Effect of Intranasally Delivered rh-VEGF165 on Angiogenesis Following Cerebral Hypoxia-Ischemia in the Cerebral Cortex of Newborn Piglets

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
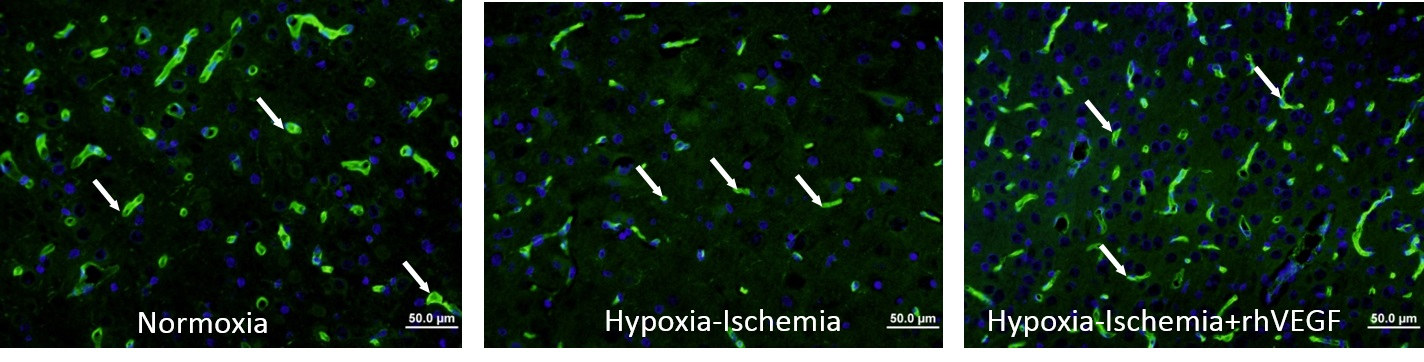
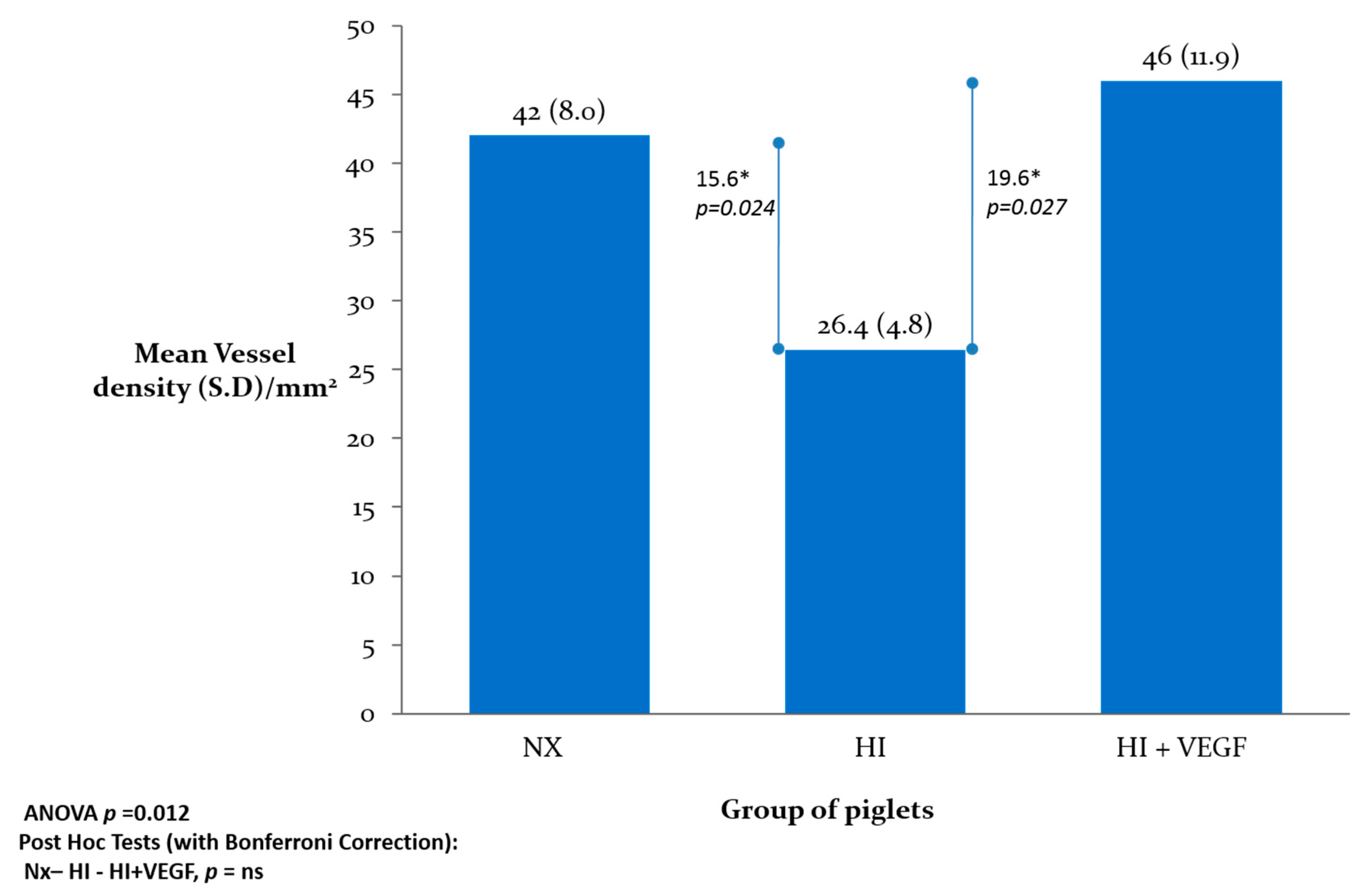
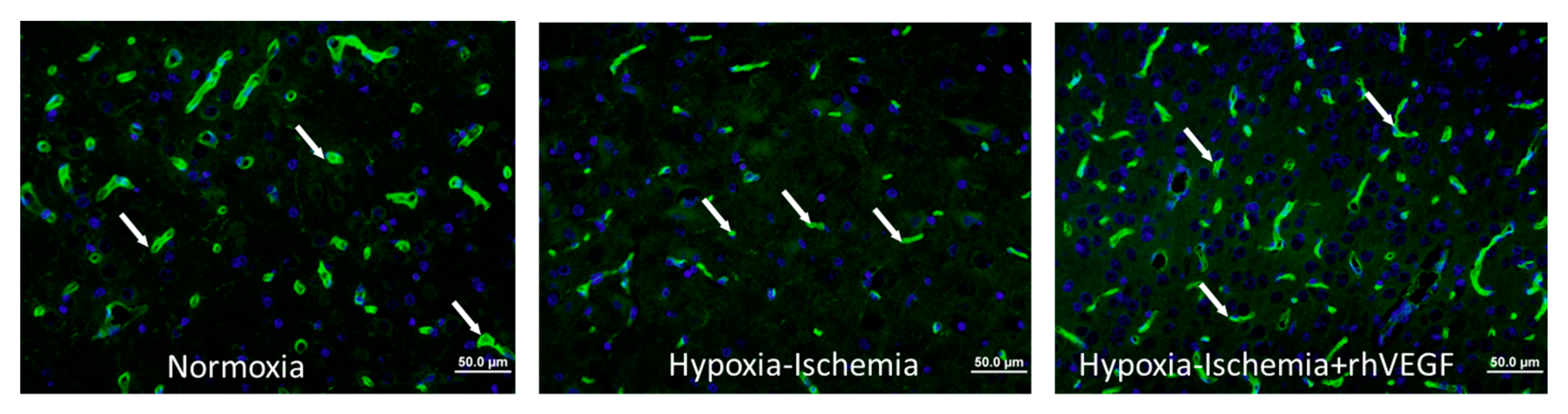
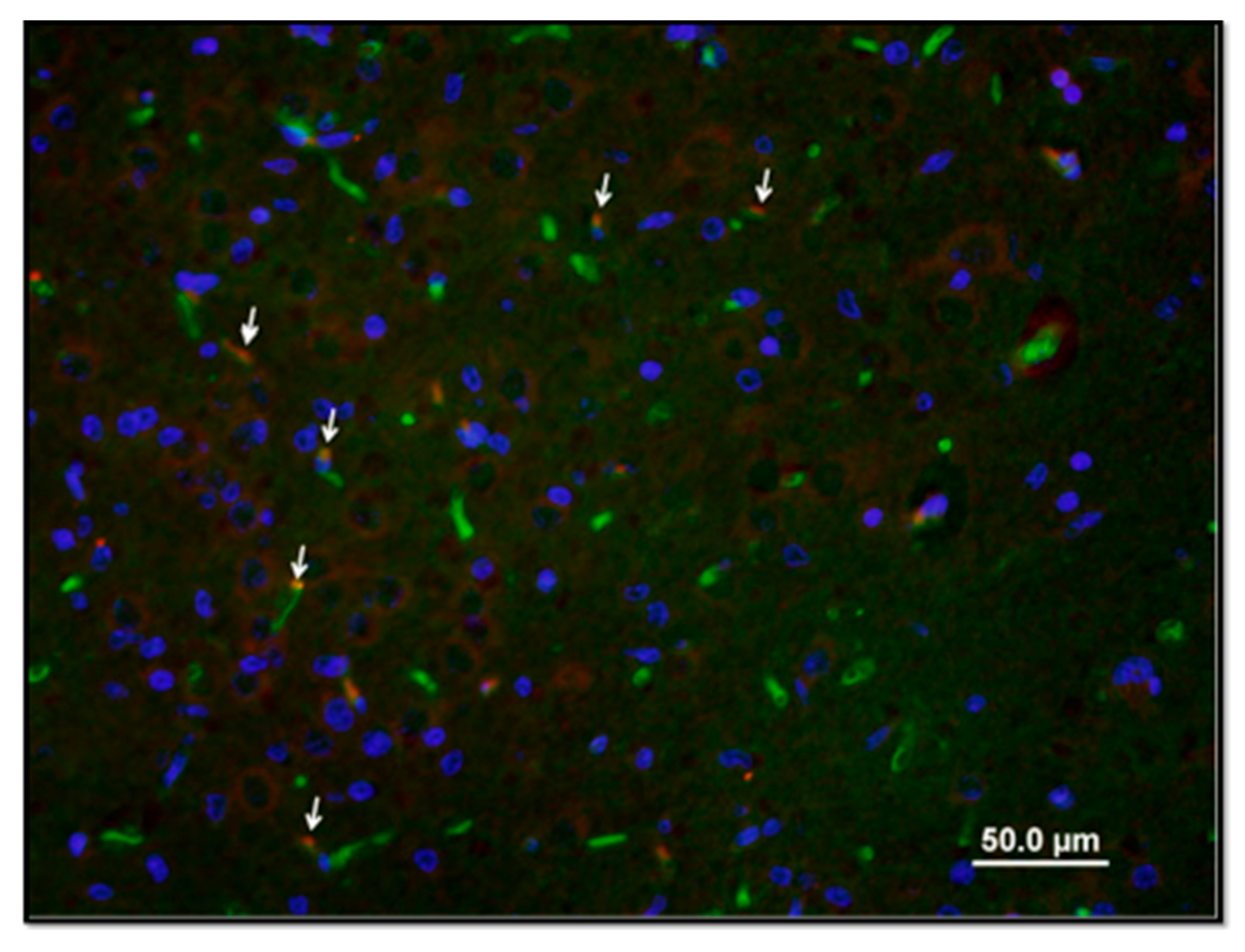
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Procedures
4.2. Induction of Cerebral Hypoxia–Ischemia
4.3. Intranasal Administration of Recombinant Human VEGF165
4.4. Immunohistochemistry
4.5. Vascular Density Assessment
4.6. Statistical Analysis
4.7. Study Limitations
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lawn, J.E.; Cousens, S.; Zupan, J. Lancet Neonatal Survival Steering Team. 4 million neonatal deaths: When? Where? Why? Lancet 2005, 365, 891–900. [Google Scholar] [CrossRef]
- Vannucci, R.C. Hypoxic-ischemic encephalopathy. Am. J. Perinatol. 2000, 17, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.D.; Brocklehurst, P.; Gunn, A.J.; Halliday, H.; Juszczak, E.; Levene, M.; Strohm, B.; Thoresen, M.; Whitelaw, A.; Azzopardi, D. Neurological outcomes at 18 months of age after moderate hypothermia for perinatal hypoxic ischaemic encephalopathy: Synthesis and meta-analysis of trial data. BMJ 2010, 340, c363. [Google Scholar] [CrossRef] [PubMed]
- Shankaran, S.; Pappas, A.; McDonald, S.A.; Vohr, B.R.; Hintz, S.R.; Yolton, K.; Gustafson, K.E.; Leach, T.M.; Green, C.; Bara, R.; et al. Childhood outcomes after hypothermia for neonatal encephalopathy. N. Engl. J. Med. 2012, 366, 2085–2092. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.L.; Eicher, D.J.; Katikaneni, L.D.; Barbosa, E.; Holden, K.R. The use of hypothermia: A role in the treatment of neonatal asphyxia? Pediatr. Neurol. 1999, 21, 429–443. [Google Scholar] [CrossRef]
- Azzopardi, D.; Strohm, B.; Marlow, N.; Brocklehurst, P.; Deierl, A.; Eddama, O.; Goodwin, J.; Halliday, H.L.; Juszczak, E.; Kapellou, O.; et al. Effects of hypothermia for perinatal asphyxia on childhood outcomes. N. Engl. J. Med. 2014, 371, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Kratimenos, P.; Koutroulis, I.; Marconi, D.; Syriopoulou, V.; Delivoria-Papadopoulos, M.; Chrousos, G.P.; Theocharis, S. Multi-targeted molecular therapeutic approach in aggressive neuroblastoma: The effect of Focal Adhesion Kinase-Src-Paxillin system. Expert Opin. Ther. Targets 2014, 18, 1395–1406. [Google Scholar] [PubMed]
