Argan Oil-Mediated Attenuation of Organelle Dysfunction, Oxidative Stress and Cell Death Induced by 7-Ketocholesterol in Murine Oligodendrocytes 158N



 ,
, 
Abstract
:
1. Introduction
2. Results
2.1. Fatty Acid, Phytosterol, Tocopherol and Polyphenol Profile of Argan Oils from Morocco and Extra Virgin Olive Oil from Tunisia
2.2. Comparison of the Antioxidant Properties of Argan Oils from Morocco and Extra Virgin Olive Oil from Tunisia Using Two Complementary Techniques: The KRL and FRAP Tests
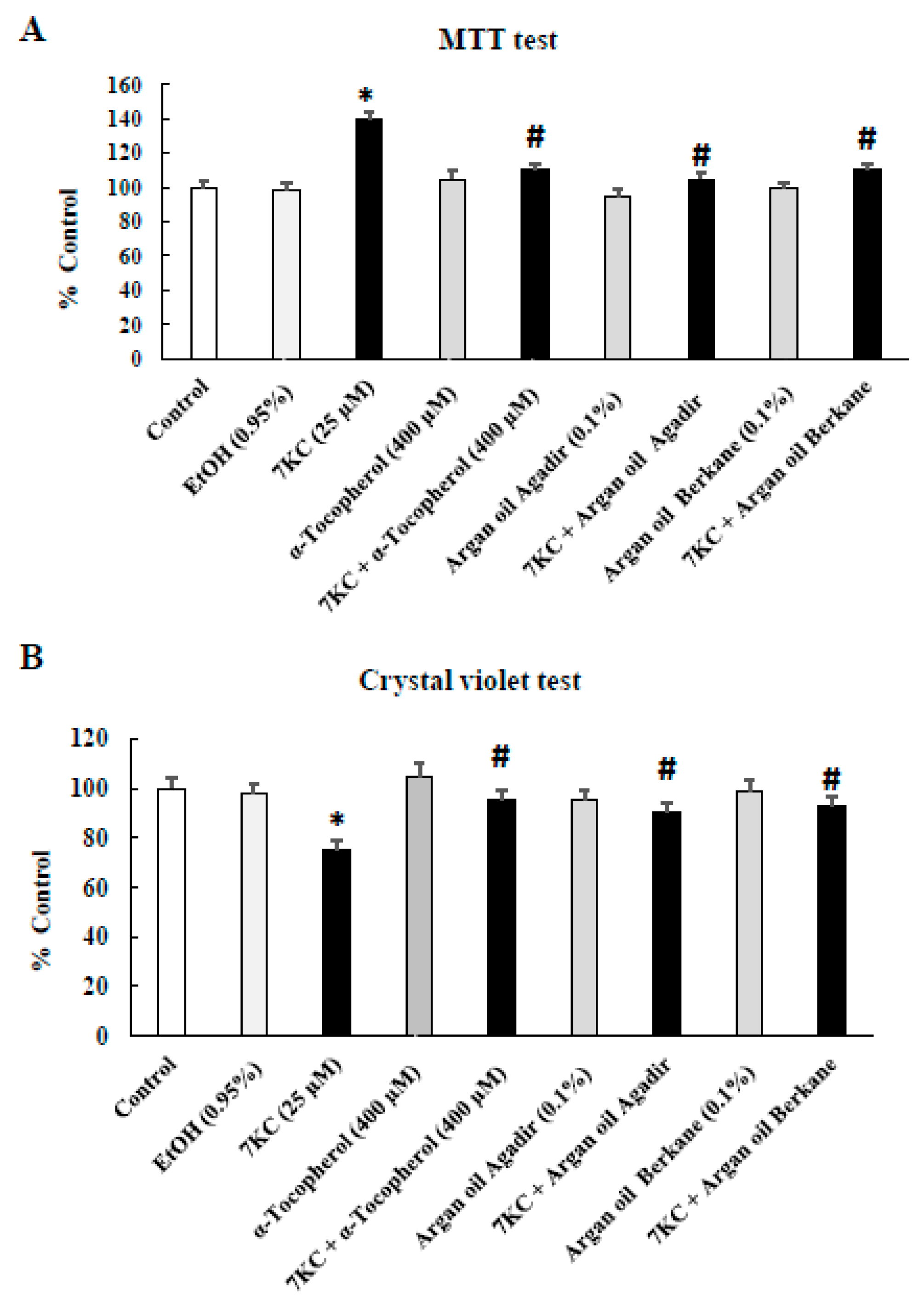
2.3. Evaluation of the Effects of Argan Oils on 7-Ketocholesterol-Induced Cell-Growth Inhibition Evaluated with the Crystal Violet and MTT Tests and by Phase-Contrast Microscopy
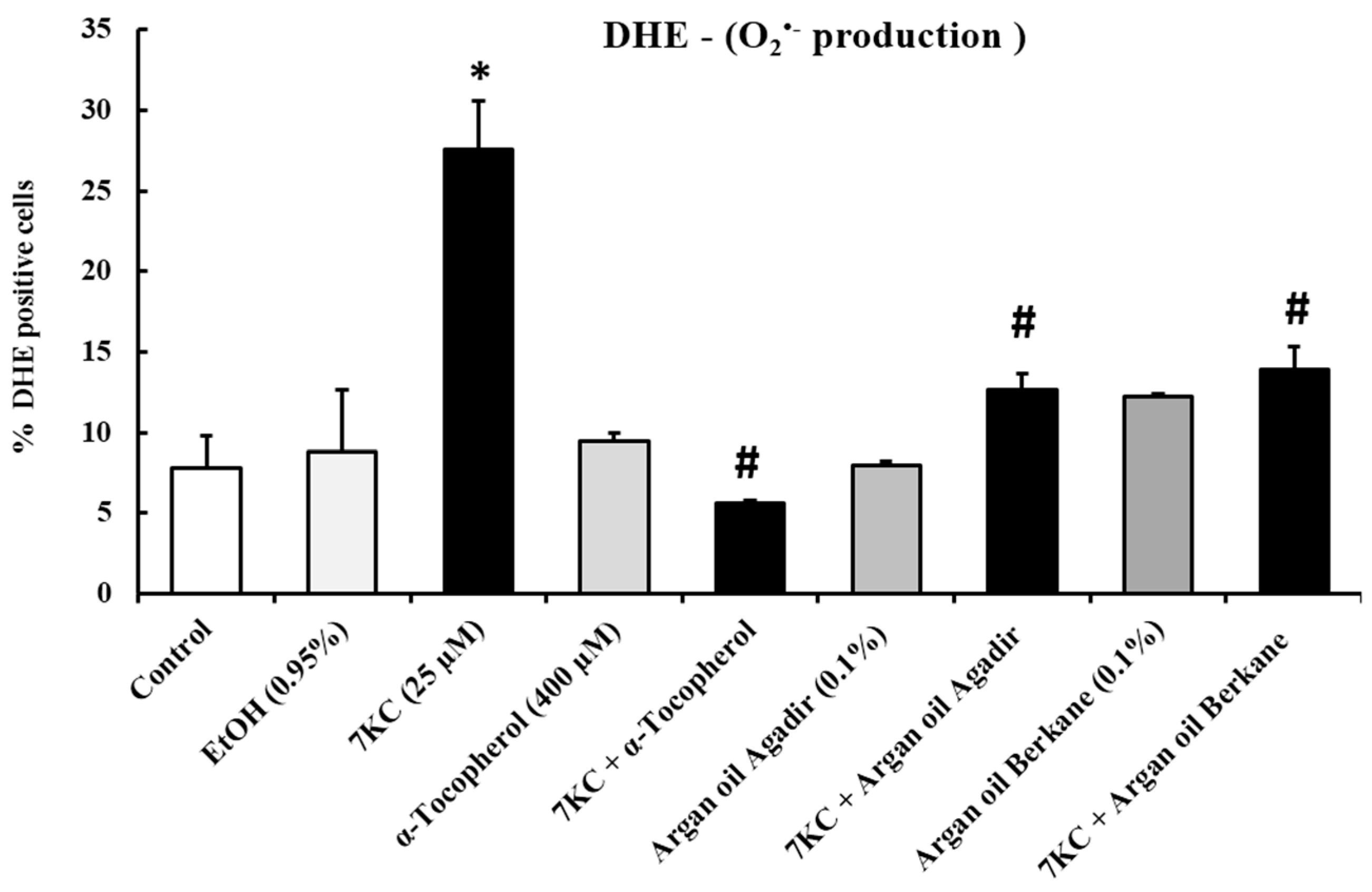
2.4. Evaluation of the Effects of Argan Oils on 7-Ketocholesterol-Induced Cell Overproduction of Reactive Oxygen Species: Measurement by Flow Cytometry after Staining with Dihydroethidine
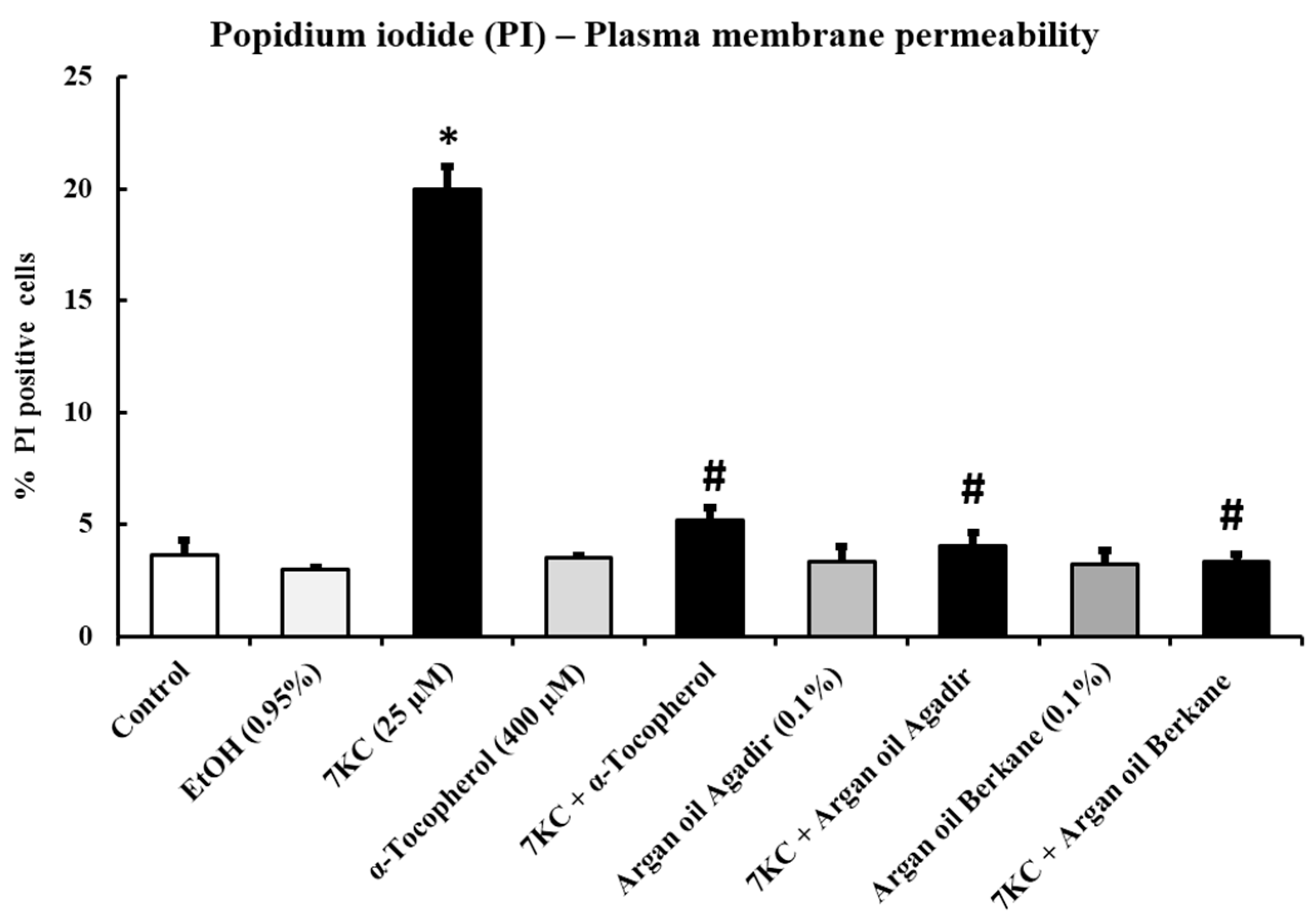
2.5. Evaluation of the Effects of Argan Oils on 7-Ketocholesterol-Induced Increased Plasma Membrane Permeability: Measurement by Flow Cytometry after Staining with Propidium Iodide
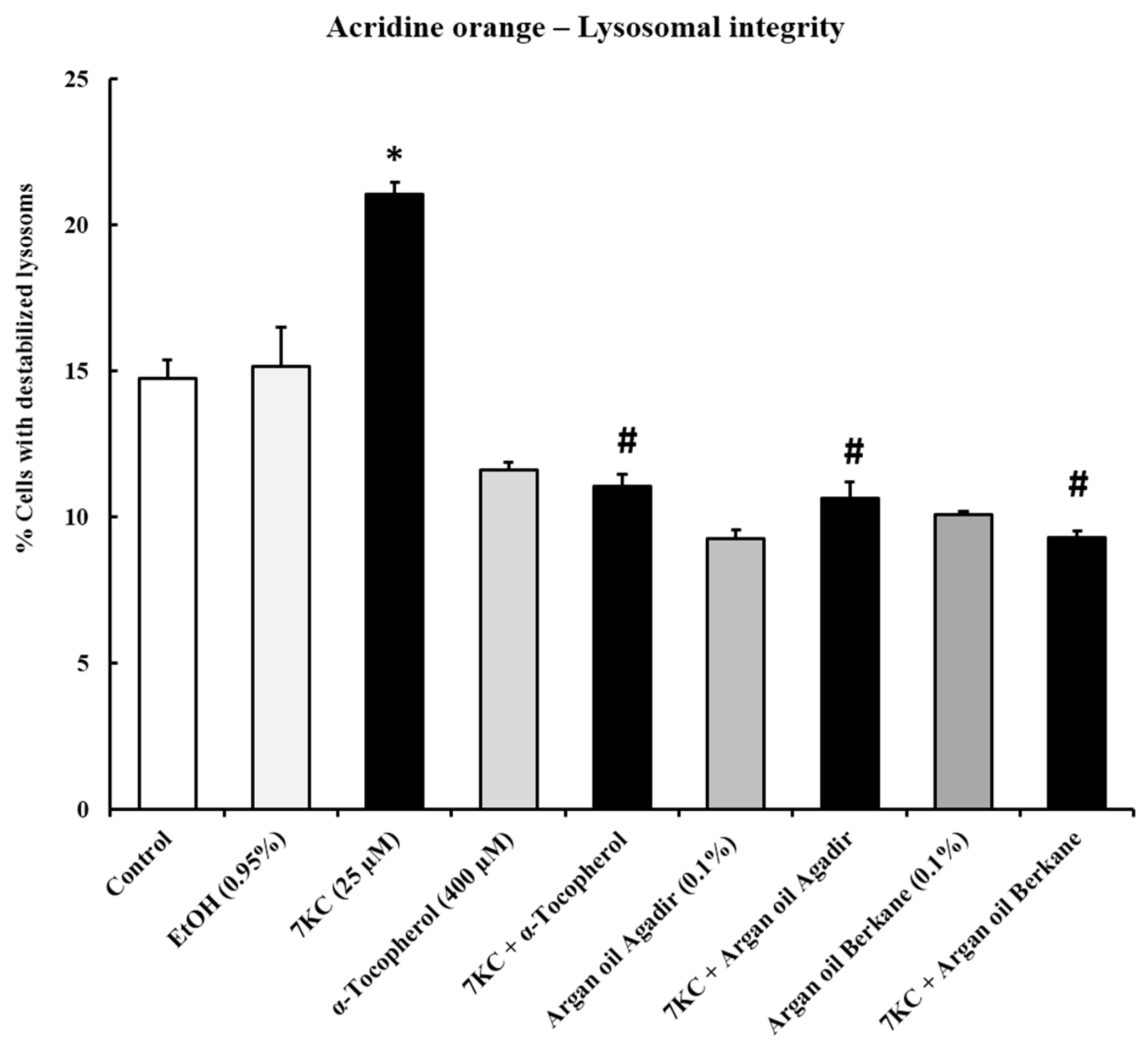
2.6. Evaluation of the Effects of Argan Oils on 7-Ketocholesterol-Induced Acidic Vesicle Formation: Measurement by Flow Cytometry after Staining with Acridine Orange
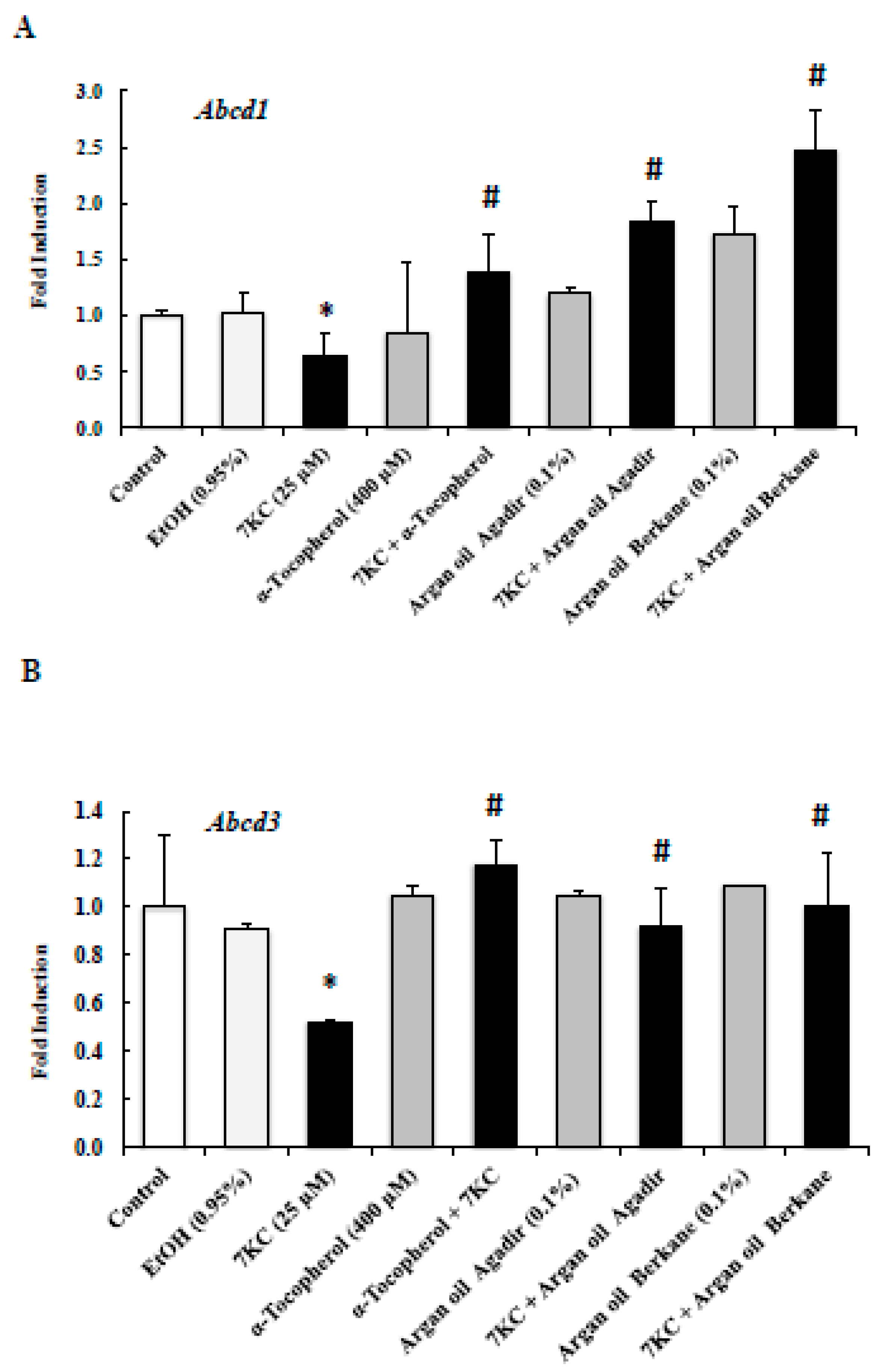
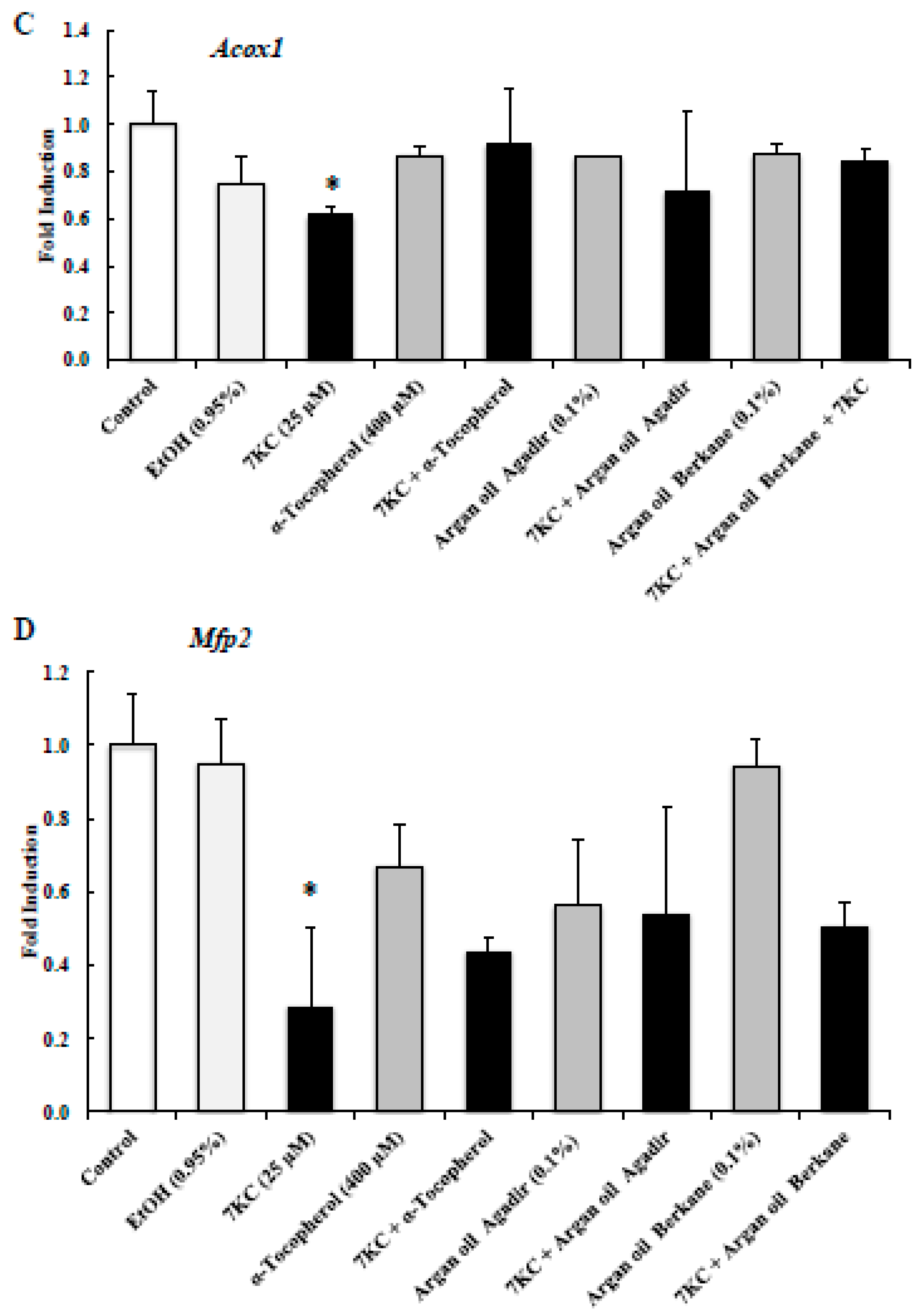
2.7. Evaluation of the Effects of Argan Oils on 7KC-Induced Peroxisomal Dysfunction Evaluated by Abcd1, Abcd3, Acox1, and Mfp2 mRNA Levels
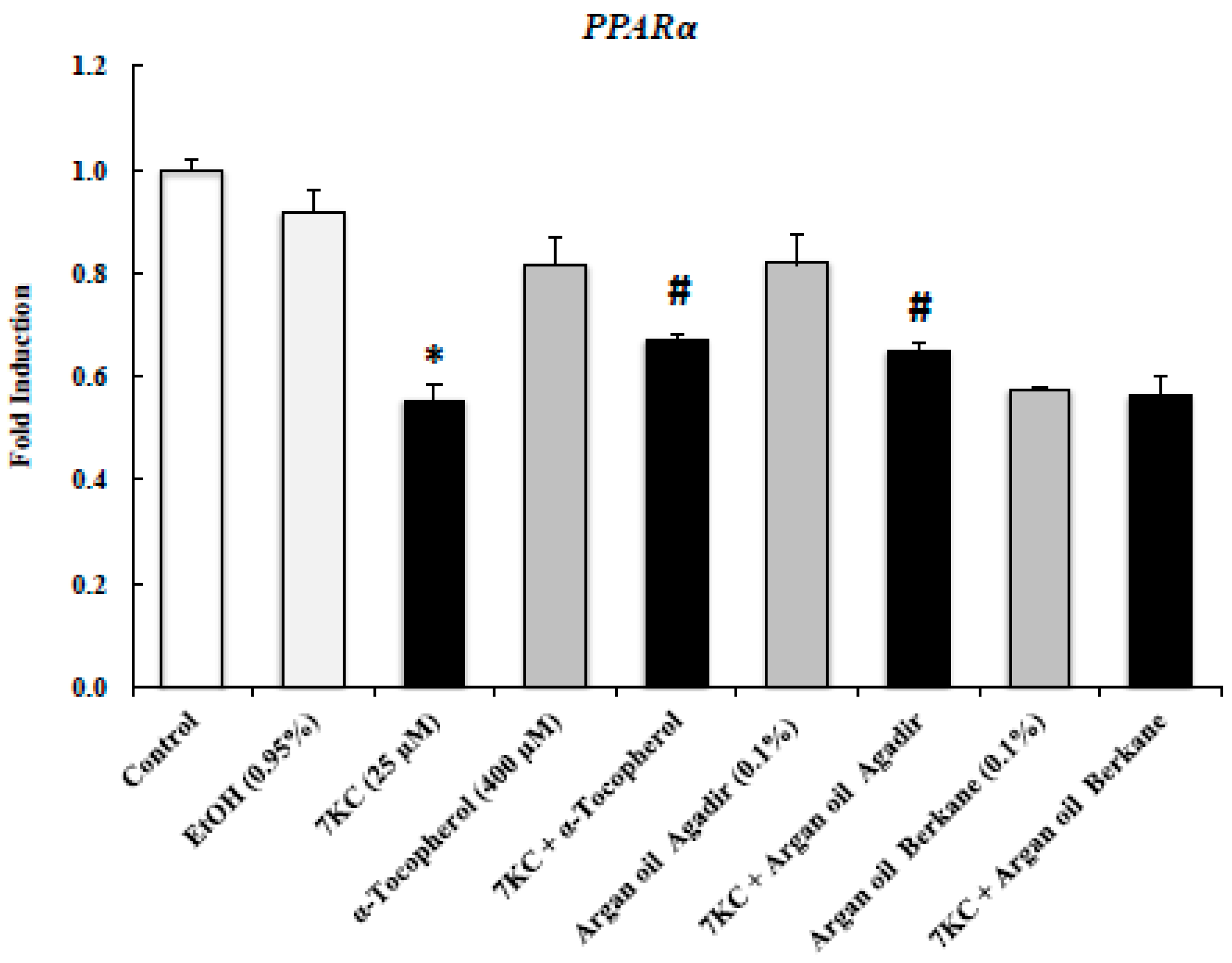
2.8. Evaluation of the Effects of Argan Oils on 7KC-Induced Decreased Transcription of PPARα mRNA Level
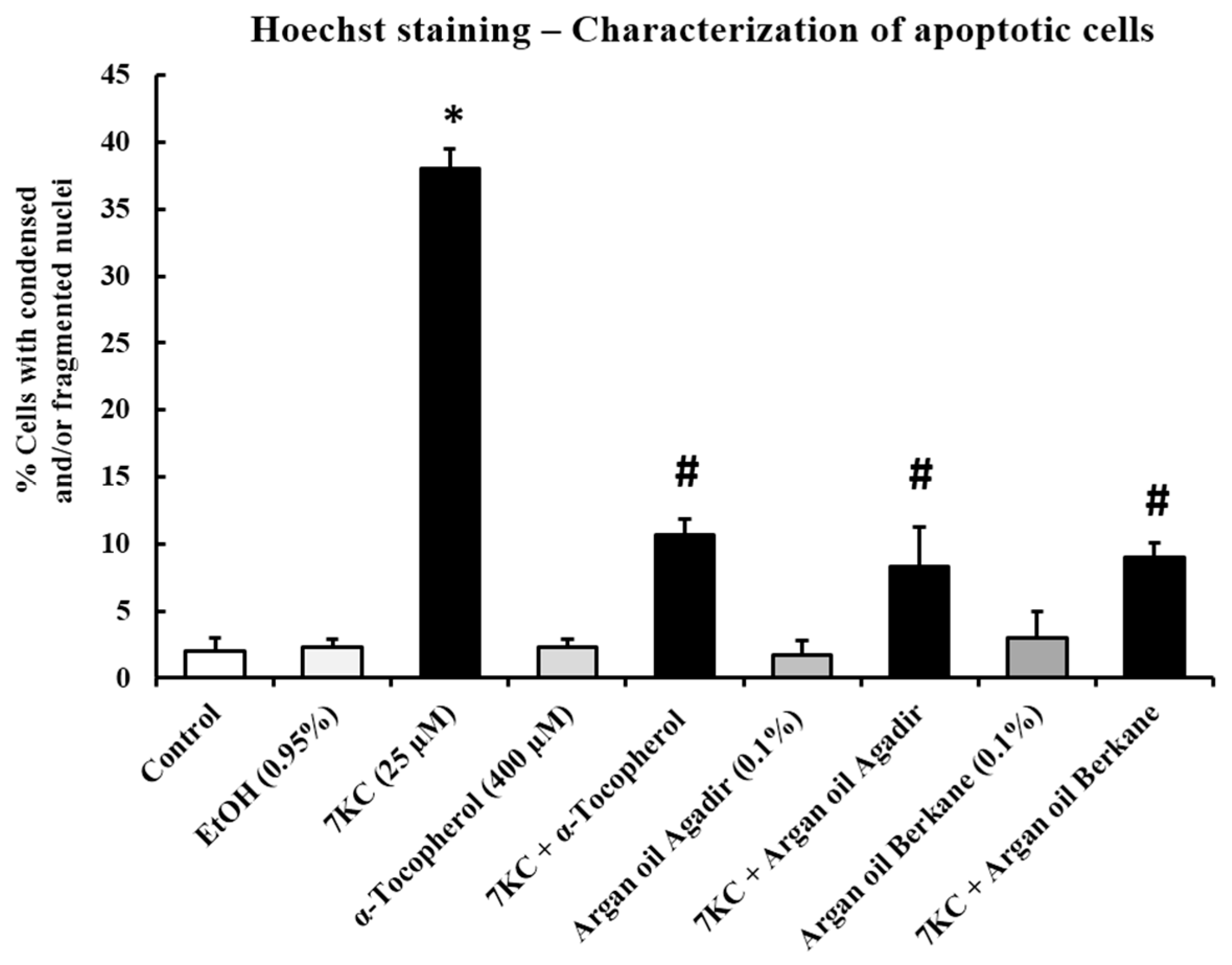
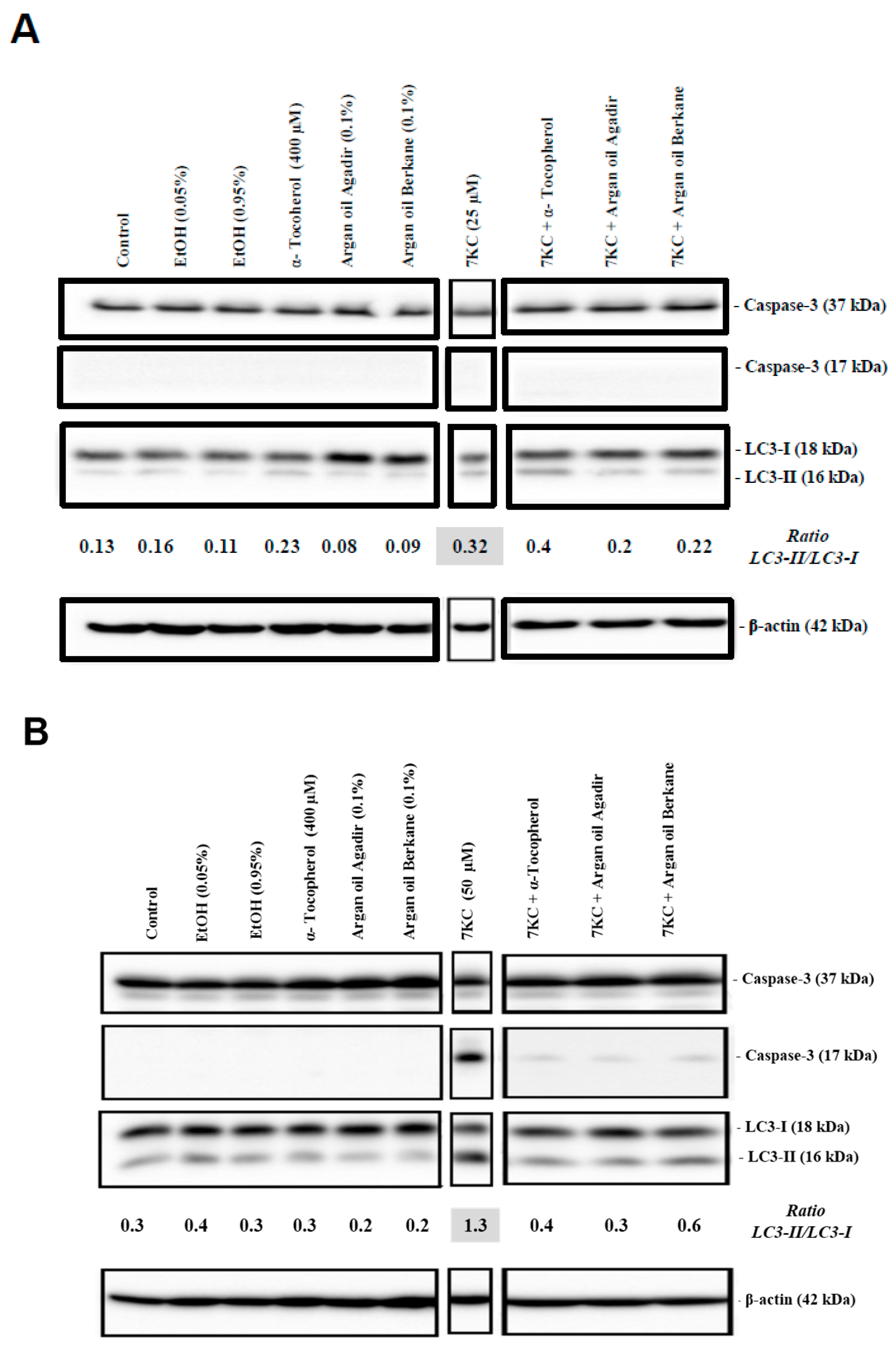
2.9. Evaluation of the Effects of Argan Oils on 7KC-Induced Apoptosis and Autophagy
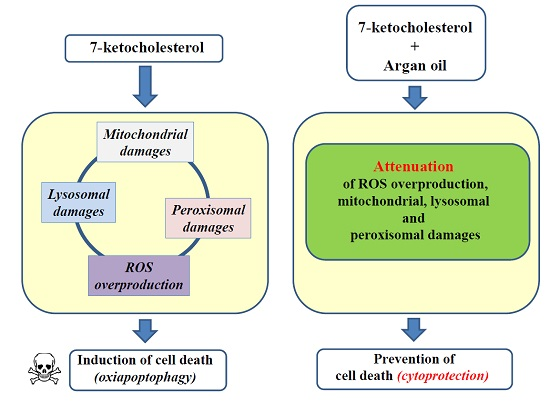
3. Discussion
4. Experimental Section
4.1. Cell Culture and Treatments
4.2. Determination of the Fatty Acid Profile of Argan Oil Versus Extra Virgin Olive Oil by Gas Chromatography
4.3. Determination of the Tocopherol Profile of Argan Oil versus Extra Virgin Olive Oil by High Pressure Liquid Chromatography
4.4. Extraction by Organic Solvent
4.4.1. Plant Material
4.4.2. Preparation of Crude, Lipidic and Non-Lipidic Extracts
4.5. KRL Test
4.6. Ferric Reducing Antioxidant Power (FRAP) Assay
4.7. Crystal Violet Test
4.8. Measurement of Mitochondrial Activity with the MTT Test
4.9. Measurement of Acidic Vesicles with Acridine Orange
4.10. Measurement of Superoxide Anion Production with Dihydroethidium
4.11. Measurement of Plasma Membrane Permeability with Propidium Iodide
4.12. Quantification of Apoptotic Cells after Staining Nuclei with Hoechst 33342
4.13. Quantification of Abcd1, Abcd2, Abcd3, Acox1, Mfp2 and PPARα mRNAs by RT-qPCR
- Abcd1: forward 5′-gccaagttgtggatgag-3′ and reverse 5′-ttccgcagagtcgggataga-3′
- Abcd2: forward 5′-tagaccgcatcctgcacagc-3′ and reverse 5′-ctccttcgccatcgaattgt-3′
- Abcd3: forward 5′-ctgggcgtgaaatgactagattg-3′ and reverse 5′-cttctcctgttgtgacaccattg-3′
- Acox1: forward 5′-gcccaactgtgacttccatt-3′ and reverse 5′-ggcatgtaacccgtagcact-3′
- Mfp2: forward 5′-aggggacttcaagggaattgg-3′ and reverse 5′-gcctgcttcaactgaatcgtaa-3′
- PPARα: forward 5′-tattcggctgaagctggtgtac-3′ and reverse 5′-ctggcatttgttccggttct-3′
4.14. Analysis of Caspase-3 and LC3 by Polyacrylamide Gel Electrophoresis and Western Blotting
4.15. Statistical Analyses
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- German, J.B.; Dillard, C.J. Composition, structure and absorption of milk lipids: A source of energy, fat-soluble nutrients and bioactive molecules. Crit. Rev. Food Sci. Nutr. 2006, 46, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, F.; Cruz-Hernandez, C.; Flück, B.; Tavazzi, I.; Thakkar, S.K.; Destaillats, F.; Braun, M. Quantification of phospholipids classes in human milk. Lipids 2013, 48, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Kamelska, A.M.; Pietrzak-Fiećko, R.; Bryl, K. Variation of the cholesterol content in breast milk during 10 days collection at early stages of lactation. Acta Biochim. Pol. 2012, 59, 243–247. [Google Scholar] [PubMed]
- Delplanque, B.; Gibson, R.; Koletzko, B.; Lapillonne, A.; Strandvik, B. Lipid Quality in Infant Nutrition: Current Knowledge and Future Opportunities. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Gow, R.V.; Hibbeln, J.R. Omega-3 fatty acid and nutrient deficits in adverse neurodevelopment and childhood behaviors. Child Adolesc. Psychiatr. Clin. N. Am. 2014, 23, 555–590. [Google Scholar] [CrossRef] [PubMed]
- Zarrouk, A.; Debbabi, M.; Bezine, M.; Karym, E.M.; Badreddine, A.; Rouaud, O.; Moreau, T.; Cherkaoui-Malki, M.; El Ayeb, M.; Nasser, B.; et al. Lipid Biomarkers in Alzheimer’s Disease. Curr. Alzheimer. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wanders, R.J.; Waterham, H.R. Biochemistry of mammalian peroxisomes revisited. Annu. Rev. Biochem. 2006, 75, 295–332. [Google Scholar] [CrossRef] [PubMed]
- Ademowo, O.S.; Dias, H.K.I.; Burton, D.G.A.; Griffiths, H.R. Lipid (per) oxidation in mitochondria: An emerging target in the ageing process? Biogerontology 2017. [Google Scholar] [CrossRef] [PubMed]
- Zarrouk, A.; Cheraif, I.; Hadj-Ahmed, S.; Chaabane, W.; Hammami, S.; Debbabi, M.; Frih, M.; Rouaud, O.; Moreau, T.; Lizard, G.; et al. Correlations between Peripheral Trans Fatty Acids, Lipid Peroxidation Markers and Cognition in Dementia. Biochem. Anal. Biochem. 2015, 4, 4. [Google Scholar]
- Abate, G.; Marziano, M.; Rungratanawanich, W.; Memo, M.; Uberti, D. Nutrition and AGE-ing: Focusing on Alzheimer’s Disease. Oxid. Med. Cell. Longev. 2017, 2017, 7039816. [Google Scholar] [CrossRef] [PubMed]
