
Multiplex Gene Expression Profiling of 16 Target Genes in Neoplastic and Non-Neoplastic Canine Mammary Tissues Using Branched-DNA Assay

Abstract
:1. Introduction
2. Results
2.1. Gene Expression
2.1.1. Fresh Frozen Tissues
2.1.2. Formalin-Fixed, Paraffin-Embedded Tissues
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.1.1. Formalin-Fixed, Paraffin-Embedded Blocks
4.1.2. Fresh Frozen Tumor Samples
4.2. Nucleic Acid Isolation and Quantification
4.2.1. Formalin-Fixed, Paraffin-Embedded Samples
4.2.2. Fresh Frozen Samples
4.2.3. Target Genes
4.2.4. Housekeeping Genes
4.2.5. Multiplexed Branched-DNA (b-DNA) Assay
4.3. Histologic Classification
4.4. Data and Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bronden, L.B.; Nielsen, S.S.; Toft, N.; Kristensen, A.T. Data from the danish veterinary cancer registry on the occurrence and distribution of neoplasms in dogs in denmark. Vet. Rec. 2010, 166, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Bostock, D.E. Canine and feline mammary neoplasms. Br. Vet. J. 1986, 142, 506–515. [Google Scholar] [CrossRef]
- Dobson, J.M. Breed-predispositions to cancer in pedigree dogs. ISRN Vet. Sci. 2013. [Google Scholar] [CrossRef] [PubMed]
- Von Bomhard, D. Epidemiologie. In Praxis der Onkologie bei Hund und Katze, 1. Auflage; Nolte, I., Nolte, M., Eds.; Enke Verlag: Stuttgart, Deutschland, 2001; pp. 104–108. [Google Scholar]
- Betz, D.; Schoenrock, D.; Mischke, R.; Baumgartner, W.; Nolte, I. Postoperative treatment outcome in canine mammary tumors: Multivariate analysis of the prognostic value of pre- and postoperatively available information. Tierärztliche Praxis Ausgabe K Kleintiere Heimtiere 2012, 40, 235–242. [Google Scholar]
- Karayannopoulou, M.; Kaldrymidou, E.; Constantinidis, T.C.; Dessiris, A. Histological grading and prognosis in dogs with mammary carcinomas: Application of a human grading method. J. Comp. Pathol. 2005, 133, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Duraiyan, J.; Govindarajan, R.; Kaliyappan, K.; Palanisamy, M. Applications of immunohistochemistry. J. Pharm. Bioallied Sci. 2012, 4, S307–309. [Google Scholar] [PubMed]
- Tavasoly, A.; Golshahi, H.; Rezaie, A.; Farhadi, M. Classification and grading of canine malignant mammary tumors. Vet. Res. Forum: Int. Q. J. 2013, 4, 25–30. [Google Scholar]