- Kratimenos, P.; Koutroulis, I.; Syriopoulou, V.; Michailidi, C.; Delivoria-Papadopoulos, M.; Klijanienko, J.; Theocharis, S. FAK-Src-PAX System Expression and Disease Outcome in Human Neuroblastoma. Pediatr. Hematol. Oncol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Kratimenos, P.; Koutroulis, I.; Jain, A.; Malaeb, S.; Delivoria-Papadopoulos, M. Effect of Concurrent Src Kinase Inhibition with Short Duration Hypothermia on Ca2+/Calmodulin kinase IV (CaM Kinase IV) Activity and Neuropathology after Hypoxia-Ischemia in the Newborn Swine Brain. Neonatology 2017, 113, 37–43. [Google Scholar] [PubMed]
- Fischer, S.; Clauss, M.; Wiesnet, M.; Renz, D.; Schaper, W.; Karliczek, G.F. Hypoxia induces permeability in brain microvessel endothelial cells via VEGF and NO. Am. J. Physiol. 1999, 276, C812–C820. [Google Scholar] [PubMed]
- Xu, F.; Severinghaus, J.W. Rat brain VEGF expression in alveolar hypoxia: Possible role in high-altitude cerebral edema. J. Appl. Physiol. 1998, 85, 53–57. [Google Scholar] [PubMed]
- Jin, K.L.; Mao, X.O.; Greenberg, D.A. Vascular endothelial growth factor rescues HN33 neural cells from death induced by serum withdrawal. J. Mol. Neurosci. 2000, 14, 197–203. [Google Scholar] [CrossRef]
- Sun, Y.; Jin, K.; Xie, L.; Childs, J.; Mao, X.O.; Logvinova, A.; Greenberg, D.A. VEGF-induced neuroprotection, neurogenesis, and angiogenesis after focal cerebral ischemia. J. Clin. Investig. 2003, 111, 1843–1851. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Zhang, L.; Jiang, Q.; Zhang, R.; Davies, K.; Powers, C.; Bruggen, N.; Chopp, M. VEGF enhances angiogenesis and promotes blood-brain barrier leakage in the ischemic brain. J. Clin. Investig. 2000, 106, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Schoch, H.J.; Fischer, S.; Marti, H.H. Hypoxia-induced vascular endothelial growth factor expression causes vascular leakage in the brain. Brain 2002, 125, 2549–2557. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.; Jiang, X.; Sheldon, R.A.; Fox, C.K.; Hamrick, S.E.; Vexler, Z.S.; Ferriero, D.M. Regulation of hypoxia-inducible factor 1alpha and induction of vascular endothelial growth factor in a rat neonatal stroke model. Neurobiol. Dis. 2003, 14, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Bills, V.L.; Hamdollah-Zadeh, M.; Soothill, P.W.; Harper, S.J.; Bates, D.O. The role of VEGF-A165b in trophoblast survival. BMC Pregnancy Childbirth 2014, 14, 278. