Femtograms of Interferon-γ Suffice to Modulate the Behavior of Jurkat Cells: A New Light in Immunomodulation

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. The Response of Jurkat Cells to IFN-γ
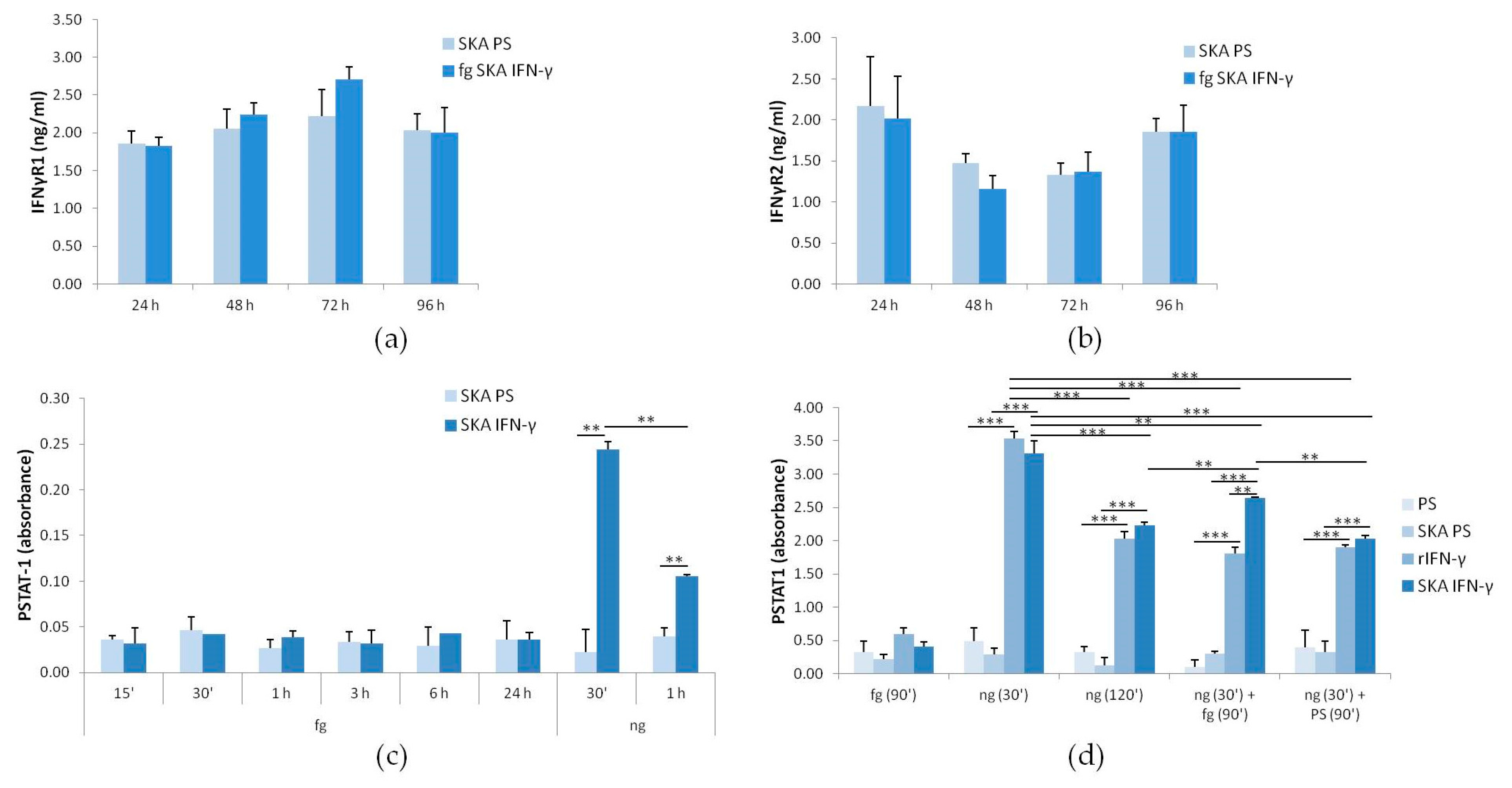
2.2. The Response of Jurkat Cells to Low Doses of SKA IFN-γ: IFN-γR1 and -R2 and STAT-1 Phosphorylation
2.3. The Response of Jurkat Cells to Low Doses of SKA IFN-γ: Expression of SOCS-1
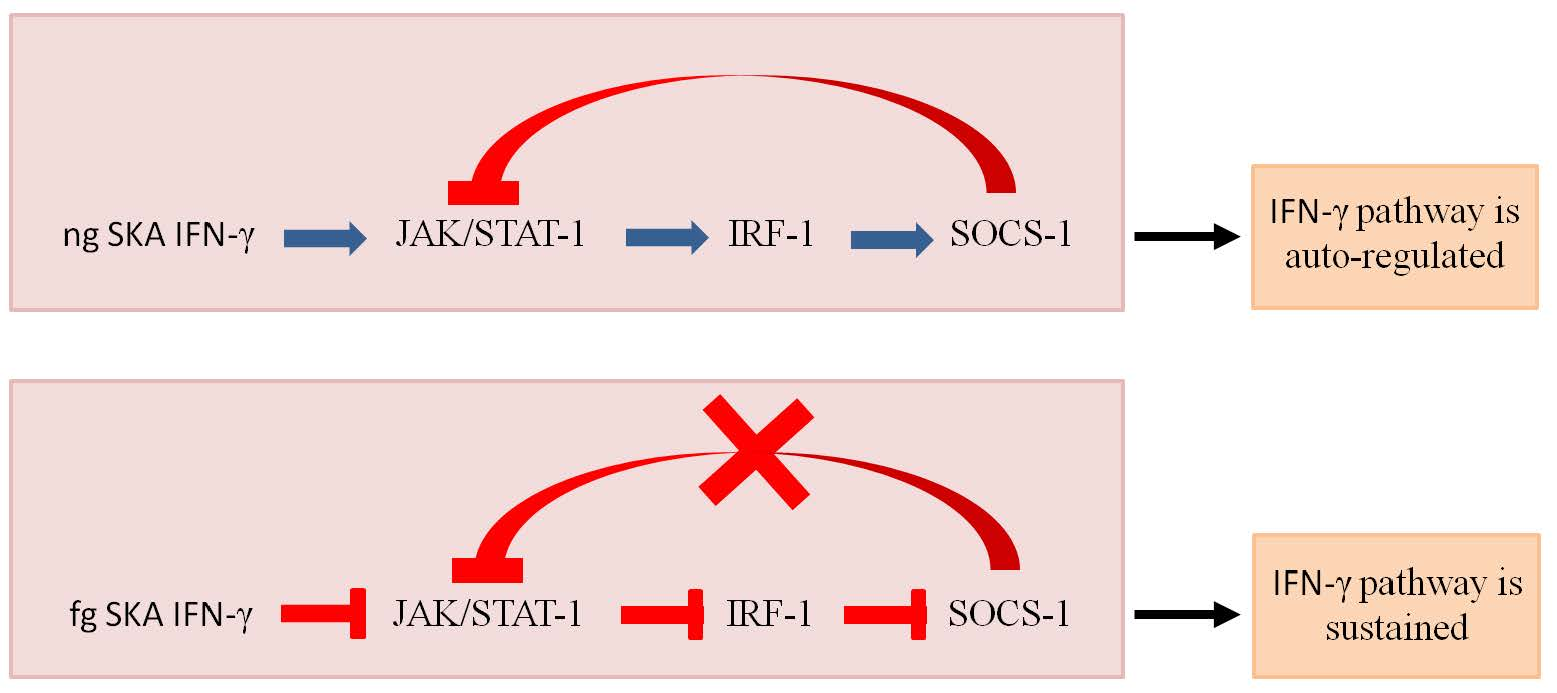
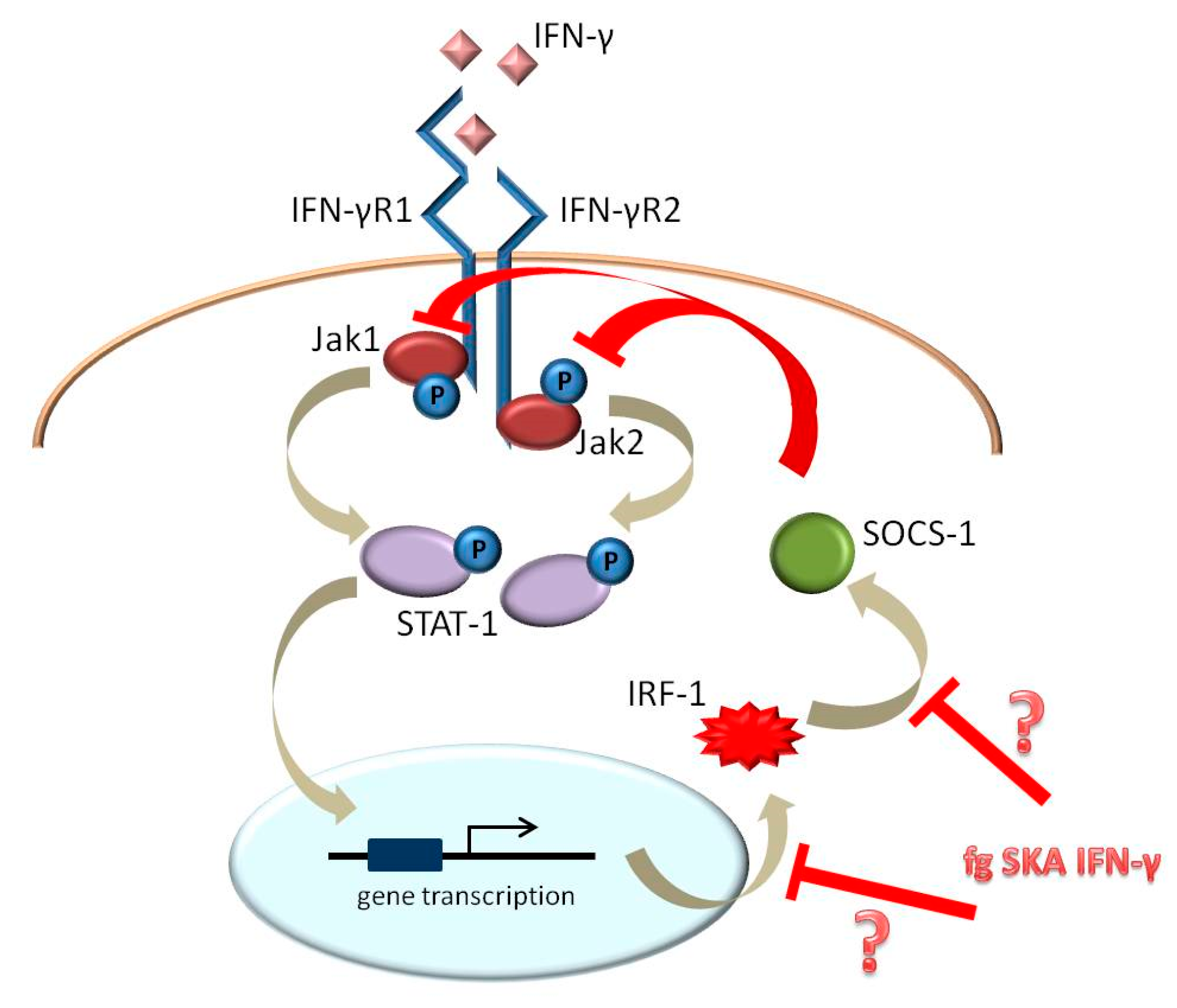
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. SKA Procedure
4.3. Real-Time PCR
4.4. ELISA
4.5. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Miller, C.H.T.; Maher, S.G.; Young, H.A. Clinical use of interferon-γ. Cytokine Ther. Ann. N. Y. Acad. Sci. 2009, 1182, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, M.R.; Merlino, G. The two faces of interferon-γ in cancer. Clin. Cancer Res. 2011, 17, 6118–6124. [Google Scholar] [CrossRef] [PubMed]
- Sakami, S.; Ishikawa, T.; Kawakami, N.; Haratani, T.; Fukui, A.; Kobayashi, F.; Fujita, O.; Araki, S.; Kawamura, N. Coemergence of insomnia and a shift in the Th1/Th2 balance toward Th2 dominance. Neuroimmunomodulation 2002, 10, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Majde, J.A.; Brown, R.A.; Krueger, J.M. Tumor necrosis factor receptor fragment attenuates interferon-γ-induced non-REM sleep in rabbits. J. Neuroimmunol. 2001, 119, 192–198. [Google Scholar] [CrossRef]
- Lin, F.C.; Young, H.A. The talented interferon-γ. Adv. Biosci. Biotechnol. 2013, 4, 6–13. [Google Scholar] [CrossRef]