- Cassali, G.D.; Lavalle, G.E.; Ferreira, E.; Estrela-Lima, A.; de Nardi, A.B.; Bertagnolli, A.C.; Alessi, A.C.; Daleck, C.R.; Salgado, B.S.; Fernandes, C.G.; et al. Consensus for the diagnosis, prognosis and treatment of canine mammary tumors. Braz. J. Vet. Pathol. 2011, 4, 153–180. [Google Scholar]
- Seidal, T.; Balaton, A.J.; Battifora, H. Interpretation and quantification of immunostains. Am. J. Sur. Pathol. 2001, 25, 1204–1207. [Google Scholar] [CrossRef]
- Flagella, M.; Bui, S.; Zheng, Z.; Nguyen, C.T.; Zhang, A.; Pastor, L.; Ma, Y.; Yang, W.; Crawford, K.L.; McMaster, G.K.; et al. A multiplex branched DNA assay for parallel quantitative gene expression profiling. Anal. Biochem. 2006, 352, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, B.S.; Allen, A.N.; McLerran, D.F.; Vessella, R.L.; Karademos, J.; Davies, J.E.; Maqsodi, B.; McMaster, G.K.; Kristal, A.R. Evaluation of the branched-chain DNA assay for measurement of rna in formalin-fixed tissues. J. Mol. Diagn. 2008, 10, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Pabinger, S.; Rödigerb, S.; Kriegnera, A.; Vierlingera, K.; Weinhäusel, A. A survey of tools for the analysis of quantitative PCR (qPCR) data. Elsevier 2014, 1, 23–33. [Google Scholar] [CrossRef]
- Queiroga, F.L.; Raposo, T.; Carvalho, M.I.; Prada, J.; Pires, I. Canine mammary tumours as a model to study human breast cancer: Most recent findings. In Vivo 2011, 25, 455–465. [Google Scholar] [PubMed]
- Hennecke, S.; Beck, J.; Bornemann-Kolatzki, K.; Neumann, S.; Murua Escobar, H.; Nolte, I.; Hammer, S.C.; Hewicker-Trautwein, M.; Junginger, J.; Kaup, F.J.; et al. Prevalence of the prefoldin subunit 5 gene deletion in canine mammary tumors. PLoS ONE 2015, 10, e0131280. [Google Scholar] [CrossRef] [PubMed]
- Rivera, P.; Melin, M.; Biagi, T.; Fall, T.; Haggstrom, J.; Lindblad-Toh, K.; von Euler, H. Mammary tumor development in dogs is associated with BRCA1 and BRCA2. Cancer Res. 2009, 69, 8770–8774. [Google Scholar] [CrossRef] [PubMed]
- Klopfleisch, R.; Gruber, A.D. Increased expression of BRCA2 and RAD51 in lymph node metastases of canine mammary adenocarcinomas. Vet. Pathol. 2009, 46, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Nieto, A.; Perez-Alenza, M.D.; del Castillo, N.; Tabanera, E.; Castano, M.; Pena, L. BRCA1 expression in canine mammary dysplasias and tumours: Relationship with prognostic variables. J. Comp. Pathol. 