Protective Effect of Argan and Olive Oils against LPS-Induced Oxidative Stress and Inflammation in Mice Livers


 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of Argan Oil Pretreatment on LPS-Induced Metabolic Dysregulations of Plasmatic Parameters and Hepatotoxicity
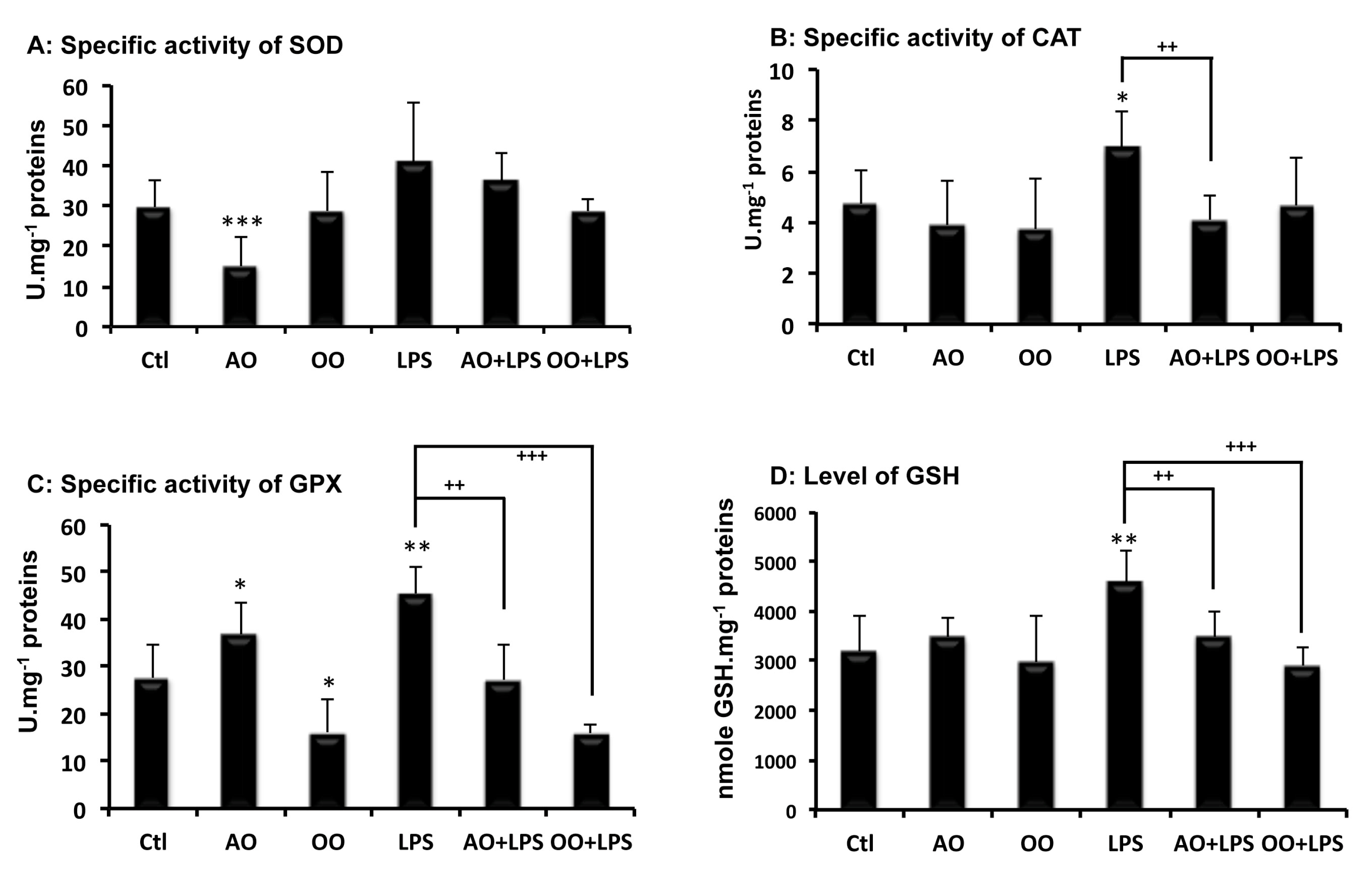
2.2. Effect of Argan Oil on LPS-Disturbed Antioxidant Activities in Mice Liver Tissues
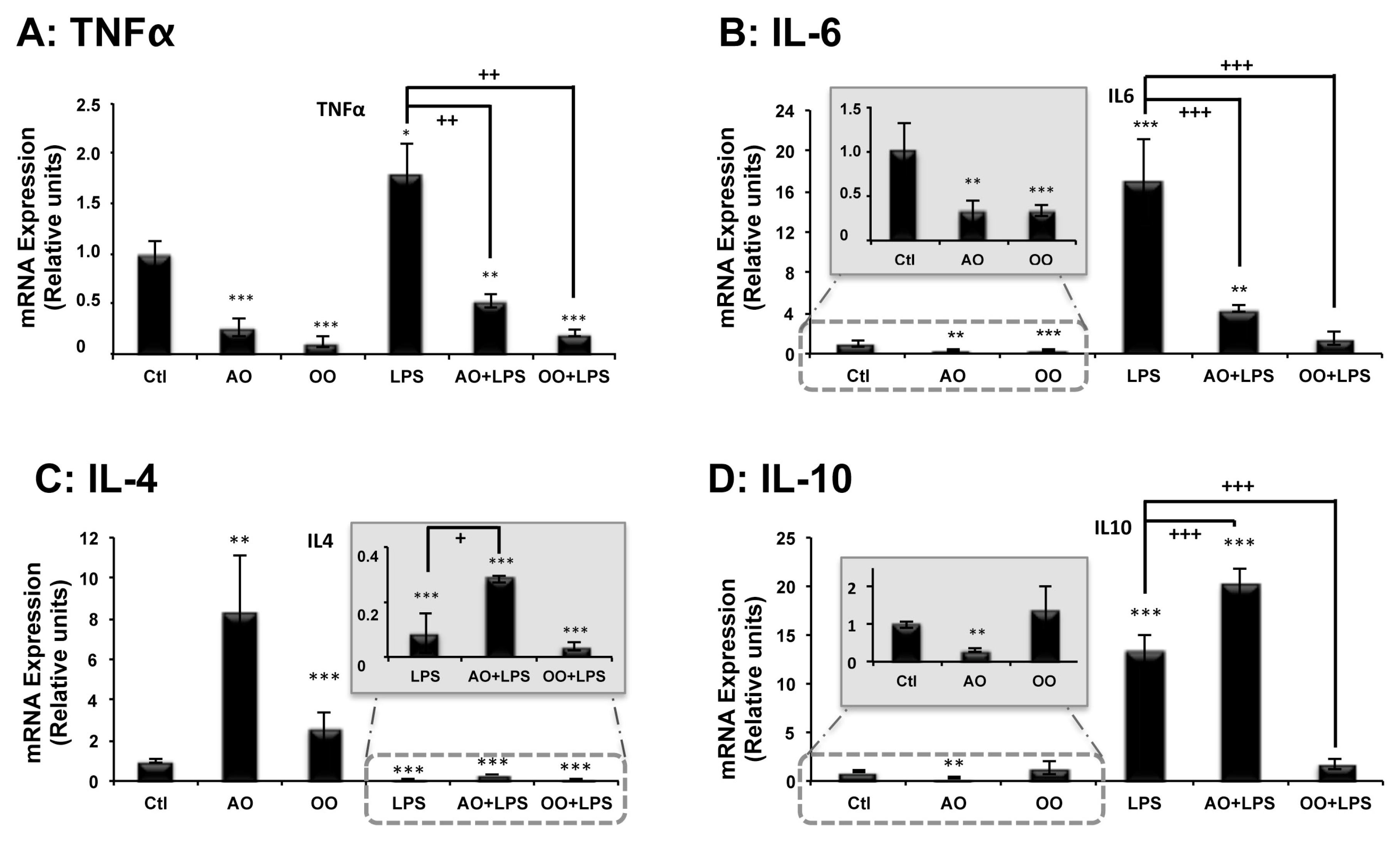
2.3. Effect of Argan Oil on LPS-Induced Inflammatory and Anti-Inflammatory Cytokines Disturbed Expression
3. Discussion
4. Material and Methods
4.1. Argan Oil Treatment
4.2. Preparation of Homogenates
4.3. Plasmatic Parameters Estimation
4.4. Reduced Glutathione Level
4.5. Catalase Activity
4.6. Superoxide Dismutase Activity
4.7. Glutathione Peroxidase Activity
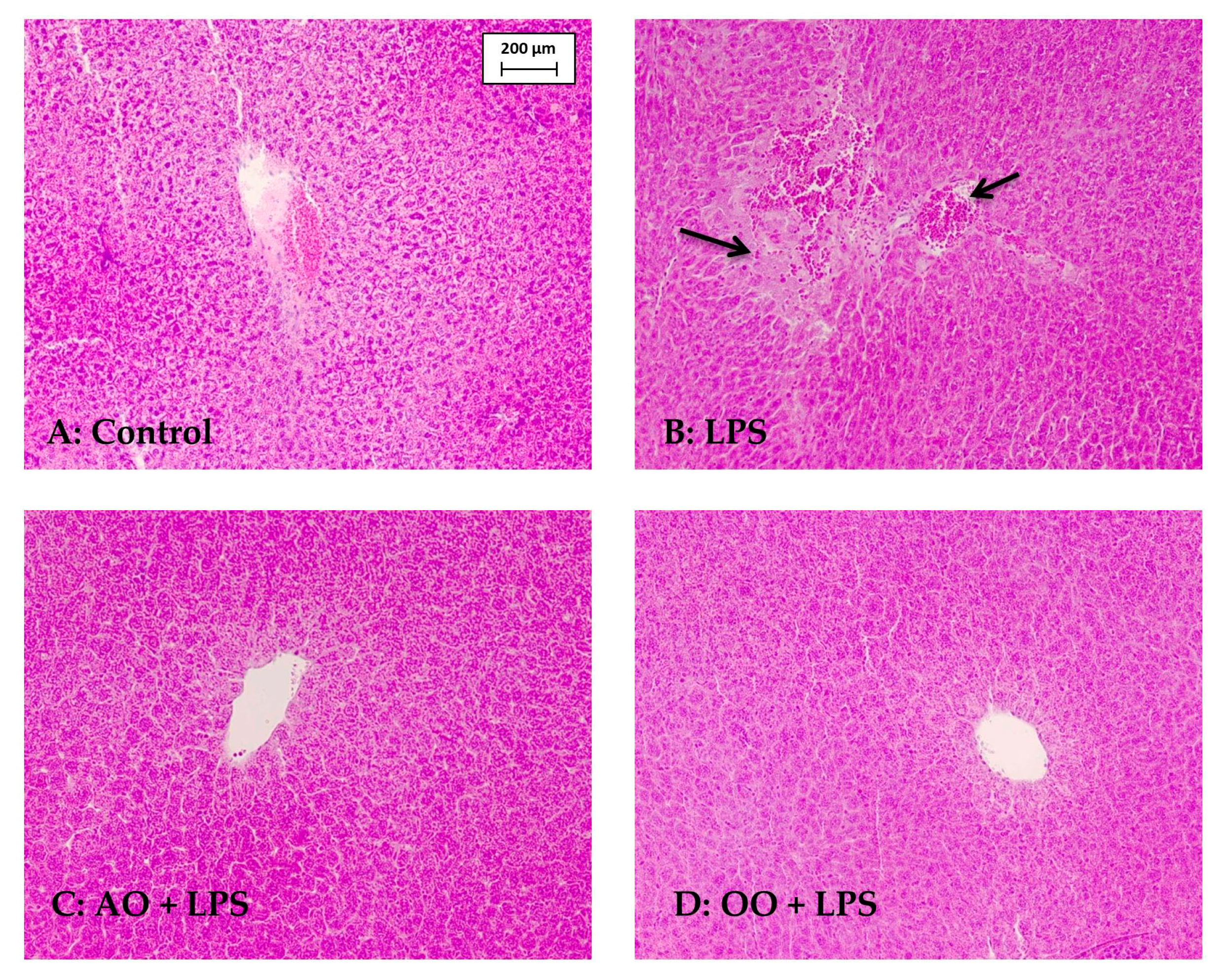
4.8. Histological Analysis
4.9. Quantitative PCR Analysis
4.10. Statistical Data Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maeder, M.; Fehr, T.; Rickli, H.; Ammann, P. Sepsis-associated myocardial dysfunction: Diagnostic and prognostic impact of cardiac troponins and natriuretic peptides. Chest 2006, 129, 1349–1366. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.L.; Sakr, Y.; Sprung, C.L.; Ranieri, V.M.; Reinhart, K.; Gerlach, H.; Moreno, R.; Carlet, J.; le Gall, J.R.; Payen, D. Sepsis in European intensive care units: Results of the SOAP study. Crit. Care Med. 2006, 34, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. The immunopathogenesis of sepsis. Nature 2002, 420, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Khovidhunkit, W.; Kim, M.S.; Memon, R.A.; Shigenaga, J.K.; Moser, A.H.; Feingold, K.R.; Grunfeld, C. Effects of infection and inflammation on lipid and lipoprotein metabolism: Mechanisms and consequences to the host. J. Lipid Res. 2004, 45, 1169–1196. [Google Scholar] [CrossRef] [PubMed]