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.W.; Sandlund, J.; Han, C.H.; Hameed, M.Q.; Connors, S.; Klagsbrun, M.; Madsen, J.R.; Irwin, N. VEGF, which is elevated in the CSF of patients with hydrocephalus, causes ventriculomegaly and ependymal changes in rats. Exp. Neurol. 2013, 247, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Yeh, W.L.; Lu, D.Y.; Lin, C.J.; Liou, H.C.; Fu, W.M. Inhibition of hypoxia-induced increase of blood-brain barrier permeability by YC-1 through the antagonism of HIF-1alpha accumulation and VEGF expression. Mol. Pharmacol. 2007, 72, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.R.; Zhang, S.S.; Yang, Y.J.; Yin, F.; Wang, X.; Zhong, L.; Yu, X.H. Adenoviral vector-mediated transduction of VEGF improves neural functional recovery after hypoxia-ischemic brain damage in neonatal rats. Brain Res. Bull. 2010, 81, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Ara, J.; De Montpellier, S. Hypoxic-preconditioning enhances the regenerative capacity of neural stem/progenitors in subventricular zone of newborn piglet brain. Stem Cell Res. 2013, 11, 669–686. [Google Scholar] [CrossRef] [PubMed]
- Ara, J.; Fekete, S.; Frank, M.; Golden, J.A.; Pleasure, D.; Valencia, I. Hypoxic-preconditioning induces neuroprotection against hypoxia-ischemia in newborn piglet brain. Neurobiol. Dis. 2011, 43, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Wobben, M.; Marti, H.H.; Renz, D.; Schaper, W. Hypoxia-induced hyperpermeability in brain microvessel endothelial cells involves VEGF-mediated changes in the expression of zonula occludens-1. Microvasc. Res. 2002, 63, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.M.; Moore, L.; Ren, Z.; Simonishvili, S.; Levison, S.W. Vascular endothelial growth factors A and C are induced in the SVZ following neonatal hypoxia-ischemia and exert different effects on neonatal glial progenitors. Transl. Stroke Res. 2013, 4, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Thoresen, M.; Haaland, K.; Loberg, E.M.; Whitelaw, A.; Apricena, F.; Hanko, E.; Steen, P.A. A piglet survival model of posthypoxic encephalopathy. Pediatr. Res. 1996, 40, 738–748. [Google Scholar] [CrossRef] [PubMed]