- Eckert, G.P.; Lipka, U.; Muller, W.E. Omega-3 fatty acids in neurodegenerative diseases: Focus on mitochondria. Prostaglandins Leukot. Essent. Fatty Acids 2013, 88, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Tangney, C.C. Dietary fat composition and dementia risk. Neurobiol. Aging 2014, 2, S59–S64. [Google Scholar] [CrossRef] [PubMed]
- Björkhem, I. Crossing the barrier: Oxysterols as cholesterol transporters and metabolic modulators in the brain. J. Intern. Med. 2006, 260, 493–508. [Google Scholar] [CrossRef] [PubMed]
- Zarrouk, A.; Vejux, A.; Mackrill, J.; O’Callaghan, Y.; Hammami, M.; O’Brien, N.; Lizard, G. Involvement of oxysterols in age-related diseases and ageing processes. Ageing Res. Rev. 2014, 18, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Aoun, M.; Tiranti, V. Mitochondria: A crossroads for lipid metabolism defect in neurodegeneration with brain iron accumulation diseases. Int. J. Biochem. Cell Biol. 2015, 63, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, S.; Rabinovitz, S. The role of essential fatty acids in anorexia nervosa and obesity. Crit. Rev. Food Sci. Nutr. 2016, 56, 2021–2035. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Pelaez, R.; Lukiw, W.J.; Bazan, N.G. Omega-3 essential fatty acids modulate initiation and progression of neurodegenerative disease. Mol. Neurobiol. 2010, 41, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Karr, J.E.; Alexander, J.E.; Winningham, R.G. Omega-3 polyunsaturated fatty acids and cognition throughout the lifespan: A review. Nutr. Neurosci. 2011, 14, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Barnard, N.D.; Bush, A.I.; Ceccarelli, A.; Cooper, J.; de Jager, C.A.; Erickson, K.I.; Fraser, G.; Kesler, S.; Levin, S.M.; Lucey, B.; et al. Dietary and lifestyle guidelines for the prevention of Alzheimer’s disease. Neurobiol. Aging 2014, 35, S74–S78. [Google Scholar] [CrossRef] [PubMed]
- Badreddine, A. Préparation et Caractérisation d’extraits d’Argania Spinosa et d’huile d’argan et Evaluation de Leurs Effets Neuroprotecteurs In Vivo et In Vitro. Ph.D. Thesis, Université de Bourgogne, Dijon, France, Université Hassan 1er, Settat, Morocco, 2016. [Google Scholar]
- El Abbassi, A.; Khalid, N.; Zbakh, H.; Ahmad, A. Physicochemical characteristics, nutritional properties, and health benefits of argan oil: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1401–1414. [Google Scholar] [CrossRef] [PubMed]
- El Monfalouti, H.; Guillaume, D.; Denhez, C.; Charrouf, Z. Therapeutic potential of argan oil—A review. J. Pharm. Pharmacol. 2010, 62, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Drissi, A.; Girona, J.; Cherki, M.; Godàs, G.; Derouiche, A.; El Messal, M.; Saile, R.; Kettani, A.; Solà, R.; Masana, L.; et al. Evidence of hypolipemiant and antioxidant properties of argan oil derived from the argan tree (Argania spinosa). Clin. Nutr. 2004, 23, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Cherki, M.; Derouiche, A.; Drissi, A.; El Messal, M.; Bamou, Y.; Idrissi-Ouadghiri, A.; Khalil, A.; Adlouni, A. Consumption of argan oil may have an antiatherogenic effect by improving paraoxonase activities and antioxidant status: Intervention study in healthy men. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Derouiche, A.; Cherki, M.; Drissi, A.; Bamou, Y.; El Messal, M.; Idrissi-Oudghiri, A.; Lecerf, J.M.; Adlouni, A. Nutritional intervention study with argan oil in man: Effects on lipids and apolipoproteins. Ann. Nutr. Metab. 2005, 49, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Vique, C.; Marfil, R.; Giménez, R.; Martínez-Augustin, O. Bioactive compounds and nutritional significance of virgin argan oil--an edible oil with potential as a functional food. Nutr. Rev. 2012, 70, 266–279. [Google Scholar] [CrossRef] [PubMed]
- Boucetta, K.Q.; Charrouf, Z.; Derouiche, A.; Rahali, Y.; Bensouda, Y. Skin hydration in postmenopausal women: Argan oil benefit with oral and/or topical use. Prz. Menopauzalny 2014, 13, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Boucetta, K.Q.; Charrouf, Z.; Aguenaou, H.; Derouiche, A.; Bensouda, Y. The effect of dietary and/or cosmetic argan oil on postmenopausal skin elasticity. Clin. Interv. Aging 2015, 10, 339–349. [Google Scholar] [PubMed]
- Bahbiti, Y.; Ammouri, H.; Berkiks, I.; El Hessni, A.; Ouichou, A.; Nakache, R.; Chakit, M.; Bikjdaouene, L.; Mesfioui, A. Anticonvulsant effect of argan oil on pilocarpine model induced status epilepticus in wistar rats. Nutr. Neurosci. 2016, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Aydın, B. Effects of argan oil on the mitochondrial function, antioxidant system and the activity of NADPH-generating enzymes in acrylamide treated rat brain. Biomed. Pharmacother. 2017, 87, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Şekeroğlu, Z.A.; Aydın, B.; Şekeroğlu, V. Argan oil reduces oxidative stress, genetic damage and emperipolesis in rats treated with acrylamide. Biomed. Pharmacother. 2017, 94, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Khallouki, F.; Younos, C.; Soulimani, R.; Oster, T.; Charrouf, Z.; Spiegelhalder, B.; Bartsch, H.; Owen, R.W. Consumption of argan oil (Morocco) with its unique profile of fatty acids, tocopherols, squalene, sterols and phenolic compounds should confer valuable cancer chemopreventive effects. Eur. J. Cancer Prev. 2003, 12, 67–75. [Google Scholar] [CrossRef] [PubMed]
- López, L.C.; Cabrera-Vique, C.; Venegas, C.; García-Corzo, L.; Luna-Sánchez, M.; Acuña-Castroviejo, D.; Escames, G. Argan oil-contained antioxidants for human mitochondria. Nat. Prod. Commun. 2013, 8, 47–50. [Google Scholar] [PubMed]