- Sriskandan, S.; Altmann, D.M. The immunology of sepsis. J. Pathol. 2008, 214, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Spolarics, Z. Endotoxemia, pentose cycle, and the oxidant/antioxidant balance in the hepatic sinusoid. J. Leukoc. Biol. 1998, 63, 534–541. [Google Scholar] [PubMed]
- Kaplowitz, N.; Tsukamoto, H. Oxidative stress and liver disease. Prog. Liver Dis. 1996, 14, 131–159. [Google Scholar] [PubMed]
- Jaeschke, H. Mechanisms of oxidant stress-induced acute tissue injury. Proc. Soc. Exp. Biol. Med. 1995, 209, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Charrouf, Z.; Guillaume, D. Should the amazigh diet (regular and moderate argan-oil consumption) have a beneficial impact on human health? Crit. Rev. Food Sci. Nutr. 2010, 50, 473–477. [Google Scholar] [CrossRef] [PubMed]
- El Kebbaj, R.; El Kamouni, S.; El Hajj, H.I.; Andreoletti, P.; Gresti, J.; Latruffe, N.; El Kebbaj, M.S.; Vamecq, J.; Lizard, G.; Nasser, B.; et al. Modulation of peroxisomes abundance by argan oil and lipopolysaccharides in acyl-CoA oxidase 1-deficient fibroblasts. Health 2013, 5, 62–69. [Google Scholar] [CrossRef]
- Berrougui, H.; Cloutier, M.; Isabelle, M.; Khalil, A. Phenolic-extract from argan oil (Argania spinosa L.) inhibits human low-density lipoprotein (LDL) oxidation and enhances cholesterol efflux from human THP-1 macrophages. Atherosclerosis 2006, 184, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Cherki, M.; Derouiche, A.; Drissi, A.; El Messal, M.; Bamou, Y.; Idrissi-Ouadghiri, A.; Khalil, A.; Adlouni, A. Consumption of argan oil may have an antiatherogenic effect by improving paraoxonase activities and antioxidant status: Intervention study in healthy men. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Derouiche, A.; Cherki, M.; Drissi, A.; Bamou, Y.; El Messal, M.; Idrissi-Oudghiri, A.; Lecerf, J.M.; Adlouni, A. Nutritional intervention study with argan oil in man: Effects on lipids and apolipoproteins. Ann. Nutr. Metab. 2005, 49, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R.; Staprans, I.; Memon, R.A.; Moser, A.H.; Shigenaga, J.K.; Doerrler, W.; Dinarello, C.A.; Grunfeld, C. Endotoxin rapidly induces changes in lipid metabolism that produce hypertriglyceridemia: Low doses stimulate hepatic triglyceride production while high doses inhibit clearance. J. Lipid Res. 1992, 33, 1765–1776. [Google Scholar] [PubMed]
