichment inhibits spontaneous apoptosis, prevents seizures and is neuroprotective. Nat. Med. 1999, 5, 448–453. [Google Scholar] [PubMed]
- Johansson, B.B.; Belichenko, P.V. Neuronal plasticity and dendritic spines: Effect of environmental enrichment on intact and postischemic rat brain. J. Cereb. Blood Flow Metab. 2002, 22, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Maegele, M.; Lippert-Gruener, M.; Ester-Bode, T.; Garbe, J.; Bouillon, B.; Neugebauer, E.; Klug, N.; Lefering, R.; Neiss, W.F.; Angelov, D.N. Multimodal early onset stimulation combined with enriched environment is associated with reduced CNS lesion volume and enhanced reversal of neuromotor dysfunction after traumatic brain injury in rats. Eur. J. Neurosci. 2005, 21, 2406–2418. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.M.; Winblad, B.; Granholm, A.C.; Mohammed, A.H. Environmental influences on brain neurotrophins in rats. Pharmacol. Biochem. Behav. 2002, 73, 167–175. [Google Scholar] [CrossRef]
- Cabezas, R.; Avila, M.; Gonzalez, J.; El-Bacha, R.S.; Baez, E.; Garcia-Segura, L.M.; Jurado Coronel, J.C.; Capani, F.; Cardona-Gomez, G.P.; Barreto, G.E. Astrocytic modulation of blood brain barrier: Perspectives on Parkinson’s disease. Front. Cell. Neurosci. 2014, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Kimelberg, H.K.; Nedergaard, M. Functions of astrocytes and their potential as therapeutic targets. Neurotherapeutics 2010, 7, 338–353. [Google Scholar] [CrossRef] [PubMed]
- Pavlichenko, N.; Sokolova, I.; Vijde, S.; Shvedova, E.; Alexandrov, G.; Krouglyakov, P.; Fedotova, O.; Gilerovich, E.G.; Polyntsev, D.G.; Otellin, V.A. Mesenchymal stem cells transplantation could be beneficial for treatment of experimental ischemic stroke in rats. Brain Res. 2008, 1233, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Katakowski, M.; Chen, X.; Li, Y.; Chopp, M. Human marrow stromal cells enhance connexin43 gap junction intercellular communication in cultured astrocytes. Cell Transplant. 2005, 14, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Li, Y.; Chopp, M. Bone marrow stromal cells increase astrocyte survival via upregulation of phosphoinositide 3-kinase/threonine protein kinase and mitogen-activated protein kinase kinase/extracellular signal-regulated kinase pathways and stimulate astrocyte trophic factor gene expression after anaerobic insult. Neuroscience 2005, 136, 123–134. [Google Scholar] [PubMed]
- Im, S.H.; Yu, J.H.; Park, E.S.; Lee, J.E.; Kim, H.O.; Park, K.I.; Kim, G.W.; Park, C.I.; Cho, S.R. Induction of striatal neurogenesis enhances functional recovery in an adult animal model of neonatal hypoxic-ischemic brain injury. Neuroscience 2010, 169, 259–268. [Google Scholar] [CrossRef] [PubMed]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, S.-R.; Suh, H.; Yu, J.H.; Kim, H.; Seo, J.H.; Seo, C.H. Astroglial Activation by an Enriched Environment after Transplantation of Mesenchymal Stem Cells Enhances Angiogenesis after Hypoxic-Ischemic Brain Injury. Int. J. Mol. Sci. 2016, 17, 1550. https://doi.org/10.3390/ijms17091550
Cho S-R, Suh H, Yu JH, Kim H, Seo JH, Seo CH. Astroglial Activation by an Enriched Environment after Transplantation of Mesenchymal Stem Cells Enhances Angiogenesis after Hypoxic-Ischemic Brain Injury. International Journal of Molecular Sciences. 2016; 17(9):1550. https://doi.org/10.3390/ijms17091550
Chicago/Turabian StyleCho, Sung-Rae, Hwal Suh, Ji Hea Yu, Hyongbum (Henry) Kim, Jung Hwa Seo, and Cheong Hoon Seo. 2016. "Astroglial Activation by an Enriched Environment after Transplantation of Mesenchymal Stem Cells Enhances Angiogenesis after Hypoxic-Ischemic Brain Injury" International Journal of Molecular Sciences 17, no. 9: 1550. https://doi.org/10.3390/ijms17091550
APA StyleCho, S.-R., Suh, H., Yu, J. H., Kim, H., Seo, J. H., & Seo, C. H. (2016). Astroglial Activation by an Enriched Environment after Transplantation of Mesenchymal Stem Cells Enhances Angiogenesis after Hypoxic-Ischemic Brain Injury. International Journal of Molecular Sciences, 17(9), 1550. https://doi.org/10.3390/ijms17091550