- De Weerd, N.A.; Nguyen, T. The interferons and their receptors-distribution and regulation. Immunol. Cell Biol. 2012, 90, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Krause, C.D.; He, W.; Kotenko, S.; Pestka, S. Modulation of the activation of STAT1 by the interferon-γ receptor complex. Cell Res. 2006, 16, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef] [PubMed]
- Lotti, T.; Hercogova, J.; Wollina, U.; Chokoeva, A.A.; Zarrab, Z.; Gianfaldoni, S.; Roccia, M.G.; Fioranelli, M.; Tchernev, G. Treating skin diseases according to the low dose medicine principles. Data and hypotheses. J. Biol. Regul. Homeost. Agents 2015, 29, 47–51. [Google Scholar] [PubMed]
- Biancotto, A.; Wank, A.; Perl, S.; Cook, W.; Olnes, M.J.; Dagur, P.K.; Fuchs, J.C.; Langweiler, M.; Wang, E.; McCoy, J.P. Baseline levels and temporal stability of 27 multiplexed serum cytokine concentrations in healthy subjects. PLoS ONE 2013, 8, e76091. [Google Scholar] [CrossRef] [PubMed]
- Grazia, R.M.; Fioranelli, M. The history of low dose medicine research review of preclinical and clinical studies with low dose SKA cytokines since 2009. Microinflammation 2014, 1, 2. [Google Scholar]
- Roberti, M.L.; Ricottini, L.; Capponi, A.; Sclauzero, E.; Vicenti, P.; Fiorentini, E.; Savoia, C.; Scornavacca, G.; Brazioli, D.; Gaio, L.; et al. Immunomodulating treatment with low dose interleukin-4, interleukin-10 and interleukin-11 in psoriasis vulgaris. J. Biol. Regul. Homeost. Agents 2014, 28, 133–139. [Google Scholar] [PubMed]