- Liu, M.S.; Spitzer, J.J. In vitro effects of E. coli endotoxin on fatty acid and lactate oxidation in canine myocardium. Circ. Shock 1977, 4, 181–190. [Google Scholar] [PubMed]
- Wang, X.; Evans, R.D. Effect of endotoxin and platelet-activating factor on lipid oxidation in the rat heart. J. Mol. Cell. Cadiol. 1997, 29, 1915–1926. [Google Scholar] [CrossRef] [PubMed]
- Zager, R.A.; Johnson, A.C.; Hanson, S.Y. Renal tubular triglyercide accumulation following endotoxic, toxic, and ischemic injury. Kidney Int. 2005, 67, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.J.; Yeh, W.C. Implication of Toll-like receptor and tumor necrosis factor α signaling in septic shock. Shock 2005, 24, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, S.; Wallace, F.A.; Calder, P.C. Dietary lipids modify the cytokine response to bacterial lipopolysaccharide in mice. Immunology 1999, 96, 404–410. [Google Scholar] [CrossRef] [PubMed]
- El Kebbaj, R.; Andreoletti, P.; El Hajj, H.I.; El Kharrassi, Y.; Vamecq, J.; Mandard, S.; Saih, F.-E.; Latruffe, N.; El Kebbaj, M.S.; Lizard, G.; et al. Argan oil prevents down-regulation induced by endotoxin on liver fatty acid oxidation and gluconeogenesis and on peroxisome proliferator-activated receptor gamma coactivator-1α, (PGC-1α), peroxisome proliferator-activated receptor α (PPARα) and estrogen related receptor α (ERRα). Biochimie Open 2015, 1, 51–59. [Google Scholar]
- Puertollano, M.A.; Puertollano, E.; Alvarez de Cienfuegos, G.; de Pablo, M.A. Significance of olive oil in the host immune resistance to infection. Br. J. Nutr. 2007, 98 (Suppl. 1), S54–S58. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Qu, D.; Zhang, Q.; Jiang, J.; Zhou, H.; Jiang, R.; Li, Y.; Zhang, Y.; Yan, H. Phytosterol esters attenuate hepatic steatosis in rats with non-alcoholic fatty liver disease rats fed a high-fat diet. Sci. Rep. 2017, 7, 41604. [Google Scholar] [CrossRef] [PubMed]
