Explaining the Microtubule Energy Balance: Contributions Due to Dipole Moments, Charges, van der Waals and Solvation Energy


 , , and
, , and 
Abstract
:Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Desai, A.; Mitchison, T.J. Microtubule Polymerization Dynamics. Ann. Rev. Cell Dev. Biol. 1997, 13, 83–117. [Google Scholar] [CrossRef] [PubMed]
- Sept, D.; Baker, N.A.; McCammon, J.A. The Physical Basis of Microtubule Structure and Stability. Protein Sci. 2003, 12, 2257–2261. [Google Scholar] [CrossRef] [PubMed]
- Drabik, P.; Gusarov, S.; Kovalenko, A. Microtubule Stability Studied by Three-Dimensional Molecular Theory of Solvation. Biophys. J. 2007, 92, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Erickson, H.P.; Pantaloni, D. The Role of Subunit Entropy in Cooperative Assembly. Nucleation of Microtubules and Other Two-Dimensional Polymers. Biophys. J. 1981, 34, 293–309. [Google Scholar] [CrossRef]
- Ayoub, A.T.; Craddock, T.; Klobukowski, M.; Tuszynski, J. Analysis of the Strength of Interfacial Hydrogen Bonds between Tubulin Dimers Using Quantum Theory of Atoms in Molecules. Biophys. J. 2014, 107, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, A.T.; Klobukowski, M.; Tuszynski, J.A. Detailed Per-residue Energetic Analysis Explains the Driving Force for Microtubule Disassembly. PLoS Comput. Biol. 2015, 11, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Kollman, P.A.; Massova, I.; Reyes, C.; Kuhn, B.; Huo, S.; Chong, L.; Lee, M.; Lee, T.; Duan, Y.; Wang, W.; et al. Calculating Structures and Free Energies of Complex Molecules: Combining Molecular Mechanics and Continuum Models. Acc. Chem. Res. 2000, 33, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Tuszynski, J.A.; Carpenter, E.J.; Huzil, J.T.; Malinski, W.; Luchko, T.; Ludueña, R.F. The Evolution of the Structure of Tubulin and its Potential Consequences for the Role and Function of Microtubules in Cells and Embryos. Int. J. Dev. Biol. 2006, 50, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, E.J.; Huzil, J.T.; Ludueña, R.F.; Tuszynski, J.A. Homology Modeling of Tubulin: Influence Predictions for Microtubule’s Biophysical Properties. Eur. Biophys. J. 2006, 36, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Katsuki, M.; Drummond, D.R.; Cross, R.A. Ectopic A-lattice Seams Destabilize Microtubules. Nat. Commun. 2014. [Google Scholar] [CrossRef] [PubMed]
- Beard, D.A.; Qian, H. Chemical Biophysics: Quantitative Analysis of Cellular Systems; Cambridge University Press: Cambridge, UK, 2008; p. 119. [Google Scholar]
- Ludueña, R.F. Multiple Forms of Tubulin: Different Gene Products and Covalent Modifications. Int. Rev. Cytol. 1997, 178, 207–275. [Google Scholar]
- Panda, D.; Miller, H.P.; Banerjee, A.; Ludueña, R.F.; Wilson, L. Microtubule Dynamics in vitro are Regulated by the Tubulin Isotype Composition. Proc. Natl. Acad. Sci. USA 1994, 91, 11358–11362. [Google Scholar] [CrossRef] [PubMed]
- Seetapun, D.; Castle, B.; McIntyre, A.; Tran, P.; Odde, D. Estimating the Microtubule GTP Cap Size in Vivo. Curr. Biol. 2012, 22, 1681–1687. [Google Scholar] [CrossRef] [PubMed]