- Trompier, D.; Vejux, A.; Zarrouk, A.; Gondcaille, C.; Geillon, F.; Nury, T.; Savary, S.; Lizard, G. Brain peroxisomes. Biochimie 2014, 98, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debbabi, M.; Nury, T.; Zarrouk, A.; Mekahli, N.; Bezine, M.; Sghaier, R.; Grégoire, S.; Martine, L.; Durand, P.; Camus, E.; et al. Protective effects of α-tocopherol, γ-tocopherol and oleic acid, three compounds of olive oils, and no effect of trolox, on 7-ketocholesterol-induced mitochondrial and peroxisomal dysfunction in microglial BV-2 cells. Int. J. Mol. Sci. 2016, 17, E1973. [Google Scholar] [CrossRef] [PubMed]
- Testa, G.; Staurenghi, E.; Zerbinati, C.; Gargiulo, S.; Iuliano, L.; Giaccone, G.; Fantò, F.; Poli, G.; Leonarduzzi, G.; Gamba, P. Changes in brain oxysterols at different stages of Alzheimer’s disease: Their involvement in neuroinflammation. Redox Biol. 2016, 10, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Leoni, V.; Lütjohann, D.; Masterman, T. Levels of 7-oxocholesterol in cerebrospinal fluid are more than one thousand times lower than reported in multiple sclerosis. J. Lipid Res. 2005, 46, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Boenzi, S.; Deodato, F.; Taurisano, R.; Goffredo, B.M.; Rizzo, C.; Dionisi-Vici, C. Evaluation of plasma cholestane-3β,5α,6β-triol and 7-ketocholesterol in inherited disorders related to cholesterol metabolism. J. Lipid Res. 2016, 57, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Nury, T.; Zarrouk, A.; Ragot, K.; Debbabi, M.; Riedinger, J.M.; Vejux, A.; Aubourg, P.; Lizard, G. 7-Ketocholesterol is increased in the plasma of X-ALD patients and induces peroxisomal modifications in microglial cells: Potential roles of 7-ketocholesterol in the pathophysiology of X-ALD. J. Steroid. Biochem. Mol. Biol. 2017, 169, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Nury, T. Caractérisation de la Neurotoxicité des 4- et 7-hydroxycholestérols: Impact sur la prolifération, le stress oxydant, l’apoptose et l’autophagie (notion d’oxyapoptophagie). Diplôme de l’Ecole Pratique des Hautes Etudes (EPHE). Master Grade, Dijon/Paris-Sorbonne, Paris, France, 2014. [Google Scholar]
- Wang, B.; Van Veldhoven, P.P.; Brees, C.; Rubio, N.; Nordgren, M.; Apanasets, O.; Kunze, M.; Baes, M.; Agostinis, P.; Fransen, M. Mitochondria are targets for peroxisome-derived oxidative stress in cultured mammalian cells. Free Radic. Biol. Med. 2013, 65, 882–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lismont, C.; Nordgren, M.; Van Veldhoven, P.P.; Fransen, M. Redox interplay between mitochondria and peroxisomes. Front. Cell Dev. Biol. 2015, 3, 35. [Google Scholar] [CrossRef] [PubMed]
- Sudo, R.; Sato, F.; Azechi, T.; Wachi, H. 7-Ketocholesterol-induced lysosomal dysfunction exacerbates vascular smooth muscle cell calcification via oxidative stress. Genes Cells 2015, 20, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Chakhchar, A.; Haworth, M.; El Modafar, C.; Lauteri, M.; Mattioni, C.; Wahbi, S.; Centritto, M. An assessment of genetic diversity and drought tolerance in argan tree (Argania spinosa) populations: Potential for the development of improved drought tolerance. Front Plant Sci. 2017, 8, 276. [Google Scholar] [CrossRef] [PubMed]
- Adlouni, A. L’huile d’argan, de la nutrition à la santé. Phytothérapie 2010, 8, 89–97. [Google Scholar] [CrossRef]
- Behrman, E.J.; Gopalan, V. Cholesterol and plants. J. Chem. Educ. 2005, 82, 1791–1793. [Google Scholar] [CrossRef]
- Sonawane, P.D.; Pollier, J.; Panda, S.; Szymanski, J.; Massalha, H.; Yona, M.; Unger, T.; Malitsky, S.; Arendt, P.; Pauwels, L.; et al. Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. Nat. Plants 2016, 3, 16205. [Google Scholar] [CrossRef] [PubMed]
- Vejux, A.; Malvitte, L.; Lizard, G. Side effects of oxysterols: Cytotoxicity, oxidation, inflammation, and phospholipidosis. Braz. J. Med. Biol. Res. 2008, 41, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Poli, G.; Biasi, F.; Leonarduzzi, G. Oxysterols in the pathogenesis of major chronic diseases. Redox Biol. 2013, 1, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Estrada, M.T.; Garcia-Llatas, G.; Lagarda, M.J. 7-Ketocholesterol as marker of cholesterol oxidation in model and food systems: When and how. Biochem. Biophys. Res. Commun. 2014, 446, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G.; Stevens, J.F. Vitamins C and E: Beneficial effects from a mechanistic perspective. Free Radic. Biol. Med. 2011, 51, 1000–1013. [Google Scholar] [CrossRef] [PubMed]
- Basli, A.; Soulet, S.; Chaher, N.; Mérillon, J.M.; Chibane, M.; Monti, J.P.; Richard, T. Wine polyphenols: Potential agents in neuroprotection. Oxid. Med. Cell. Longev. 2012, 2012, 805762. [Google Scholar] [CrossRef] [PubMed]
- Cilla, A.; Alegría, A.; Attanzio, A.; Garcia-Llatas, G.; Tesoriere, L.; Livrea, M.A. Dietary phytochemicals in the protection against oxysterol-induced damage. Chem. Phys. Lipids 2017, 207 Pt B, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Debbabi, M.; Zarrouk, A.; Bezine, M.; Meddeb, W.; Nury, T.; Badreddine, A.; Karym, E.M.; Sghaier, R.; Bretillon, L.; Guyot, S.; et al. Comparison of the effects of major fatty acids present in the Mediterranean diet (oleic acid, docosahexaenoic acid) and in hydrogenated oils (elaidic acid) on 7-ketocholesterol-induced oxiapoptophagy in microglial BV-2 cells. Chem. Phys. Lipids 2017, 207 Pt B, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Ragot, K.; Delmas, D.; Athias, A.; Nury, T.; Baarine, M.; Lizard, G. α-Tocopherol impairs 7-ketocholesterol-induced caspase-3-dependent apoptosis involving GSK-3 activation and Mcl-1 degradation in 158N murine oligodendrocytes. Chem. Phys. Lipids 2011, 164, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Lizard, G.; Deckert, V.; Dubrez, L.; Moisant, M.; Gambert, P.; Lagrost, L. Induction of apoptosis in endothelial cells treated with cholesterol oxides. Am. J. Pathol. 1996, 148, 1625–1638. [Google Scholar] [PubMed]