2003, 128, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, K.; Morimatsu, M.; Tomizawa, N.; Syuto, B. Cloning and sequencing full length of canine BRCA2 and RAD51 cDNA. J. Vet. Med. Sci. 2001, 63, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Enginler, S.O.; Akis, I.; Toydemir, T.S.; Oztabak, K.; Haktanir, D.; Gunduz, M.C.; Kirsan, I.; Firat, I. Genetic variations of BRCA1 and BRCA2 genes in dogs with mammary tumours. Vet. Res. Commun. 2014, 38, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Muto, T.; Wakui, S.; Takahashi, H.; Maekawa, S.; Masaoka, T.; Ushigome, S.; Furusato, M. P53 gene mutations occurring in spontaneous benign and malignant mammary tumors of the dog. Vet. Pathol. 2000, 37, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.L.; Rutteman, G.R.; Kong, J.M.; Ghahremani, M.; Schmeing, M.; Misdorp, W.; van Garderen, E.; Pelletier, J. Genomic organization of the canine p53 gene and its mutational status in canine mammary neoplasia. Breast Cancer Res. Treat. 1998, 50, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Kanae, Y.; Endoh, D.; Yokota, H.; Taniyama, H.; Hayashi, M. Expression of the pten tumor suppressor gene in malignant mammary gland tumors of dogs. Am. J. Vet. Res. 2006, 67, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Lin, D.; Wang, J.; Wang, L. Expression and significance of pten in canine mammary gland tumours. Res. Vet. Sci. 2008, 85, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Ressel, L.; Millanta, F.; Caleri, E.; Innocenti, V.M.; Poli, A. Reduced pten protein expression and its prognostic implications in canine and feline mammary tumors. Vet. Pathol. 2009, 46, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.; Hennecke, S.; Bornemann-Kolatzki, K.; Urnovitz, H.B.; Neumann, S.; Strobel, P.; Kaup, F.J.; Brenig, B.; Schutz, E. Genome aberrations in canine mammary carcinomas and their detection in cell-free plasma DNA. PLoS ONE 2013, 8, e75485. [Google Scholar] [CrossRef] [PubMed]
- Borge, K.S.; Nord, S.; van Loo, P.; Lingjaerde, O.C.; Gunnes, G.; Alnaes, G.I.; Solvang, H.K.; Luders, T.; Kristensen, V.N.; Borresen-Dale, A.L.; et al. Canine mammary tumours are affected by frequent copy number aberrations, including amplification of myc and loss of pten. PLoS ONE 2015, 10, e0126371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, W.L.; Huang, H.M.; Liao, J.W.; Wong, M.L.; Chang, S.C. Increased survival in dogs with malignant mammary tumours overexpressing HER-2 protein and