- Cadenas, S.; Cadenas, A.M. Fighting the stranger-antioxidant protection against endotoxin toxicity. Toxicology 2002, 180, 45–63. [Google Scholar] [CrossRef]
- Raetz, C.R. Biochemistry of endotoxins. Annu. Rev. Biochem. 1990, 59, 129–170. [Google Scholar] [CrossRef] [PubMed]
- Lichtenstein, A.H. Nutrient supplements and cardiovascular disease: A heartbreaking story. J. Lipid Res. 2009, 50 (Suppl.), S429–S433. [Google Scholar] [CrossRef] [PubMed]
- De Bock, M.; Derraik, J.G.; Brennan, C.M.; Biggs, J.B.; Morgan, P.E.; Hodgkinson, S.C.; Hofman, P.L.; Cutfield, W.S. Olive (Olea europaea L.) leaf polyphenols improve insulin sensitivity in middle-aged overweight men: A randomized, placebo-controlled, crossover trial. PLoS ONE 2013, 8, e57622. [Google Scholar] [CrossRef] [PubMed]
- Memon, R.A.; Grunfeld, C.; Moser, A.H.; Feingold, K.R. Tumor necrosis factor mediates the effects of endotoxin on cholesterol and triglyceride metabolism in mice. Endocrinology 1993, 132, 2246–2253. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Diomede, L.; Albani, D.; Bianchi, M.; Salmona, M. Endotoxin regulates the maturation of sterol regulatory element binding protein-1 through the induction of cytokines. Eur. Cytokine Netw. 2001, 12, 625–630. [Google Scholar] [PubMed]
- Yvan-Charvet, L.; Wang, N.; Tall, A.R. Role of HDL, ABCA1, and ABCG1 transporters in cholesterol efflux and immune responses. Arterioscl. Thromb. Vascul. Biol. 2010, 30, 139–143. [Google Scholar] [CrossRef] [PubMed]
- El Kharrassi, Y.; Samadi, M.; Lopez, T.; Nury, T.; El Kebbaj, R.; Andreoletti, P.; El Hajj, H.I.; Vamecq, J.; Moustaid, K.; Latruffe, N.; et al. Biological activities of Schottenol and Spinasterol, two natural phytosterols present in argan oil and in cactus pear seed oil, on murine miroglial BV2 cells. Biochem. Biophys. Res. Commun. 2014, 446, 798–804. [Google Scholar] [CrossRef] [PubMed]