- Zahm, J.M.; Baconnais, S.; Monier, S.; Bonnet, N.; Bessède, G.; Gambert, P.; Puchelle, E.; Lizard, G. Chronology of cellular alterations during 7-ketocholesterol-induced cell death on A7R5 rat smooth muscle cells: Analysis by time lapse-video microscopy and conventional fluorescence microscopy. Cytometry A 2003, 52, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Zarrouk, A.; Nury, T.; Karym, E.M.; Vejux, A.; Sghaier, R.; Gondcaille, C.; Andreoletti, P.; Trompier, D.; Savary, S.; Cherkaoui-Malki, M.; et al. Attenuation of 7-ketocholesterol-induced overproduction of reactive oxygen species, apoptosis, and autophagy by dimethyl fumarate in 158N murine oligodendrocytes. J. Steroid. Biochem. Mol. Biol. 2017, 169, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Monier, S.; Samadi, M.; Prunet, C.; Denance, M.; Laubriet, A.; Athias, A.; Berthier, A.; Steinmetz, E.; Jürgens, G.; Nègre-Salvayre, A.; et al. Impairment of the cytotoxic and oxidative activities of 7beta-hydroxycholesterol and 7-ketocholesterol by esterification with oleate. Biochem. Biophys. Res. Commun. 2003, 303, 814–824. [Google Scholar] [CrossRef]
- Nury, T.; Zarrouk, A.; Mackrill, J.J.; Samadi, M.; Durand, P.; Riedinger, J.M.; Doria, M.; Vejux, A.; Limagne, E.; Delmas, D.; et al. Induction of oxiapoptophagy in 158N murine oligodendrocytes treated by 7-ketocholesterol-, 7β-hydroxycholesterol-, or 24(S)-hydroxycholesterol: Protective effects of α-tocopherol and docosahexaenoic acid (DHA; C22:6 n-3). Steroids 2015, 99 Pt B, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Baes, M.; Aubourg, P. Peroxisomes, myelination, and axonal integrity in the CNS. Neuroscientist 2009, 15, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.; Rice, C.; Hares, K.; Redondo, J.; Kemp, K.; Williams, M.; Brown, A.; Scolding, N.; Wilkins, A. Reductions in neuronal peroxisomes in multiple sclerosis grey matter. Mult. Scler. 2014, 20, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Jana, M.; Modi, K.; Gonzalez, F.J.; Sims, K.B.; Berry-Kravis, E.; Pahan, K. Activation of peroxisome proliferator-activated receptor α induces lysosomal biogenesis in brain cells: Implications for lysosomal storage disorders. J. Biol. Chem. 2015, 290, 10309–10324. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, M.A.; Yadav, S.; Gupta, R.K.; Waggoner, G.R.; Deloach, A.; Calingasan, N.Y.; Beal, M.F.; Kiaei, M. Preferential PPAR-α activation reduces neuroinflammation, and blocks neurodegeneration in vivo. Hum. Mol. Genet. 2016, 25, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Yadav, A.; Chaturvedi, R.K. Peroxisome proliferator-activated receptors (PPARs) as therapeutic target in neurodegenerative disorders. Biochem. Biophys. Res. Commun. 2017, 483, 1166–1177. [Google Scholar] [CrossRef] [PubMed]
- Ragot, K.; Mackrill, J.J.; Zarrouk, A.; Nury, T.; Aires, V.; Jacquin, A.; Athias, A.; Pais de Barros, J.P.; Vejux, A.; Riedinger, J.M.; et al. Absence of correlation between oxysterol accumulation in lipid raft microdomains, calcium increase, and apoptosis induction in 158N murine oligodendrocytes. Biochem. Pharmacol. 2013, 86, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Nury, T.; Zarrouk, A.; Vejux, A.; Doria, M.; Riedinger, J.M.; Delage-Mourroux, R.; Lizard, G. Induction of oxiapoptophagy, a mixed mode of cell death associated with oxidative stress, apoptosis and autophagy, on 7-ketocholesterol-treated 158N murine oligodendrocytes: Impairment by α-tocopherol. Biochem. Biophys. Res. Commun. 2014, 446, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Goldfinger, T.M. Beyond the French paradox: The impact of moderate beverage alcohol and wine consumption in the prevention of cardiovascular disease. Cardiol. Clin. 2003, 21, 449–457. [Google Scholar] [CrossRef]
- Ferrières, J. The French paradox: Lessons for other countries. Heart 2004, 90, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Lecour, S.; Lamont, K.T. Natural polyphenols and cardioprotection. Mini Rev. Med. Chem. 2011, 11, 1191–1199. [Google Scholar] [PubMed]
- Nadtochiy, S.M.; Redman, E.K. Mediterranean diet and cardioprotection: The role of nitrite; polyunsaturated fatty acids, and polyphenols. Nutrition 2011, 27, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Martínez-González, M.A.; Salas-Salvadó, J.; Estruch, R.; Corella, D.; Fitó, M.; Ros, E. Ppredimed investigators. Benefits of the mediterranean diet: Insights from the predimed study. Prog. Cardiovasc. Dis. 2015, 58, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Canhada, S.; Castro, K.; Perry, I.S.; Luft, V.C. Omega-3 fatty acids’ supplementation in Alzheimer’s disease: A systematic review. Nutr. Neurosci. 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Casamenti, F.; Stefani, M. Olive polyphenols: New promising agents to combat aging-associated neurodegeneration. Expert Rev. Neurother. 2017, 7, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Burg, V.K.; Grimm, H.S.; Rothhaar, T.L.; Grösgen, S.; Hundsdörfer, B.; Haupenthal, V.J.; Zimmer, V.C.; Mett, J.; Weingärtner, O.; Laufs, U.; et al. Plant sterols the better cholesterol in Alzheimer’s disease? A mechanistical study. J. Neurosci. 2013, 33, 16072–16087. [Google Scholar] [CrossRef] [PubMed]
- Shuang, R.; Rui, X.; Wenfang, L. Phytosterols and Dementia. Plant Foods Hum. Nutr. 2016, 71, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.O.; Mett, J.; Hartmann, T. The Impact of Vitamin E and Other Fat-Soluble Vitamins on Alzheimer’s Disease. Int. J. Mol. Sci. 2016, 17, E1785. [Google Scholar] [CrossRef] [PubMed]