- Martin-Martin, L.S.; Giovannangeli, F.; Bizzi, E.; Massafra, U.; Ballanti, E.; Cassol, M.; Migliore, A. An open randomized active-controlled clinical trial with low-dose SKAcytokines versus DMARDs evaluating low disease activity maintenance in patients with rheumatoid arthritis. Drug Des. Dev. Ther. 2017, 11, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Gariboldi, S.; Palazzo, M.; Zanobbio, L.; Dusio, G.F.; Mauro, V.; Solimene, U.; Cardani, D.; Mantovani, M.; Rumio, C. Low dose oral administration of cytokines for treatment of allergic asthma. Pulm. Pharmacol. Ther. 2009, 22, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Barygina, V.; Becatti, M.; Lotti, T.; Moretti, S.; Taddei, N.; Fiorillo, C. Treatment with low-dose cytokines reduces oxidative-mediated injury in perilesional keratinocytes from vitiligo skin. J. Dermatol. Sci. 2015, 79, 163–170. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, L.; Ruffini, E.; Ferracini, R.; Roato, I. Low dose of IL-12 stimulates T cell response in cultures of PBMCs derived from non-small cell lung cancer patients. J. Cancer Ther. 2012, 3, 337–342. [Google Scholar] [CrossRef]
- Radice, E.; Miranda, V.; Bellone, G. Low-doses of sequential-kinetic-activated interferon-γ enhance the ex vivo cytotoxicity of peripheral blood natural killer cells from subjects with early-stage colorectal cancer. A preliminary study. Int. Immunopharmacol. 