detection of a silent single nucleotide polymorphism in the canine HER-2 gene. Vet. J. 2009, 180, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Kano, R.; Yano, T.; Nagamatsu, K.; Maruyama, H.; Kamata, H.; Hasegawa, A. Effectiveness of small interfering RNA (siRNA) against the Mcl-1 gene in a canine mammary gland tumor cell line. Res. Vet. Sci. 2009, 87, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Ripoli, F.L.; Mohr, A.; Hammer, S.C.; Willenbrock, S.; Hewicker-Trautwein, M.; Hennecke, S.; Escobar, H.M.; Nolte, I. A comparison of fresh frozen vs. formalin-fixed, paraffin-embedded specimens of canine mammary tumors via branched-DNA assay. Int. J. Mol. Sci. 2016. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, T.; Kong, S.Y.; Kwon, S.; Bae, C.Y.; Choi, J.; Kim, C.H.; Lee, E.S.; Park, J.K. Breast cancer diagnosis using a microfluidic multiplexed immunohistochemistry platform. PLoS ONE 2010, 5, e10441. [Google Scholar] [CrossRef] [PubMed]
- Rivera, P.; von Euler, H. Molecular biological aspects on canine and human mammary tumors. Vet. Pathol. 2011, 48, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Mavaddat, N.; Antoniou, A.C.; Easton, D.F.; Garcia-Closas, M. Genetic susceptibility to breast cancer. Mol. Oncol. 2010, 4, 174–191. [Google Scholar] [CrossRef] [PubMed]
- Gadzicki, D.; Evans, D.G.; Harris, H.; Julian-Reynier, C.; Nippert, I.; Schmidtke, J.; Tibben, A.; van Asperen, C.J.; Schlegelberger, B. Genetic testing for familial/hereditary breast cancer—comparison of guidelines and recommendations from the UK, France, the Netherlands and Germany. J. Commun. Genet. 2011, 2, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Accili, D.; Arden, K.C. Foxos at the crossroads of cellular metabolism, differentiation, and transformation. Cell 2004, 117, 421–426. [Google Scholar] [CrossRef]
- Gasco, M.; Shami, S.; Crook, T. The p53 pathway in breast cancer. Breast Cancer Res. 2002, 4, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yen, C.; Liaw, D.; Podsypanina, K.; Bose, S.; Wang, S.I.; Puc, J.; Miliaresis, C.; Rodgers, L.; McCombie, R.; et al. Pten, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science 1997, 275, 1943–1947. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, Y.; Taira, T.; Maeda, Y.; Tanaka, S.; Nishihara, H.; Iguchi-Ariga, S.M.; Nagashima, K.; Ariga, H. Mm-1, a c-Myc-binding protein, is a candidate for a tumor suppressor in leukemia/lymphoma and tongue cancer. J. Biol. Chem. 2001, 276, 45137–45144. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zou, L.; Lu, W.Q.; Zhang, Y.; Shen, A.G. Foxo3a expression is a prognostic marker in breast cancer. PLoS ONE 2013, 8, e70746. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, I.S.; Hellmen, E.; Cornelisse, C.J.; van den Burgh, B.; Rutteman, G.R. P53 mutations in mammary tumor cell lines and corresponding tumor tissues in the dog. Anticancer Res. 1996, 16, 3737–3744. [Google Scholar] [PubMed]
- Yang, P.; Du, C.W.; Kwan, M.; Liang, S.X.; Zhang, G.J. The impact of p53 in predicting clinical outcome of breast cancer patients with visceral metastasis. Sci. Rep. 2013. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Wang, J. A physical mechanism and global quantification of breast cancer. PLoS ONE 2016, 11, e0157422. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Crane, A.; Hibshoosh, H.; Mansukhani, M.; Sandweis, L.; Parsons, R. Reduced expression of PTEN correlates with breast cancer progression. Hum. Pathol. 2002, 33, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lindblom, P.; Lindblom, A. A study of the PTEN/MMAC1 gene in 136 breast cancer families. Hum. Genet. 1998, 102, 124–125. [Google Scholar] [PubMed]
- Li, X.; Wang, Q.; Fu, L.; Liu, M.; Yu, X. Expression of PTEN, p53 and EGFR in the molecular subtypes of breast carcinoma and the correlation among them. Med. Sci. 2015, 40, 973–978. [Google Scholar]
- Sivaraman, V.S.; Wang, H.; Nuovo, G.J.; Malbon, C.C. Hyperexpression of mitogen-activated protein kinase in human breast cancer. J. Clin. Investig. 1997, 99, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zhang, W.; Cui, Z.; Chen, Q.; Xie, P.; Zhou, C.; Liu, B.; Peng, X.; Zhang, Y. High mobility group box-1 and its clinical value in breast cancer. Onco Targets Ther. 2015, 8, 413–419. [Google Scholar] [PubMed]
- Fusco, A.; Fedele, M. Roles of HMGA proteins in cancer. Nat. Rev. Cancer 2007, 7, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Levine, D.A.; Bogomolniy, F.; Yee, C.J.; Lash, A.; Barakat, R.R.; Borgen, P.I.; Boyd, J. Frequent mutation of the PIK3CA gene in ovarian and breast cancers. Clin. Cancer Res. 2005, 11, 2875–2878. [Google Scholar] [CrossRef] [PubMed]
- O’Driscoll, L.; Cronin, D.; Kennedy, S.M.; Purcell, R.; Linehan, R.; Glynn, S.; Larkin, A.; Scanlon, K.; McDermott, E.W.; Hill, A.D.; et al. Expression and prognostic relevance of Mcl-1 in breast cancer. Anticancer Res. 2004, 24, 473–482. [Google Scholar] [PubMed]