- Zelcer, N.; Tontonoz, P. Liver X receptors as integrators of metabolic and inflammatory signaling. J. Clin. Investig. 2006, 116, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Mensink, R.P. Increased intestinal ABCA1 expression contributes to the decrease in cholesterol absorption after plant stanol consumption. Faseb. J. 2002, 16, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Valerio, M.S.; Minderman, H.; Mace, T.; Awad, A.B. β-Sitosterol modulates TLR4 receptor expression and intracellular MyD88-dependent pathway activation in J774A.1 murine macrophages. Cell. Immunol. 2013, 285, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Cambi, V.; David, S. Basic therapeutic requirements in the treatment of sepsis in acute renal failure. Nephrol. Dial. Transplant. 1994, 9 (Suppl. 4), 183–186. [Google Scholar] [PubMed]
- Mitra, S.K.; Venkataranganna, M.V.; Sundaram, R.; Gopumadhavan, S. Protective effect of HD-03, a herbal formulation, against various hepatotoxic agents in rats. J. Ethnopharmacol. 1998, 63, 181–186. [Google Scholar] [CrossRef]
- Drissi, A.; Girona, J.; Cherki, M.; Godas, G.; Derouiche, A.; El Messal, M.; Saile, R.; Kettani, A.; Sola, R.; Masana, L.; et al. Evidence of hypolipemiant and antioxidant properties of argan oil derived from the argan tree (Argania spinosa). Clin. Nutr. 2004, 23, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.R.; Flamm, S.L.; di Bisceglie, A.M.; Bodenheimer, H.C.; Public Policy Committee of the American Association for the Study of Liver Disease. Serum activity of alanine aminotransferase (ALT) as an indicator of health and disease. Hepatology 2008, 47, 1363–1370. [Google Scholar] [CrossRef] [PubMed]
- Nathwani, R.A.; Pais, S.; Reynolds, T.B.; Kaplowitz, N. Serum alanine aminotransferase in skeletal muscle diseases. Hepatology 2005, 41, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Cori, C.F. The glucose-lactic acid cycle and gluconeogenesis. Curr. Top. Cell. Regul. 1981, 18, 377–387. [Google Scholar] [PubMed]