- DiGiacomo, J.E.; Pane, C.R.; Gwiazdowski, S.; Mishra, O.P.; Delivoria-Papadopoulos, M. Effect of graded hypoxia on brain cell membrane injury in newborn piglets. Biol. Neonate 1992, 61, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Odden, J.P.; Stiris, T.; Hansen, T.W.; Bratlid, D. Cerebral blood flow during experimental hypoxaemia and ischaemia in the newborn piglet. Acta Paediatr. Scand. Suppl. 1989, 360, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J.; Brambrink, A.M.; Lehmann, C.; Portera-Cailliau, C.; Koehler, R.; Rothstein, J.; Traystman, R.J. Hypoxia-ischemia causes abnormalities in glutamate transporters and death of astroglia and neurons in newborn striatum. Ann. Neurol. 1997, 42, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J.; Brambrink, A.; Koehler, R.C.; Traystman, R.J. Primary sensory and forebrain motor systems in the newborn brain are preferentially damaged by hypoxia-ischemia. J. Comp. Neurol. 1997, 377, 262–285. [Google Scholar] [CrossRef]
- Brambrink, A.M.; Martin, L.J.; Hanley, D.F.; Becker, K.J.; Koehler, R.C.; Traystman, R.J. Effects of the AMPA receptor antagonist NBQX on outcome of newborn pigs after asphyxic cardiac arrest. J. Cereb. Blood Flow Metab. 1999, 19, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Guerguerian, A.M.; Brambrink, A.M.; Traystman, R.J.; Huganir, R.L.; Martin, L.J. Altered expression and phosphorylation of N-methyl-D-aspartate receptors in piglet striatum after hypoxia-ischemia. Brain Res. Mol. Brain Res. 2002, 104, 66–80. [Google Scholar] [CrossRef]
- Haaland, K.; Loberg, E.M.; Steen, P.A.; Thoresen, M. Posthypoxic hypothermia in newborn piglets. Pediatr. Res. 1997, 41, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Ezzati, M.; Kawano, G.; Rocha-Ferreira, E.; Alonso-Alconada, D.; Hassell, J.K.; Broad, K.D.; Fierens, I.; Fleiss, B.; Bainbridge, A.; Price, D.L.; et al. Dexmedetomidine Combined with Therapeutic Hypothermia Is Associated with Cardiovascular Instability and Neurotoxicity in a Piglet Model of Perinatal Asphyxia. Dev. Neurosci. 2017, 39, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Groenendaal, F.; de Graaf, R.A.; van Vliet, G.; Nicolay, K. Effects of hypoxia-ischemia and inhibition of nitric oxide synthase on cerebral energy metabolism in newborn piglets. Pediatr. Res. 1999, 45, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Mehmet, H.; Yue, X.; Penrice, J.; Cady, E.; Wyatt, J.C.; Sarraf, C.; Squier, M.; Edwards, A.D. Relation of impaired energy metabolism to apoptosis and necrosis following transient cerebral hypoxia-ischaemia. Cell Death Differ. 