- Lizard, G.; Filali-Zegzouti, Y.; Midaoui, A.E. Benefits of Argan Oil on Human Health-May 4–6 2017, Errachidia, Morocco. Int. J. Mol. Sci. 2017, 18, 1383. [Google Scholar] [CrossRef] [PubMed]
- Tritschler, H.J.; Packer, L.; Medori, R. Oxidative stress and mitochondrial dysfunction in neurodegeneration. Biochem. Mol. Biol. Int. 1994, 34, 169–181. [Google Scholar] [PubMed]
- Titorenko, V.I.; Terlecky, S.R. Peroxisome metabolism and cellular aging. Traffic 2011, 12, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Fransen, M.; Nordgren, M.; Wang, B.; Apanasets, O.; Van Veldhoven, P.P. Aging, age-related diseases and peroxisomes. Subcell. Biochem. 2013, 69, 45–65. [Google Scholar] [PubMed]
- Galea, E.; Launay, N.; Portero-Otin, M.; Ruiz, M.; Pamplona, R.; Aubourg, P.; Ferrer, I.; Pujol, A. Oxidative stress underlying axonal degeneration in adrenoleukodystrophy: A paradigm for multifactorial neurodegenerative diseases? Biochim. Biophys. Acta 2012, 1822, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Depreter, M.; Espeel, M.; Roels, F. Human peroxisomal disorders. Microsc. Res. Tech. 2003, 61, 203–223. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, V.K.; Jin, W.; Mochizuki, A.; Chitou, B.; Goodenowe, D.B. Metabolic dysfunctions in multiple sclerosis: Implications as to causation, early detection, and treatment, a case control study. BMC Neurol. 2015, 15, 154. [Google Scholar] [CrossRef] [PubMed]
- Kou, J.; Kovacs, G.G.; Höftberger, R.; Kulik, W.; Brodde, A.; Forss-Petter, S.; Hönigschnabl, S.; Gleiss, A.; Brügger, B.; Wanders, R.; et al. Peroxisomal alterations in Alzheimer’s disease. Acta Neuropathol. 2011, 122, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Lizard, G.; Rouaud, O.; Demarquoy, J.; Cherkaoui-Malki, M.; Iuliano, L. Potential roles of peroxisomes in Alzheimer’s disease and in dementia of the Alzheimer’s type. J. Alzheimers Dis. 2012, 29, 241–254. [Google Scholar] [PubMed]
- Zarrouk, A.; Riedinger, J.M.; Ahmed, S.H.; Hammami, S.; Chaabane, W.; Debbabi, M.; Ben Ammou, S.; Rouaud, O.; Frih, M.; Lizard, G.; et al. Fatty acid profiles in demented patients: Identification of hexacosanoic acid (C26:0) as a blood lipid biomarker of dementia. J. Alzheimers Dis. 2015, 44, 1349–1359. [Google Scholar] [PubMed]
- Loos, B.; Klionsky, D.J.; Wong, E. Augmenting brain metabolism to increase macro- and chaperone-mediated autophagy for decreasing neuronal proteotoxicity and aging. Prog. Neurobiol. 2017, 156, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Rimbach, G.; Minihane, A.M.; Majewicz, J.; Fischer, A.; Pallauf, J.; Virgli, F.; Weinberg, P.D. Regulation of cell signalling by vitamin E. Proc. Nutr. Soc. 2002, 61, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Grimsrud, P.A.; Xie, H.; Griffin, T.J.; Bernlohr, D.A. Oxidative stress and covalent modification of protein with bioactive aldehydes. J. Biol. Chem. 2008, 283, 21837–21841. [Google Scholar] [CrossRef] [PubMed]
- Guyot, E.; Coumoul, X.; Chassé, J.F.; Khallouki, F.; Savouret, J.F.; Poirot, M.; Barouki, R. Identification of a new stilbene-derived inducer of paraoxonase 1 and ligand of the Aryl hydrocarbon Receptor. Biochem. Pharmacol. 2012, 83, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Khallouki, F.; Breuer, A.; Merieme, E.; Ulrich, C.M.; Owen, R.W. Characterization and quantitation of the polyphenolic compounds detected in methanol extracts of Pistacia atlantica Desf. Fruits from the Guelmim region of Morocco. J. Pharm. Biomed. Anal. 2017, 134, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Abdelmohsen, K.; Abe, A.; Abedin, M.J.; Abeliovich, H.; Acevedo Arozena, A.; Adachi, H.; Adams, C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12, 1–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guina, T.; Deiana, M.; Calfapietra, S.; Cabboi, B.; Maina, M.; Tuberoso, C.I.; Leonarduzzi, G.; Gamba, P.; Gargiulo, S.; Testa, G.; et al. The role of p38 MAPK in the induction of intestinal inflammation by dietary oxysterols: Modulation by wine phenolics. Food Funct. 2015, 6, 1218. [Google Scholar] [CrossRef] [PubMed]
- Steffen, Y.; Wiswedel, I.; Peter, D.; Schewe, T.; Sies, H. Cytotoxicity of myeloperoxidase/nitrite-oxidized low-density lipoprotein toward endothelial cells is due to a high 7beta-hydroxycholesterol to 7-ketocholesterol ratio. Free Radic. Biol. Med. 2006, 41, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Dugas, B.; Charbonnier, S.; Baarine, M.; Ragot, K.; Delmas, D.; Ménétrier, F.; Lherminier, J.; Malvitte, L.; Khalfaoui, T.; Bron, A.; et al. Effects of oxysterols on cell viability, inflammatory cytokines, VEGF, and reactive oxygen species production on human retinal cells: Cytoprotective effects and prevention of VEGF secretion by resveratrol. Eur. J. Nutr. 2010, 49, 435–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesoriere, L.; Attanzio, A.; Allegra, M.; Gentile, C.; Livrea, M.A. Phytochemical indicaxanthin suppresses 7-ketocholesterol-induced THP-1 cell apoptosis by preventing cytosolic Ca2+ increase and oxidative stress. Br. J. Nutr. 2013, 110, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Vejux, A.; Guyot, S.; Montange, T.; Riedinger, J.M.; Kahn, E.; Lizard, G. Phospholipidosis and down-regulation of the PI3-K/PDK-1/Akt signalling pathway are vitamin E inhibitable events associated with 7-ketocholesterol-induced apoptosis. J. Nutr. Biochem. 2009, 20, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Said, T.; Tremblay-Mercier, J.; Berrougui, H.; Rat, P.; Khalil, A. Effects of vegetable oils on biochemical and biophysical properties of membrane retinal pigment epithelium cells. Can. J. Physiol. Pharmacol. 2013, 91, 812–817. [Google Scholar] [CrossRef] [PubMed]