- Felig, P. The glucose-alanine cycle. Metabolism 1973, 22, 179–207. [Google Scholar] [CrossRef]
- Katz, J.; Tayek, J.A. Gluconeogenesis and the Cori cycle in 12-, 20-, and 40-h-fasted humans. Am. J. Physiol. 1998, 275, E537–E542. [Google Scholar] [PubMed]
- El-Tanbouly, D.M.; Abdelsalam, R.M.; Attia, A.S.; Abdel-Aziz, M.T. Pretreatment with magnesium ameliorates lipopolysaccharide-induced liver injury in mice. Pharmacol. Rep. 2015, 67, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Galli, C. The role of antioxidants in the Mediterranean diet. Lipids 2001, 36, S49–S52. [Google Scholar] [CrossRef] [PubMed]
- Papa, S.; Bubici, C.; Zazzeroni, F.; Franzoso, G. Mechanisms of liver disease: Cross-talk between the NF-κB and JNK pathways. Biol. Chem. 2009, 390, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.S.; Riehle, K.J.; Brooling, J.T.; Bauer, R.L.; Mitchell, C.; Fausto, N. Proinflammatory cytokine production in liver regeneration is Myd88-dependent, but independent of Cd14, Tlr2, and Tlr4. J. Immunol. 2006, 176, 2522–2528. [Google Scholar] [CrossRef] [PubMed]
- Kono, K.; Kamijo, Y.; Hora, K.; Takahashi, K.; Higuchi, M.; Kiyosawa, K.; Shigematsu, H.; Gonzalez, F.J.; Aoyama, T. PPARα attenuates the proinflammatory response in activated mesangial cells. Am. J. Physiol. Renal. Physiol. 2009, 296, F328–F336. [Google Scholar] [CrossRef] [PubMed]
- Zambon, A.; Gervois, P.; Pauletto, P.; Fruchart, J.C.; Staels, B. Modulation of hepatic inflammatory risk markers of cardiovascular diseases by PPAR-α activators: Clinical and experimental evidence. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Opal, S.M.; DePalo, V.A. Anti-inflammatory cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Ricardo-Gonzalez, R.R.; Red Eagle, A.; Odegaard, J.I.; Jouihan, H.; Morel, C.R.; Heredia, J.E.; Mukundan, L.; Wu, D.; Locksley, R.M.; Chawla, A. IL-4/STAT6 immune axis regulates peripheral nutrient metabolism and insulin sensitivity. Proc. Natl. Acad. Sci. USA 2010, 107, 22617–22622. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A. Control of macrophage activation and function by PPARs. Circ. Res. 2010, 106, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, D.F.; Zlotnik, A.; Mosmann, T.R.; Howard, M.; O’Garra, A. IL-10 inhibits cytokine production by activated macrophages. J. Immunol. 1991, 147, 3815–3822. [Google Scholar] [PubMed]
- Clarke, C.J.; Hales, A.; Hunt, A.; Foxwell, B.M. IL-10-mediated suppression of TNF-α production is independent of its ability to inhibit NF κB activity. Eur. J. Immunol. 1998, 28, 1719–1726. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Cherkaoui Malki, M.; Bardot, O.; Lhuguenot, J.C.; Latruffe, N. Expression of liver peroxisomal proteins as compared to other organelle marker enzymes in rats treated with hypolipidemic agents. Biol. Cell 1990, 69, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Flohe, L.; Gunzler, W.A. Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar] [PubMed]
- Vluggens, A.; Andreoletti, P.; Viswakarma, N.; Jia, Y.; Matsumoto, K.; Kulik, W.; Khan, M.; Huang, J.; Guo, D.; Yu, S.; et al. Reversal of mouse Acyl-CoA oxidase 1 (ACOX1) null phenotype by human ACOX1b isoform [corrected]. Lab. Investig. 2010, 90, 696–708. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Plasma Parameters | Control | LPS | AO | AO + LPS | OO | OO + LPS |
|---|---|---|---|---|---|---|
| Glycaemia (mg/dL) | 2.91 ± 0.08 | 4.63 ± 0.6 ** | 2.28 ± 0.21 * | 3.7 ± 0.5 ** | 3.56 ± 0.23 * | 3.48 ± 0.39 ## |
| 100% | 159% a | 78.3% a | 80% b | 122% a | 75.2% b | |
| Cholesterol (mg/dL) | 1.97 ± 0.11 | 2.33 ± 0.28* | 1.96 ± 0.13 | 2.07 ± 0.06 | 1.66 ± 0.15 * | 1.02 ± 0.03 ***## |
| 100% | 118% a | 99.5% a | 88.8% b | 84.3% a | 43.8% b | |
| Proteins (mg/dL) | 48.17 ± 7.89 | 68.15 ± 12.8 * | 42.60 ± 2.67 | 45.75 ± 0.9 # | 51.83 ± 5.8 | 38.80 ± 3.39 # |
| 100% | 141% a | 88.4% a | 67.1% b | 107.6% a | 56.9% b | |
| Uremia (mmol/dL) | 6.35 ± 0.07 | 23.17±2.29 *** | 6.6 ± 0.36 | 19.18 ±1.18 ***# | 4.80 ± 0.99 | 16.95 ± 0.07 ***## |
| 100% | 365% a | 104% a | 82.8% b | 75.6% a | 73.15% b | |