1998, 5, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Mehmet, H.; Yue, X.; Squier, M.V.; Lorek, A.; Cady, E.; Penrice, J.; Sarraf, C.; Wylezinska, M.; Kirkbride, V.; Cooper, C.; et al. Increased apoptosis in the cingulate sulcus of newborn piglets following transient hypoxia-ischaemia is related to the degree of high energy phosphate depletion during the insult. Neurosci. Lett. 1994, 181, 121–125. [Google Scholar] [CrossRef]
- Yue, X.; Mehmet, H.; Penrice, J.; Cooper, C.; Cady, E.; Wyatt, J.S.; Reynolds, E.O.; Edwards, A.D.; Squier, M.V. Apoptosis and necrosis in the newborn piglet brain following transient cerebral hypoxia-ischaemia. Neuropathol. Appl. Neurobiol. 1997, 23, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, S.T.; Foster, K.A.; O’Driscoll, S.M.; Healy, G.N.; Lingwood, B.E.; Burke, C.; Colditz, P.B. Hypoxic/Ischemic models in newborn piglet: Comparison of constant FiO2 versus variable FiO2 delivery. Brain Res. 2006, 1100, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.N.; Al Ahmad, A.; Lee, B.; Parham, C.; Auckland, L.; Fertala, A.; Kahle, M.; Shaw, C.S.; Roberts, J.; Bix, G.J. Perlecan Domain V induces VEGf secretion in brain endothelial cells through integrin alpha5beta1 and ERK-dependent signaling pathways. PLoS ONE 2012, 7, e45257. [Google Scholar] [CrossRef] [PubMed]
- Saraswat, D.; Nehra, S.; Chaudhary, K.; Cvs, S.P. Novel vascular endothelial growth factor blocker improves cellular viability and reduces hypobaric hypoxia-induced vascular leakage and oedema in rat brain. Clin. Exp. Pharmacol. Physiol. 2015, 42, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Trollmann, R.; Richter, M.; Jung, S.; Walkinshaw, G.; Brackmann, F. Pharmacologic stabilization of hypoxia-inducible transcription factors protects developing mouse brain from hypoxia-induced apoptotic cell death. Neuroscience 2014, 278, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Baburamani, A.A.; Castillo-Melendez, M.; Walker, D.W. VEGF expression and microvascular responses to severe transient hypoxia in the fetal sheep brain. Pediatr. Res. 2013, 73, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Benderro, G.F.; Sun, X.; Kuang, Y.; Lamanna, J.C. Decreased VEGF expression and microvascular density, but increased HIF-1 and 2alpha accumulation and EPO expression in chronic moderate hyperoxia in the mouse brain. Brain Res. 2012, 1471, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.R.; Zhang, S.S.; Yin, F.; Tang, J.L.; Yang, Y.J.; Wang, X.; Zhong, L. Neuroprotection of VEGF-expression neural stem cells in neonatal cerebral palsy rats. Behav. Brain Res. 2012, 230, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Illum, L. Is nose-to-brain transport of drugs in man a reality? J. Pharm. Pharmacol. 2004, 56, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Hao, J.; Fei, X.; Wang, X.; Hou, Y.; Deng, C. Isoflurane inhibits occludin expression via up-regulation of hypoxia-inducible factor 1alpha. Brain Res. 2014, 1562, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Thelin, E.P.; Frostell, A.; Mulder, J.; Mitsios, N.; Damberg, P.; Aski, S.N.; Risling, M.; Svensson, M.; Morganti-Kossmann, M.C.; Bellander, B.M. Lesion Size Is Exacerbated in Hypoxic Rats Whereas Hypoxia-Inducible Factor-1 Alpha and Vascular Endothelial Growth Factor Increase in Injured Normoxic Rats: A Prospective Cohort Study of Secondary Hypoxia in Focal Traumatic Brain Injury. Front. Neurol. 2016, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yao, Y.; Chen, T.; Zhang, T. VEGF ameliorates cognitive impairment in in vivo and in vitro ischemia via improving neuronal viability and function. Neuromol. Med. 2014, 16, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Scafidi, J.; Hammond, T.R.; Scafidi, S.; Ritter, J.; Jablonska, B.; Roncal, M.; Szigeti-Buck, K.; Coman, D.; Huang, Y.; McCarter, R.J., Jr.; et al. Intranasal epidermal growth factor treatment rescues neonatal brain injury. Nature 2014, 506, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Dal Pra, I.; Armato, U.; Chioffi, F.; Pacchiana, R.; Whitfield, J.F.; Chakravarthy, B.; Gui, L.; Chiarini, A. The Abeta peptides-activated calcium-sensing receptor stimulates the production and secretion of vascular endothelial growth factor-A by normoxic adult human cortical astrocytes. Neuromol. Med. 2014, 16, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Bernaudin, M.; Nedelec, A.S.; Divoux, D.; MacKenzie, E.T.; Petit, E.; Schumann-Bard, P. Normobaric hypoxia induces tolerance to focal permanent cerebral ischemia in association with an increased expression of hypoxia-inducible factor-1 and its target genes, erythropoietin and VEGF, in the adult mouse brain. J. Cereb. Blood Flow Metab. 