- Altmeyer, A.; Jung, A.C.; Ignat, M.; Benzina, S.; Denis, J.M.; Gueulette, J.; Noël, G.; Mutter, D.; Bischoff, P. Pharmacological enhancement of autophagy induced in a hepatocellular carcinoma cell line by high-LET radiation. Anticancer Res. 2010, 30, 303–310. [Google Scholar] [PubMed]
- El Kharrassi, Y.; Samadi, M.; Lopez, T.; Nury, T.; El Kebbaj, R.; Andreoletti, P.; El Hajj, H.I.; Vamecq, J.; Moustaid, K.; Latruffe, N.; et al. Biological activities of Schottenol and Spinasterol, two natural phytosterols present in argan oil and in cactus pear seed oil, on murine miroglial BV2 cells. Biochem. Biophys. Res. Commun. 2014, 446, 798–804. [Google Scholar] [CrossRef] [PubMed]
- El Kebbaj, R.; El Kamouni, S.; El Hajj, H.I.; Andreoletti, P.; Gresti, J.; Latruffe, N.; El Kebbaj, S.; Vamecq, J.; Lizard, G.; Nasser, B.; et al. Modulation of peroxisomes abundance by argan oil and lipopolysaccharides in acyl-CoA oxidase 1-deficient fibroblasts. Health 2013, 5, 62–69. [Google Scholar] [CrossRef]
- El Kebbaj, R.; Andreoletti, P.; El Hajj, H.I.; El Kharrassi, Y.; Vamecq, J.; Mandard, S.; Ezzahra-Saih, F.; Latruffe, N.; El Kebbaj, S.; Lizard, G.; et al. Argan oil prevents down-regulation induced by endotoxin on liver fatty acid oxidation and gluconeogenesis and on peroxisome proliferator-activated receptor gamma coactivator-1α, (PGC-1α), peroxisome proliferator-activated receptor α (PPARα) and estrogen related receptor α (ERRα). Biochim. Open 2015, 1, 51–59. [Google Scholar]
- Malvitte, L.; Montange, T.; Vejux, A.; Joffre, C.; Bron, A.; Creuzot-Garcher, C.; Lizard, G. Activation of a caspase-3-independent mode of cell death associated with lysosomal destabilization in cultured human retinal pigment epithelial cells (ARPE-19) exposed to 7beta-hydroxycholesterol. Curr. Eye Res. 2008, 33, 769–781. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Zhu, H.; Zhang, W.; Okon, I.; Wang, Q.; Li, H.; Le, Y.Z.; Xie, Z. 7-Ketocholesterol induces autophagy in vascular smooth muscle cells through Nox4 and Atg4B. Am. J. Pathol. 2013, 183, 626–637. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Autophagy in the pathogenesis of disease. Cell 2009, 132, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Baell, J.; Walters, M.A. Chemistry: Chemical con artists foil drug discovery. Nature 2014, 513, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, J.L.; Chin, C.M. Pan-Assay interference compounds (PAINS): Warning signs in biochemical-pharmacological evaluations. Biochem. Pharmacol. 2015, 4, e137. [Google Scholar] [CrossRef]
- Jasial, S.; Hu, Y.; Bajorath, J. How Frequently Are Pan-Assay Interference Compounds Active? Large-Scale Analysis of Screening Data Reveals Diverse Activity Profiles, Low Global Hit Frequency, and Many Consistently Inactive Compounds. J. Med. Chem. 2017, 60, 3879–3886. [Google Scholar] [CrossRef] [PubMed]
- Vejux, A.; Lizard, G. Cytotoxic effects of oxysterols associated with human diseases: Induction of cell death (apoptosis and/or oncosis), oxidative and inflammatory activities, and phospholipidosis. Mol. Asp. Med. 2009, 30, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Capuzzi, S.J.; Muratov, E.N.; Tropsha, A. Phantom PAINS: Problems with the utility of alerts for Pan-Assay INterference CompoundS. J. Chem. Inf. Model. 2017, 57, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Vandenhaute, E.; Dehouck, L.; Boucau, M.C.; Sevin, E.; Uzbekov, R.; Tardivel, M.; Gosselet, F.; Fenart, L.; Cecchelli, R.; Dehouck, M.P. Modelling the neurovascular unit and the blood-brain barrier with the unique function of pericytes. Curr. Neurovasc. Res. 2011, 8, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Gosselet, F.; Saint-Pol, J.; Fenart, L. Effects of oxysterols on the blood-brain barrier: Implications for Alzheimer’s disease. Biochem. Biophys. Res. Commun. 2014, 446, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Baarine, M.; Ragot, K.; Genin, E.C.; El Hajj, H.; Trompier, D.; Andreoletti, P.; Ghandour, M.S.; Menetrier, F.; Cherkaoui-Malki, M.; Savary, S.; et al. Peroxisomal and mitochondrial status of two murine oligodendrocytic cell lines (158N, 158JP) potential models for the study of peroxisomal disorders associated with dysmyelination processes. J. Neurochem. 2009, 111, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Leoni, V.; Nury, T.; Vejux, A.; Zarrouk, A.; Caccia, C.; Debbabi, M.; Fromont, A.; Sghaier, R.; Moreau, T.; Lizard, G. Mitochondrial dysfunctions in 7-ketocholesterol-treated 158N oligodendrocytes without or with α-tocopherol: Impacts on the cellular profil of tricarboxylic cycle-associated organic acids, long chain saturated and unsaturated fatty acids, oxysterols, cholesterol and cholesterol precursors. J. Steroid. Biochem. Mol. Biol. 2017, 169, 96–110. [Google Scholar] [PubMed]
- Zarrouk, A.; Nury, T.; Samadi, M.; O’Callaghan, Y.; Hammami, M.; O’Brien, N.M.; Lizard, G.; Mackrill, J.J. Effects of cholesterol oxides on cell death induction and calcium increase in human neuronal cells (SK-N-BE) and evaluation of the protective effects of docosahexaenoic acid (DHA; C22:6 n-3). Steroids 2015, 99 Pt B, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Miguet, C.; Monier, S.; Bettaieb, A.; Athias, A.; Besséde, G.; Laubriet, A.; Lemaire, S.; Néel, D.; Gambert, P.; Lizard, G. Ceramide generation occurring during 7beta-hydroxycholesterol- and 7-ketocholesterol-induced apoptosis is caspase independent and is not required to trigger cell death. Cell Death Differ. 2001, 8, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Kahn, E.; Ménétrier, F.; Vejux, A.; Montange, T.; Dumas, D.; Riedinger, J.M.; Frouin, F.; Tourneur, Y.; Brau, F.; Stoltz, J.F.; et al. Flow cytometry and spectral imaging multiphoton microscopy analysis of CD36 expression with quantumdots 605 of untreated and 7-ketocholesterol-treated human monocytic cells. Anal. Quant. Cytol. Histol. 2006, 28, 316–330. [Google Scholar] [PubMed]