- Ross, J.S.; Fletcher, J.A.; Linette, G.P.; Stec, J.; Clark, E.; Ayers, M.; Symmans, W.F.; Pusztai, L.; Bloom, K.J. The HER-2/NEU gene and protein in breast cancer 2003: Biomarker and target of therapy. Oncologist 2003, 8, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H. Gene and chromosomal alterations in sporadic breast cancer: Correlation with histopathological features and implications for genesis and progression. Breast Cancer 2009, 16, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Sana, M.; Irshad, S. A review on breast cancer biomarkers BRCA1 and BRCA2. Res. Cancer Tumor 2012, 1, 1–4. [Google Scholar]



{kind=link}
{kind=link}
| Group | FF (n) | FFPE (n) |
|---|---|---|
| 1. Healthy mammary tissue (non-neoplastic tissue) | 15 | 4 |
| 2. Lobular hyperplasias (non-neoplastic tissue) | 3 | 20 |
| 3. Benign tumors | 33 | 47 |
| 4. Malignant tumors | 60 | 84 |
| Total | 111 | 155 |
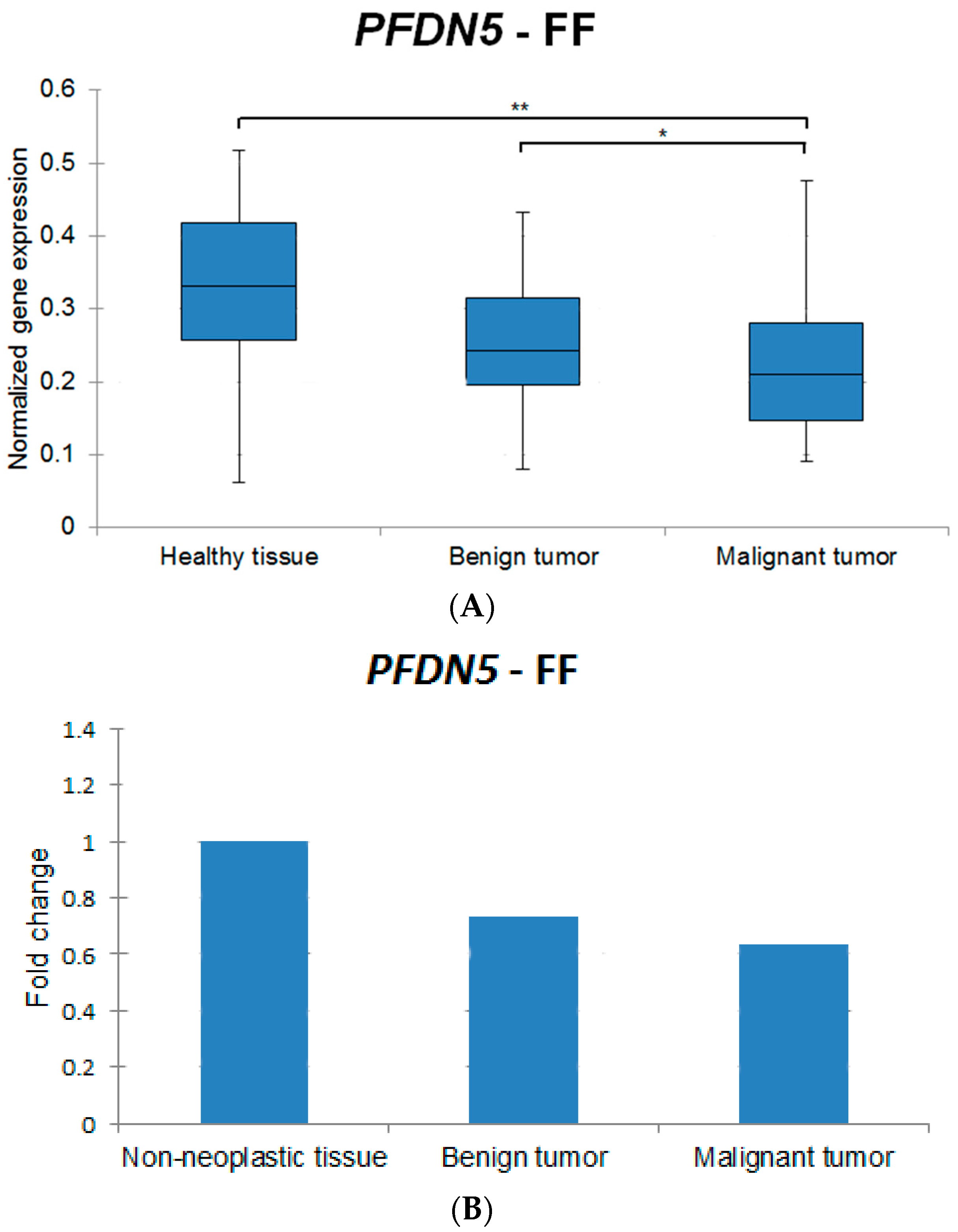
| Target Gene | Healthy Tissue vs. Benign Tumors | Healthy Tissue vs. Malignant Tumors | Benign Tumors vs. Malignant Tumors |
|---|---|---|---|
| BRCA1 | - | - | - |
| BRCA2 | - | - | - |
| FOXO3 | - | ↑ * | ↑ * |
| GATA4 | - | - | - |
| HER2 | - | ↑ * | ↑ * |
| HMGA1 | ↑ ** | ↑ * | - |
| HMGA2 | - | - | |
| HMGB1 | ↑ * | - | - |
| MAPK1 | - | - | - |
| MAPK3 | - | ↑ * | - |
| MCL1 | ↑ * | ↑ ** | ↑ * |
| MYC | - | - | - |
| PFDN5 | - | ↑ ** | ↑ * |
| PIK3CA | - | - | - |
| PTEN | - | - | - |
| TP53 | - | ↑ * | - |
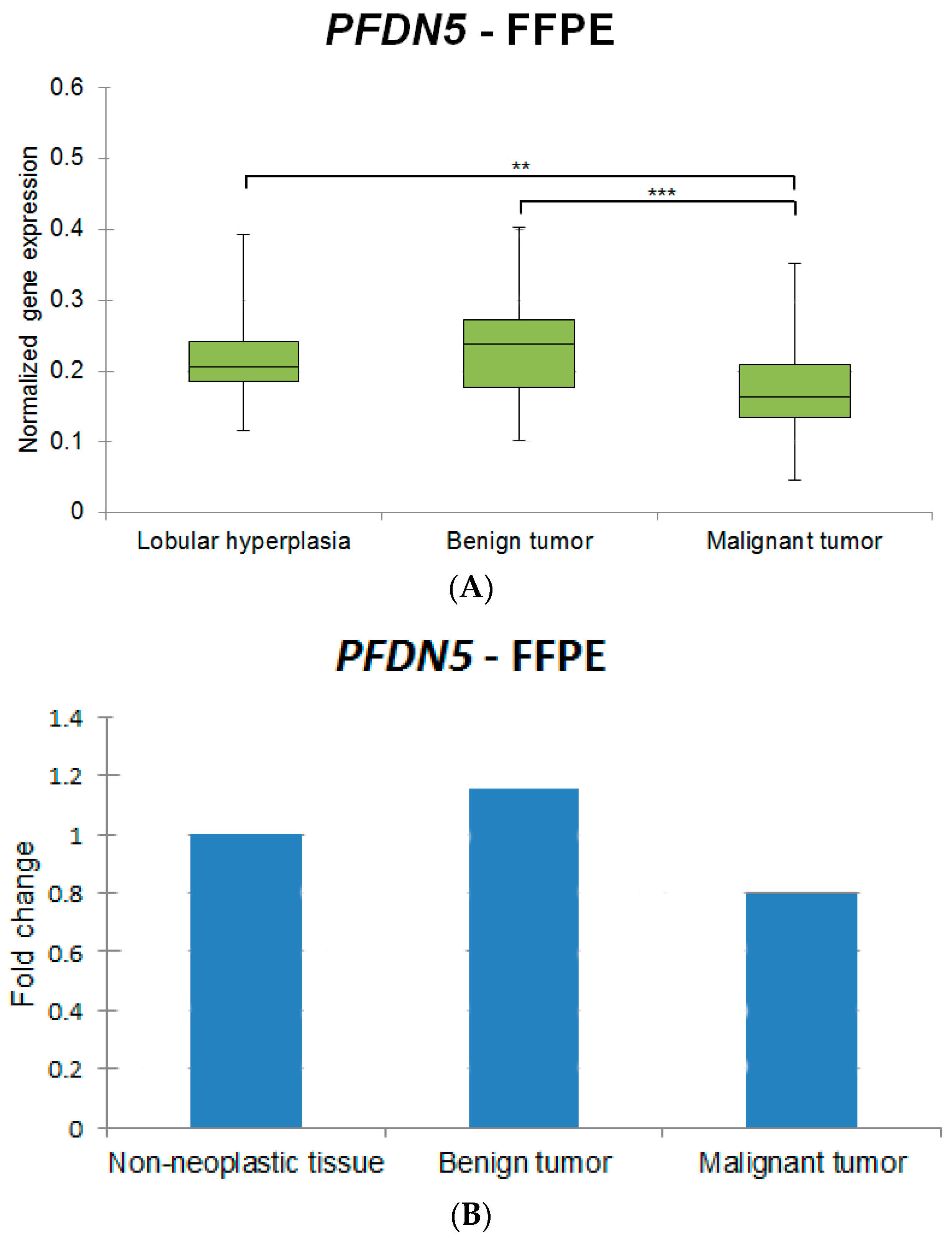
| Target Gene | Lobular Hyperplasias vs. Malignant Tumors | Benign Tumors vs. Malignant Tumors |
|---|---|---|
| BRCA1 | - | - |
| BRCA2 | - | - |
| FOXO3 | ↑ ** | ↑ *** |
| GATA4 | - | - |
| HER2 | ↑ *** | ↑ *** |
| HMGA1 | ↑ * | - |
| HMGA2 | - | - |
| HMGB1 | ↑ *** | ↑ * |
| MAPK1 | - | - |
| MAPK3 | ↑ ** | ↑ *** |
| MCL1 | ↑ *** | ↑ *** |
| MYC | - | - |
| PFDN5 | ↑ ** | ↑ *** |
| PIK3CA | ↑ * | ↑ ** |
| PTEN | - | ↑ ** |
| TP53 | ↑ * | ↑ * |
| Target Gene | Approved Name | Accession Number |
|---|---|---|
| BRCA1 | BRCA1, DNA repair associated | NM_001013416 |
| BRCA2 | BRCA2, DNA repair associated | NM_001006653 |
| FOXO3 | Forkhead box O3 | NM_003639400 |
| GATA4 | GATA binding protein 4 | NM_001048112 |
| HER2 | Erb-b2 receptor tyrosine kinase 2 | NM_001003217 |