2014, 19, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Radice, E.; Bellone, G.; Miranda, V. Enhancement of the immunostimulatory functions of ex vivo generated dendritic cells from early stage colon cancer subjects by consecutive exposure to low doses of sequential-kinetic-activated IL-4 and IL-12. A preliminary study. Transl. Oncol. 2015, 8, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Abraham, R.T.; Weiss, A. Jurkat T cells and development of the T-cell receptor signalling paradigm. Nat. Rev. Immunol. 2004, 4, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Alexander, W.S.; Starr, R.; Fenner, J.E.; Scott, C.L.; Handman, E.; Sprigg, N.S.; Corbin, J.E.; Cornish, A.L.; Darwiche, R.; Owczarek, C.M.; et al. SOCS1 is a critical inhibitor of interferon-γ signaling and prevents the potentially fatal neonatal actions of this cytokine. Cell 1999, 98, 597–608. [Google Scholar] [CrossRef]
- Andrè, N.; Tsai, K.; Carré, M.; Pasquier, E. Metronomic chemotherapy: Direct targeting of cancer cells after all? Trends Cancer 2017, 3, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Uberti, F.; Bardelli, C.; Morsanuto, V.; Ghirlanda, S.; Cochis, A.; Molinari, C. Stimulation of the nonneuronal cholinergic system by highly diluted acetylcholine in keratinocytes. Cells Tissues Organs 2017, 203, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Carello, R.; Ricottini, L.; Miranda, V.; Panei, P.; Rocchi, L.; Arcieri, R.; Galli, E. Long-term treatment with low-dose medicine in chronic childhood eczema: A double-blind two-stage randomized control trial. Ital. J. Pediatr. 2017, 43, 78. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, H.; Misawa, H.; Sakamoto, H.; Masuhara, M.; Sasaki, A.; Wakioka, T.; Ohtsuka, S.; Imaizumi, T.; Matsuda, T.; Ihle, J.N.; et al. The JAK-binding protein JAB inhibits Janus tyrosine kinase activity through binding in the activation loop. EMBO J. 1999, 18, 1309–1320. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castiglioni, S.; Miranda, V.; Cazzaniga, A.; Campanella, M.; Nichelatti, M.; Andena, M.; Maier, J.A.M. Femtograms of Interferon-γ Suffice to Modulate the Behavior of Jurkat Cells: A New Light in Immunomodulation. Int. J. Mol. Sci. 2017, 18, 2715. https://doi.org/10.3390/ijms18122715
Castiglioni S, Miranda V, Cazzaniga A, Campanella M, Nichelatti M, Andena M, Maier JAM. Femtograms of Interferon-γ Suffice to Modulate the Behavior of Jurkat Cells: A New Light in Immunomodulation. International Journal of Molecular Sciences. 2017; 18(12):2715. https://doi.org/10.3390/ijms18122715
Chicago/Turabian StyleCastiglioni, Sara, Vincenzo Miranda, Alessandra Cazzaniga, Marilena Campanella, Michele Nichelatti, Marco Andena, and Jeanette A. M. Maier. 2017. "Femtograms of Interferon-γ Suffice to Modulate the Behavior of Jurkat Cells: A New Light in Immunomodulation" International Journal of Molecular Sciences 18, no. 12: 2715. https://doi.org/10.3390/ijms18122715
APA StyleCastiglioni, S., Miranda, V., Cazzaniga, A., Campanella, M., Nichelatti, M., Andena, M., & Maier, J. A. M. (2017). Femtograms of Interferon-γ Suffice to Modulate the Behavior of Jurkat Cells: A New Light in Immunomodulation. International Journal of Molecular Sciences, 18(12), 2715. https://doi.org/10.3390/ijms18122715