- Vurusaner, B.; Gamba, P.; Testa, G.; Gargiulo, S.; Biasi, F.; Zerbinati, C.; Iuliano, L.; Leonarduzzi, G.; Basaga, H.; Poli, G. Survival signalingelicited by 27-hydroxycholesterol through the combined modula-tion of cellular redox state and ERK/Akt phosphorylation. Free Radic. Biol. Med. 2014, 77, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Charrouf, Z. Valorisation des produits de l’arganier pour une gestion durable des zones arides du sud-ouest marocain. In Proceedings of the Actes du 4ème Colloque Produits naturels d’origine végétale, Ottawa, ON, Canada, 26–29 May 1998; Collin, G., et Garneau, F.X., Eds.; Université du Québec: Chicoutimi, QC, Canada, 1999; pp. 195–209. [Google Scholar]
- Moilanen, T.; Nikkari, T. The effect of storage on the fatty acid composition of human serum. Clin. Chim. Acta 1981, 114, 111–116. [Google Scholar] [CrossRef]
- Morrison, W.R.; Smith, L.M. Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride-methanol. J. Lipid Res. 1964, 5, 600–608. [Google Scholar] [PubMed]
- Lyan, B.; Azaïs-Braesco, V.; Cardinault, N.; Tyssandier, V.; Borel, P.; Alexandre-Gouabau, M.C.; Grolier, P. Simple method for clinical determination of 13 carotenoids in human plasma using an isocratic high-performance liquid chromatographic method. J. Chromatogr. B Biomed. Sci. Appl. 2001, 751, 297–303. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Khali, M.; Mahmoudi, N. Etude de l’extraction des composés phénoliques de différentes parties de la fleur d’artichaut (Cynara scolymus L.). Nat. Technol. B Sci. Agron. Biol. 2013, 9, 35–40. [Google Scholar]
- Rossi, R.; Pastorelli, G.; Corino, C. Application of KRL test to assess total antioxidant activity in pigs: Sensitivity to dietary antioxidants. Res. Vet. Sci. 2013, 94, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Oyaizu, M. Studies on products of browning reaction: Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. 1986, 44, 307. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as ameasure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorunetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Colin, D.; Limagne, E.; Jeanningros, S.; Jacquel, A.; Lizard, G.; Athias, A.; Gambert, P.; Hichami, A.; Latruffe, N.; Solary, E.; et al. Endocytosis of resveratrol via lipid rafts and activation of downstream signaling pathways in cancer cells. Cancer Prev. Res. (Phila.) 2011, 4, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Lizard, G.; Gueldry, S.; Deckert, V.; Gambert, P.; Lagrost, L. Evaluation of the cytotoxic effects of some oxysterols and of cholesterol on endothelial cell growth: Methodological aspects. Pathol. Biol. (Paris) 1997, 45, 281–290. [Google Scholar] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Olsson, M.; Rundquist, I.; Brunk, U. Flow cytofluorometry of lysosomal acridine orange uptake by living cultured cells. Acta Path. Microbiol. Immunol. Scand. (Sect. A) 1987, 95, 159–165. [Google Scholar]
- Olsson, G.M.; Rungby, J.; Rundquist, I.; Brunk, U.T. Evaluation of lysosomal stability in living cultured macrophages by cytofluorometry. Effect of silver lactate and hypotonic conditions. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1989, 56, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.M.; Li, W.; Brunk, U.T.; Dalen, H.; Chang, Y.H.; Sevanian, A. Lysosomal destabilization during macrophage damage induced by cholesterol oxidation products. Free Radic. Biol. Med. 2000, 28, 208–218. [Google Scholar] [CrossRef]
- Pierzyńska-Mach, A.; Janowski, P.A.; Dobrucki, J.W. Evaluation of acridine orange, LysoTracker Red, and quinacrine as fluorescent probes for long-term tracking of acidic vesicles. Cytometry A 2014, 85, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Rothe, G.; Valet, G. Flow cytometric analysis of respiratory burst activity in phagocytes with hydroethidine and 2′,7′-dichlorofluorescin. J. Leukoc. Biol. 1990, 47, 440–448. [Google Scholar] [PubMed]
- Zarrouk, A.; Vejux, A.; Nury, T.; El Hajj, H.I.; Haddad, M.; Cherkaoui-Malki, M.; Riedinger, J.M.; Hammami, M.; Lizard, G. Induction of mitochondrial changes associated with oxidative stress on very long chain fatty acids (C22:0, C24:0, or C26:0)-treated human neuronal cells (SK-NB-E). Oxid. Med. Cell. Longev. 2012, 2012, 623257. [Google Scholar] [CrossRef] [PubMed]
- Lizard, G.; Fournel, S.; Genestier, L.; Dhedin, N.; Chaput, C.; Flacher, M.; Mutin, M.; Panaye, G.; Revillard, J.P. Kinetics of plasma membrane and mitochondrial alterations in cells undergoing apoptosis. Cytometry 1995, 21, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Zerbinati, C.; Iuliano, L. Cholesterol and related sterols autoxidation. Free Radic. Biol. Med. 2017, 111, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Badreddine, A.; El Mostafa, K.; Zarrouk, A.; Nury, T.; El Kharrassi, Y.; Nasser, B.; Cherkaoui Malki, M.; Lizard, G.; Samadi, M. An expeditious synthesis of spinasterol and schottenol, two phytosterols present in argan oil and in cactus pear seed oil, and evaluation of their biological activities on cells of the central nervous system. Steroids 2015, 99, 119–124. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids (mg/g of Total Lipids) | Dietary Argan Oils | Extra Virgin Olive Oil | |
|---|---|---|---|
| Morocco (Agadir) | Morocco (Berkane) | Tunisia (Mahdia) | |
| C12:0 | 0.00 | 0.00 | 0.00 |
| C14:0 | 1.47 ± 0.00 | 1.47 ± 0.00 | 0.00 |
| C15:0 | 0.52 ± 0.01 | 0.59 ± 0.02 | 0.00 |