| AST (U/mL) | 395.5 ± 7.78 | 480.8 ± 11.6 *** | 387.1 ± 0.71 | 295 ± 8.49 ***### | 329.95 ± 13.79 ** | 292 ± 16.97 ***### |
| 100% | 121.6% a | 97.9% a | 61.4% b | 83.4% a | 60.7% b | |
| ALT (U/mL) | 43.45 ± 2.19 | 72.9 ± 1.56 *** | 63.03 ± 1.78 *** | 47.3 ± 5.80 ### | 37.9 ± 3.96 | 48.05 ± 10.11 ## |
| 100% | 167.8% a | 145% a | 64.9% b | 87.2% a | 65.9% b | |
| ALT to AST % increase ratio | 1 | 1.38 | 1.48 | 1.05 | 1.05 | 1.09 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Kamouni, S.; El Kebbaj, R.; Andreoletti, P.; El Ktaibi, A.; Rharrassi, I.; Essamadi, A.; El Kebbaj, M.S.; Mandard, S.; Latruffe, N.; Vamecq, J.; et al. Protective Effect of Argan and Olive Oils against LPS-Induced Oxidative Stress and Inflammation in Mice Livers. Int. J. Mol. Sci. 2017, 18, 2181. https://doi.org/10.3390/ijms18102181
El Kamouni S, El Kebbaj R, Andreoletti P, El Ktaibi A, Rharrassi I, Essamadi A, El Kebbaj MS, Mandard S, Latruffe N, Vamecq J, et al. Protective Effect of Argan and Olive Oils against LPS-Induced Oxidative Stress and Inflammation in Mice Livers. International Journal of Molecular Sciences. 2017; 18(10):2181. https://doi.org/10.3390/ijms18102181
Chicago/Turabian StyleEl Kamouni, Soufiane, Riad El Kebbaj, Pierre Andreoletti, Abderrahim El Ktaibi, Issam Rharrassi, Abdelkhalid Essamadi, M’hammed Saïd El Kebbaj, Stéphane Mandard, Norbert Latruffe, Joseph Vamecq, and et al. 2017. "Protective Effect of Argan and Olive Oils against LPS-Induced Oxidative Stress and Inflammation in Mice Livers" International Journal of Molecular Sciences 18, no. 10: 2181. https://doi.org/10.3390/ijms18102181
APA StyleEl Kamouni, S., El Kebbaj, R., Andreoletti, P., El Ktaibi, A., Rharrassi, I., Essamadi, A., El Kebbaj, M. S., Mandard, S., Latruffe, N., Vamecq, J., Nasser, B., & Cherkaoui-Malki, M. (2017). Protective Effect of Argan and Olive Oils against LPS-Induced Oxidative Stress and Inflammation in Mice Livers. International Journal of Molecular Sciences, 18(10), 2181. https://doi.org/10.3390/ijms18102181