| HMGA1 | High mobility group AT-hook 1 | NM_001003387 |
| HMGA2 | High mobility group AT-hook 2 | XM_005625590 |
| HMGB1 | High mobility group box 1 | NM_001002937 |
| MAPK1 | Mitogen-activated protein kinase 1 | NM_001110800 |
| MAPK3 | Mitogen-activated protein kinase 3 | NM_001252035 |
| MCL1 | Myeloid cell leukemia 1 | NM_001003016 |
| MYC | V-myc avian myelocytomatosis viral oncogene homolog | NM_001003246 |
| PFDN5 | Prefoldin subunit 5 | NM_001251949 |
| PIK3CA | Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit α | XM_545208.4 |
| PTEN | Phosphatase and tensin homolog | NM_001003192 |
| TP53 | Tumor protein p53 | NM_001003210 |
| ACTB | β-actin | XM_536888 |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase | NM_001003142 |
| HPRT1 | Hypoxanthine phosphoribosyltransferase 1 | NM_001003357 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lüder Ripoli, F.; Conradine Hammer, S.; Mohr, A.; Willenbrock, S.; Hewicker-Trautwein, M.; Brenig, B.; Murua Escobar, H.; Nolte, I. Multiplex Gene Expression Profiling of 16 Target Genes in Neoplastic and Non-Neoplastic Canine Mammary Tissues Using Branched-DNA Assay. Int. J. Mol. Sci. 2016, 17, 1589. https://doi.org/10.3390/ijms17091589
Lüder Ripoli F, Conradine Hammer S, Mohr A, Willenbrock S, Hewicker-Trautwein M, Brenig B, Murua Escobar H, Nolte I. Multiplex Gene Expression Profiling of 16 Target Genes in Neoplastic and Non-Neoplastic Canine Mammary Tissues Using Branched-DNA Assay. International Journal of Molecular Sciences. 2016; 17(9):1589. https://doi.org/10.3390/ijms17091589
Chicago/Turabian StyleLüder Ripoli, Florenza, Susanne Conradine Hammer, Annika Mohr, Saskia Willenbrock, Marion Hewicker-Trautwein, Bertram Brenig, Hugo Murua Escobar, and Ingo Nolte. 2016. "Multiplex Gene Expression Profiling of 16 Target Genes in Neoplastic and Non-Neoplastic Canine Mammary Tissues Using Branched-DNA Assay" International Journal of Molecular Sciences 17, no. 9: 1589. https://doi.org/10.3390/ijms17091589
APA StyleLüder Ripoli, F., Conradine Hammer, S., Mohr, A., Willenbrock, S., Hewicker-Trautwein, M., Brenig, B., Murua Escobar, H., & Nolte, I. (2016). Multiplex Gene Expression Profiling of 16 Target Genes in Neoplastic and Non-Neoplastic Canine Mammary Tissues Using Branched-DNA Assay. International Journal of Molecular Sciences, 17(9), 1589. https://doi.org/10.3390/ijms17091589