| C16:0 | 119.00 ± 0.37 | 132.00 ± 4.97 | 198.00 ± 6.51 |
| C16:1 n-7 | 1.04 ± 0.03 | 1.06 ± 0.05 | 29.00 ± 1.00 |
| C16:1 n-9 | 0.00 | 0.00 | 1.00 ± 0.00 |
| C17:0 | 0.84 ± 0.04 | 0.97 ± 0.02 | 0.00 |
| C18:0 | 62.60 ± 1.27 | 65.00 ± 2.70 | 24.70 ± 1.53 |
| C18:1 trans | 10.90 ± 2.70 | 10.70 ± 0.40 | 10.00 ± 2.00 |
| C18:1n-9 (oleic acid) | 443.00 ± 0.51 | 464.00 ± 18.40 | 471.00 ± 15.00 |
| C18:1 n-7 | 3.86 ± 0.075 | 4.01 ± 0.01 | 38.00 ± 100.00 |
| C18:2 n-6 cis trans | 0.00 | 0.00 | 0.00 |
| C18:2 n-6 trans cis | 0.00 | 0.00 | 0.00 |
| C18:2 n-6 (linoleic acid) | 332.00 ± 5.72 | 324.00 ± 9.21 | 190 ± 3.51 |
| C20:0 | 3.23 ± 0.10 | 3.66 ± 0.09 | 4.00 ± 0.00 |
| C20:1 n-9 | 2.95 ± 0.01 | 3.38 ± 0.06 | 2.00 ± 0.00 |
| C18:3 n-3 | 1.16 ± 0.04 | 1.14 ± 0.01 | 7.33 ± 0.58 |
| C20:2 n-6 | 0.00 | 0.00 | 0.00 |
| C22:0 | 1.11 ± 0.00 | 1.34 ± 0.02 | 1.00 ± 0.00 |
| C22:1 n-9 | 0.00 | 0.00 | 0.00 |
| C24:0 | 0.53 ± 0.02 | 0.00 | 0.00 |
| C24:1 n-9 | 0.00 | 0.00 | 0.00 |
| Conjugated C18:3 | 0.00 | 0.00 | 0.00 |
| Sterol Contents (mg/kg of Oil) | Dietary Argan Oils | Extra Virgin Olive Oil | |
|---|---|---|---|
| Morocco (Agadir) | Morocco (Berkane) | Tunisia (Madhia) | |
| Cholesterol | ND | ND | ND |
| Brassicasterol | ND | ND | ND |
| 24-Methylenecholesterol | ND | ND | 10.90 ±1.05 |
| Campesterol | ND | ND | 75.60 ± 7.14 |
| Campestanol | 16.40 ± 6.76 | 15.90 ± 7.11 | 11.30 ± 0.6.1 |
| Stigmasterol | ND | ND | 15.20 ± 0.31 |
| ∆7-Stigmasterol | 48.20 ± 18.70 | 46.60 ± 9.31 | ND |
| ∆7-Campesterol | 37.50 ± 12.70 | 35.30 ±28.00 | ND |
| Spinasterol | 64.40 ± 249.00 | 567.00 ±74.60 | ND |
| Clerosterol | ND | ND | 27.40 ± 0.61 |
| β-Sitosterol | ND | ND | 1700.00 ± 32.50 |
| ∆5-Avenasterol | ND | ND | 203.00 ± 2.75 |
| β-amyrine | 211.00 ± 130.00 | 178.00 ± 13.00 | 20.10 ± 0.40 |
| Fucosterol | ND | ND | 20.20 ± 1.40 |
| Graminasterol | ND | ND | 29.00 ± 20.00 |
| Schottenol | 849.00 ± 402.00 | 62.50 ± 96.40 | ND |
| Cycloartenol | 239.00 ± 51.40 | 218.00 ± 46.80 | 272.00 ± 7.69 |
| ∆7-avenasterol | 85.80 ± 47.10 | 57.8 ± 3.00 | 12.30 ± 1.35 |
| 24-Methylene cycloartenol | 51.50 ± 27.00 | 44.50 ±22.30 | 333.30 ± 69.00 |
| Lupeol | 15.60 ± 7.13 | 22.70 ± 5.77 | ND |
| Citrostadienol | 132.00 ± 61.60 | 70.30 ± 14.00 | 202.00 ± 7.44 |
| Unkown | 133.00 ± 42.50 | 135.00 ± 204.00 | 29.30 ± 4.45 |
| Tocopherols (mg/kg of Oil) | Dietary Argan Oils | Extra Virgin Olive Oil | |
|---|---|---|---|
| Morocco (Agadir) | Morocco (Berkane) | Tunisia (Mahdia) | |
| α-tocopherol | 429.00 ± 7.00 (995.35 ± 16.24 µM) | 12.00 ± 1.00 (27.84 ± 2.32 µM) | 112.00 ± 3.00 (259.86 ± 6.96 µM) |
| γ-tocopherol | 12.00 ± 1.00 (28.77 ± 2.39 µM) | 355.00 ± 19.00 (851.31 ± 45.56 µM) | 9.00± 2.00 (21.58 ± 4.79 µM) |
| δ-tocopherol | 12.00 ± 1.00 (29.77 ± 2.48 µM) | 18.00 ± 2.00 (44.66 ± 4.96 µM) | 0.00 (0.00 µM) |
| Ratio [(α-tocopherol)/(γ-tocopherol)] | 35.75 | 0.03 | 12.44 |
| Polyphenols | Dietary Argan Oils | Extra Virgin Olive Oil | |
|---|---|---|---|
| Morocco (Agadir) | Morocco (Berkane) | Tunisia (Mahdia) | |
| Homovanillic acid | ND | ND | ND |
| Vanillin | ND | ND | ND |
| p-Coumaric acid | ND | ND | 0.3 |
| Quercetin-3β-glucoside | ND | ND | ND |
| Quercetin | ND | ND | ND |
| Apigenin | ND | ND | ND |
| 2.6-dihydroxybenzoic acid | ND | ND | ND |
| Chlorogenic acid | ND | ND | ND |
| Ferrulic acid | ND | ND | 0.11 |
| Thymoquinone | ND | ND | ND |
| Hydroxytyrosol | ND | ND | 0.65 |
| Tyrosol | 0.07 | 0.06 | 1.22 |
| Oleuropein | ND | ND | 0.39 |
| Luteoline | ND | ND | 0.39 |
| Protocatechic acid | ND | 0.17 | ND |
| Sum of identified pics | 0.07 | 0.18 | 2.78 |
| Sum of 280 nm pics | 0.54 | 0.75 | 4.62 |
| Compounds | Antioxidant Activity (Trolox Equivalent) | |
|---|---|---|
| KRL | FRAP | |
| α-tocopherol | 0.96 ± 0.01 * | 0.86 ± 0.03 * |
| Argan oil (Agadir) | 6372 ± 318 # | 7463 ± 373 # |
| Argan oil (Berkane) | 7524 ± 376 # | 6372 ± 318 # |
| Extra virgin olive oil (Madhia, Tunisia) | 7480 ± 374 # | 7524 ± 376 # |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badreddine, A.; Zarrouk, A.; Karym, E.M.; Debbabi, M.; Nury, T.; Meddeb, W.; Sghaier, R.; Bezine, M.; Vejux, A.; Martine, L.; et al. Argan Oil-Mediated Attenuation of Organelle Dysfunction, Oxidative Stress and Cell Death Induced by 7-Ketocholesterol in Murine Oligodendrocytes 158N. Int. J. Mol. Sci. 2017, 18, 2220. https://doi.org/10.3390/ijms18102220
Badreddine A, Zarrouk A, Karym EM, Debbabi M, Nury T, Meddeb W, Sghaier R, Bezine M, Vejux A, Martine L, et al. Argan Oil-Mediated Attenuation of Organelle Dysfunction, Oxidative Stress and Cell Death Induced by 7-Ketocholesterol in Murine Oligodendrocytes 158N. International Journal of Molecular Sciences. 2017; 18(10):2220. https://doi.org/10.3390/ijms18102220
Chicago/Turabian StyleBadreddine, Asmaa, Amira Zarrouk, El Mostafa Karym, Meryam Debbabi, Thomas Nury, Wiem Meddeb, Randa Sghaier, Maryem Bezine, Anne Vejux, Lucy Martine, and et al. 2017. "Argan Oil-Mediated Attenuation of Organelle Dysfunction, Oxidative Stress and Cell Death Induced by 7-Ketocholesterol in Murine Oligodendrocytes 158N" International Journal of Molecular Sciences 18, no. 10: 2220. https://doi.org/10.3390/ijms18102220
APA StyleBadreddine, A., Zarrouk, A., Karym, E. M., Debbabi, M., Nury, T., Meddeb, W., Sghaier, R., Bezine, M., Vejux, A., Martine, L., Grégoire, S., Bretillon, L., Prost-Camus, E., Durand, P., Prost, M., Moreau, T., Cherkaoui-Malki, M., Nasser, B., & Lizard, G. (2017). Argan Oil-Mediated Attenuation of Organelle Dysfunction, Oxidative Stress and Cell Death Induced by 7-Ketocholesterol in Murine Oligodendrocytes 158N. International Journal of Molecular Sciences, 18(10), 2220. https://doi.org/10.3390/ijms18102220