- Duellberg, C.; Cade, N.I.; Holmes, D.; Surrey, T. The Size of the EB Cap Determines Instantaneous Microtubule Stability. Elife 2016. [Google Scholar] [CrossRef] [PubMed]
- Coombes, C.; Yamamoto, A.; Kenzie, M.; Odde, D.; Gardner, M. Evolving Tip Structures Can Explain Age-Dependent Microtubule Catastrophe. Curr. Biol. 2013, 23, 1342–1348. [Google Scholar] [CrossRef] [PubMed]
- Duellberg, C.; Cade, N.I.; Surrey, T. Microtubule Aging Probed by Microfluidics-assisted Tubulin Washout. Mol. Biol. Cell 2016, 27, 3563–3573. [Google Scholar] [CrossRef] [PubMed]
- Margolin, G.; Gregoretti, I.V.; Cickovski, T.M.; Li, C.; Shi, W.; Alber, M.S.; Goodson, H.V. The Mechanisms of Microtubule Catastrophe and Rescue: Implications from Analysis of a Dimer-scale Computational Model. Mol. Biol. Cell 2012, 23, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Deriu, M.A.; Soncini, M.; Orsi, M.; Patel, M.; Essex, J.W.; Montevecchi, F.M.; Redaelli, A. Anisotropic Elastic Network Modeling of Entire Microtubules. Biophy. J. 2010, 99, 2190–2199. [Google Scholar] [CrossRef] [PubMed]
- Schek, H.T.; Gardner, M.K.; Cheng, J.; Odde, D.J.; Hunt, A.J. Microtubule Assembly Dynamics at the Nanoscale. Curr. Biol. 2007, 17, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.; Charlebois, B.; Jánosi, I.; Howard, J.; Hunt, A.; Odde, D. Rapid Microtubule Self-Assembly Kinetics. Cell 2011, 146, 582–592. [Google Scholar] [CrossRef] [PubMed]


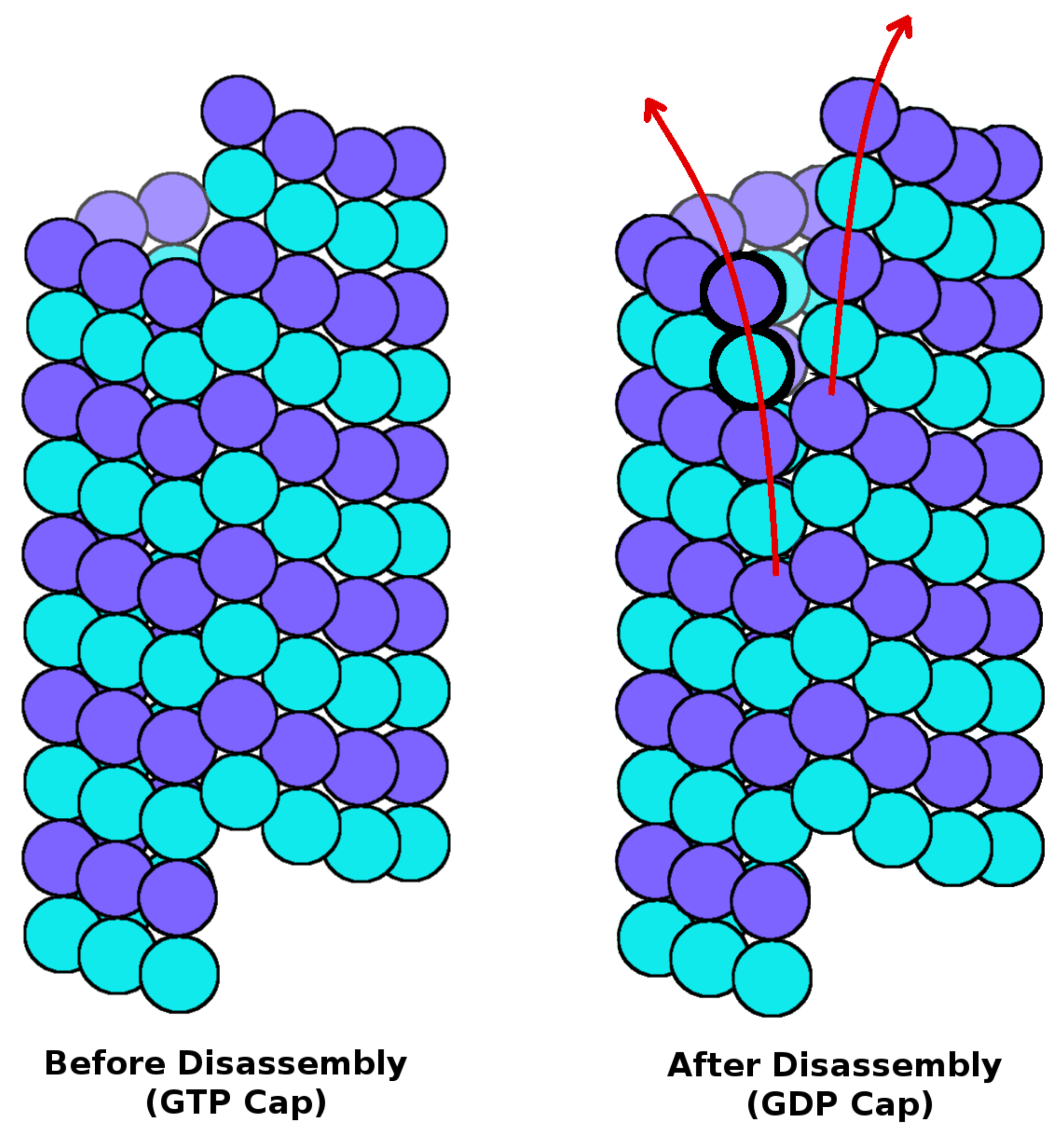{kind=link}
| Component | Before Disassembly | After Disassembly |
|---|---|---|
| Dipole–Dipole | 27 | 27 |
| vdW | 0 | |
| Electrostatic + GB | 84 | 92 |
| SASA | 0 | |
| Total | 119 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayoub, A.T.; Staelens, M.; Prunotto, A.; Deriu, M.A.; Danani, A.; Klobukowski, M.; Tuszynski, J.A. Explaining the Microtubule Energy Balance: Contributions Due to Dipole Moments, Charges, van der Waals and Solvation Energy. Int. J. Mol. Sci. 2017, 18, 2042. https://doi.org/10.3390/ijms18102042
Ayoub AT, Staelens M, Prunotto A, Deriu MA, Danani A, Klobukowski M, Tuszynski JA. Explaining the Microtubule Energy Balance: Contributions Due to Dipole Moments, Charges, van der Waals and Solvation Energy. International Journal of Molecular Sciences. 2017; 18(10):2042. https://doi.org/10.3390/ijms18102042
Chicago/Turabian StyleAyoub, Ahmed Taha, Michael Staelens, Alessio Prunotto, Marco A. Deriu, Andrea Danani, Mariusz Klobukowski, and Jack Adam Tuszynski. 2017. "Explaining the Microtubule Energy Balance: Contributions Due to Dipole Moments, Charges, van der Waals and Solvation Energy" International Journal of Molecular Sciences 18, no. 10: 2042. https://doi.org/10.3390/ijms18102042
APA StyleAyoub, A. T., Staelens, M., Prunotto, A., Deriu, M. A., Danani, A., Klobukowski, M., & Tuszynski, J. A. (2017). Explaining the Microtubule Energy Balance: Contributions Due to Dipole Moments, Charges, van der Waals and Solvation Energy. International Journal of Molecular Sciences, 18(10), 2042. https://doi.org/10.3390/ijms18102042