2002, 22, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Ghosh, S.; Wang, X.; Das, A.; Anderson, F.P.; Kukreja, R.C. Hypercholesterolemia enhances tolerance to lethal systemic hypoxia in middle-aged mice: Possible role of VEGF downregulation in brain. Mol. Cell. Biochem. 2006, 291, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Goochee, C.F.; Rasband, W.S.; Sokoloff, L. A computer-assisted image-processing system for the analysis of autoradiographs of cerebral metabolic activity. J. Med. Syst. 1980, 4, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Felix, B.; Leger, M.E.; Albe-Fessard, D.; Marcilloux, J.C.; Rampin, O.; Laplace, J.P. Stereotaxic atlas of the pig brain. Brain Res. Bull. 1999, 49, 1–137. [Google Scholar] [CrossRef]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jain, A.; Kratimenos, P.; Koutroulis, I.; Jain, A.; Buddhavarapu, A.; Ara, J. Effect of Intranasally Delivered rh-VEGF165 on Angiogenesis Following Cerebral Hypoxia-Ischemia in the Cerebral Cortex of Newborn Piglets. Int. J. Mol. Sci. 2017, 18, 2356. https://doi.org/10.3390/ijms18112356
Jain A, Kratimenos P, Koutroulis I, Jain A, Buddhavarapu A, Ara J. Effect of Intranasally Delivered rh-VEGF165 on Angiogenesis Following Cerebral Hypoxia-Ischemia in the Cerebral Cortex of Newborn Piglets. International Journal of Molecular Sciences. 2017; 18(11):2356. https://doi.org/10.3390/ijms18112356
Chicago/Turabian StyleJain, Amit, Panagiotis Kratimenos, Ioannis Koutroulis, Amishi Jain, Amulya Buddhavarapu, and Jahan Ara. 2017. "Effect of Intranasally Delivered rh-VEGF165 on Angiogenesis Following Cerebral Hypoxia-Ischemia in the Cerebral Cortex of Newborn Piglets" International Journal of Molecular Sciences 18, no. 11: 2356. https://doi.org/10.3390/ijms18112356
APA StyleJain, A., Kratimenos, P., Koutroulis, I., Jain, A., Buddhavarapu, A., & Ara, J. (2017). Effect of Intranasally Delivered rh-VEGF165 on Angiogenesis Following Cerebral Hypoxia-Ischemia in the Cerebral Cortex of Newborn Piglets. International Journal of Molecular Sciences, 18(11), 2356. https://doi.org/10.3390/ijms18112356