
Differential Gene Expression in the Meristem and during Early Fruit Growth of Pisum sativum L. Identifies Potential Targets for Breeding


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
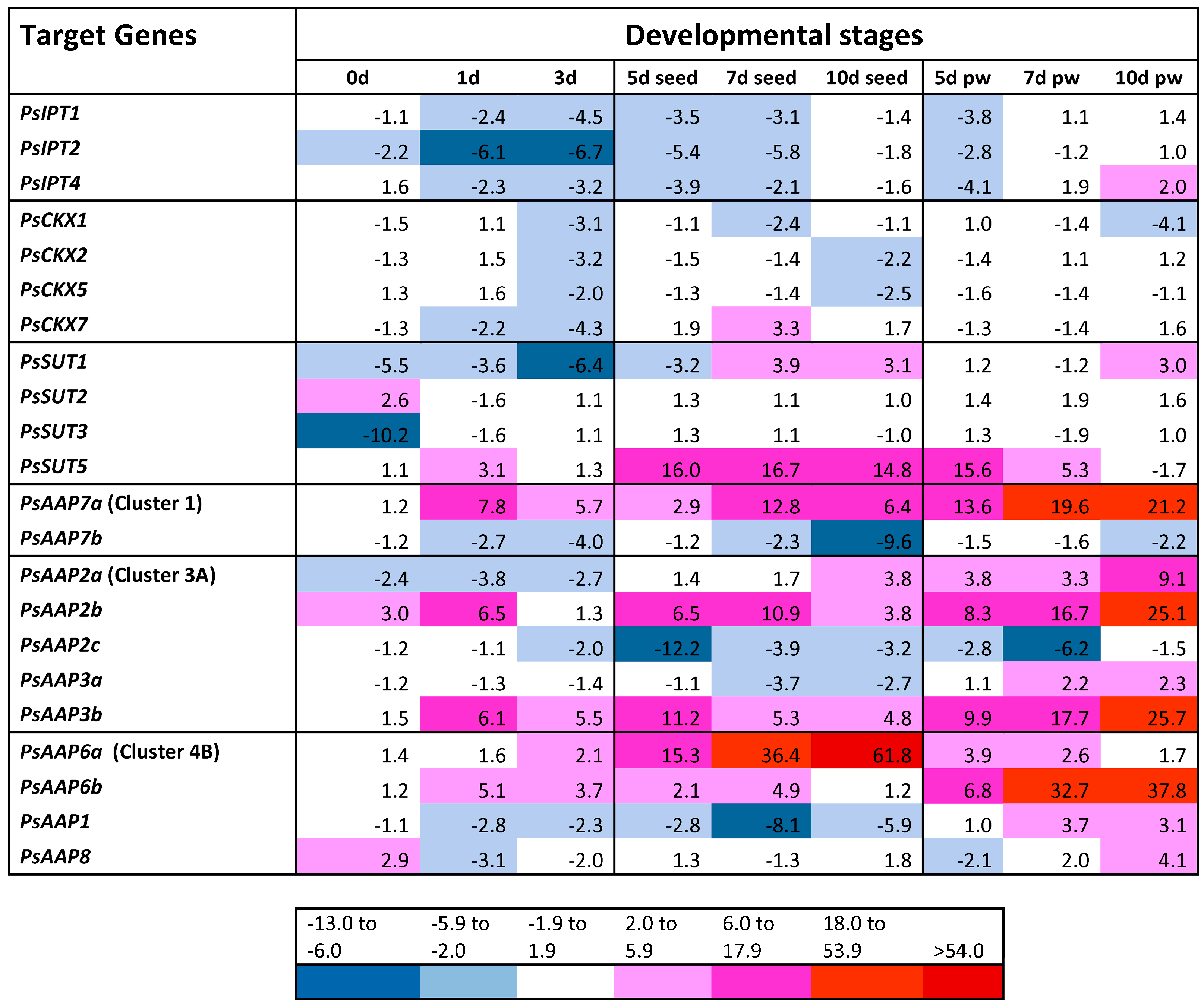
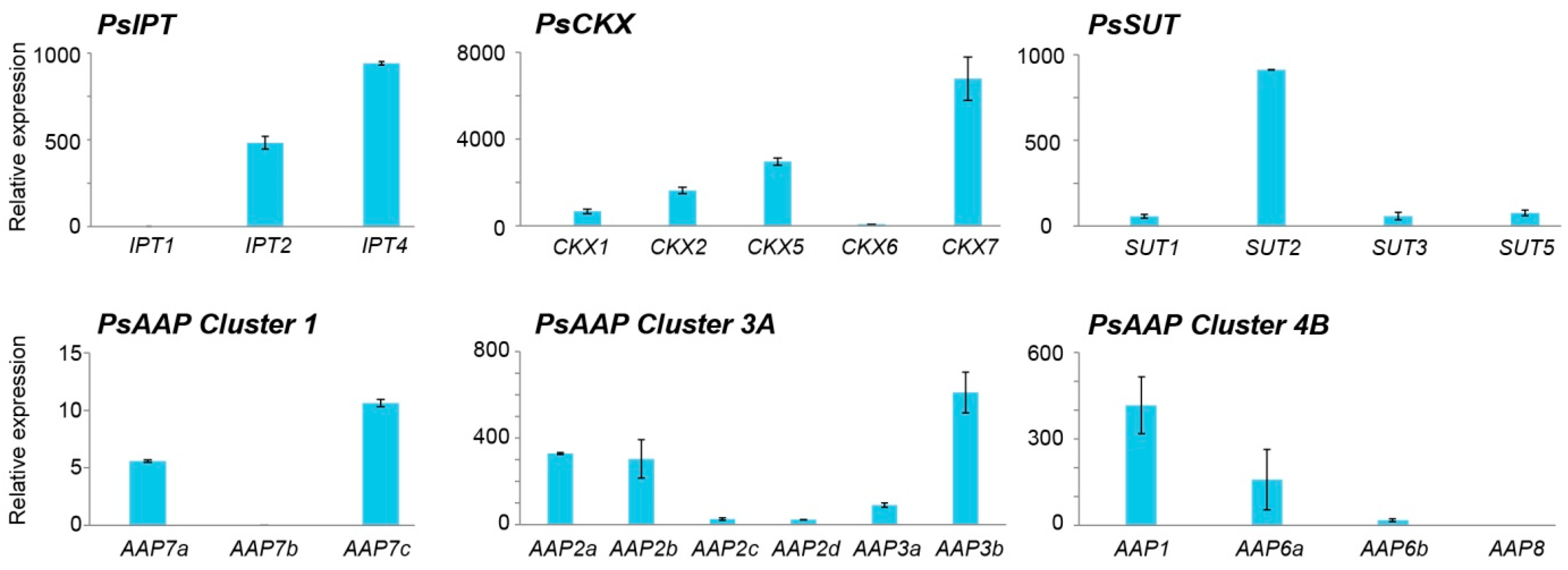
2.1. Differential Gene Expression in Fruits
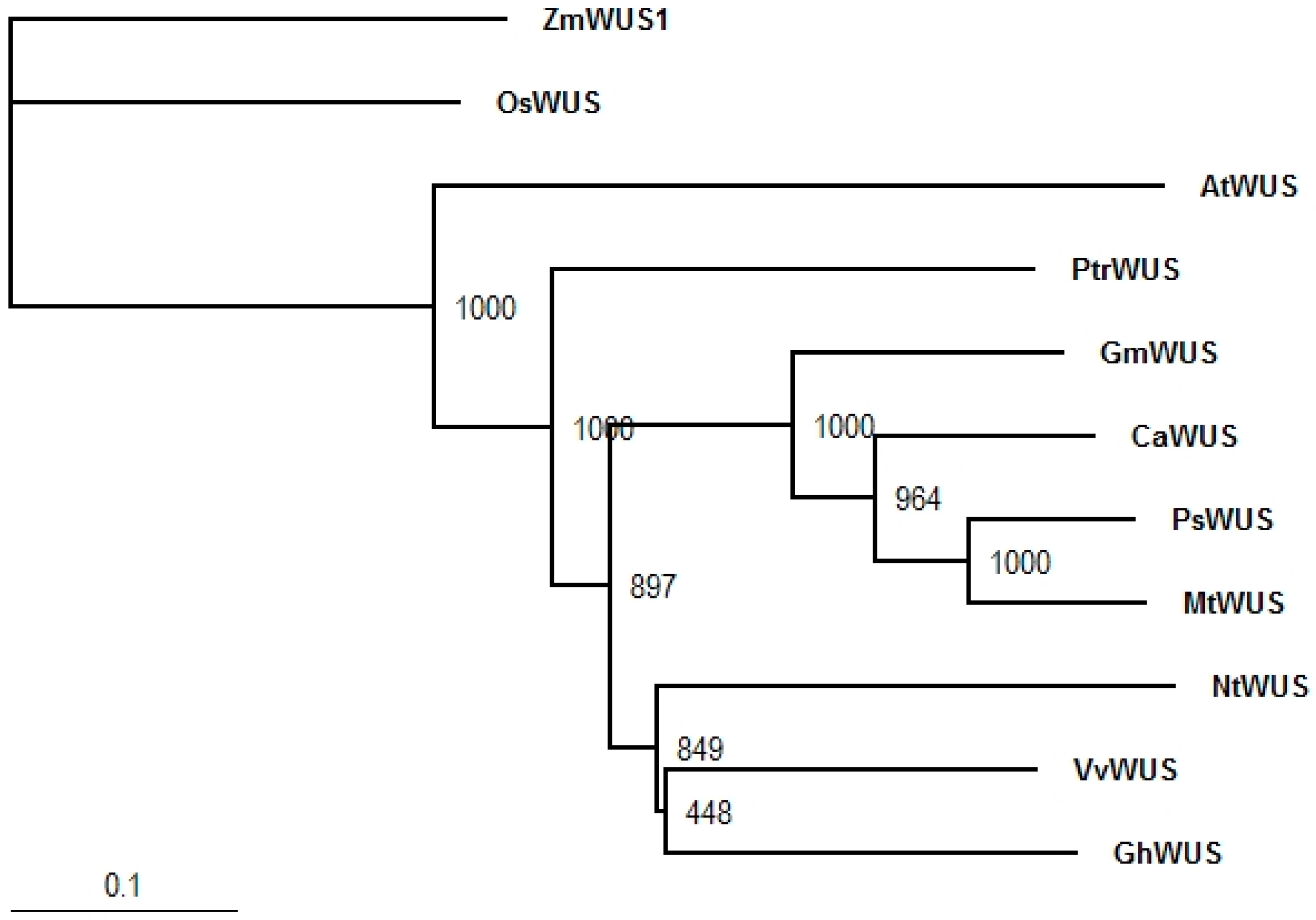
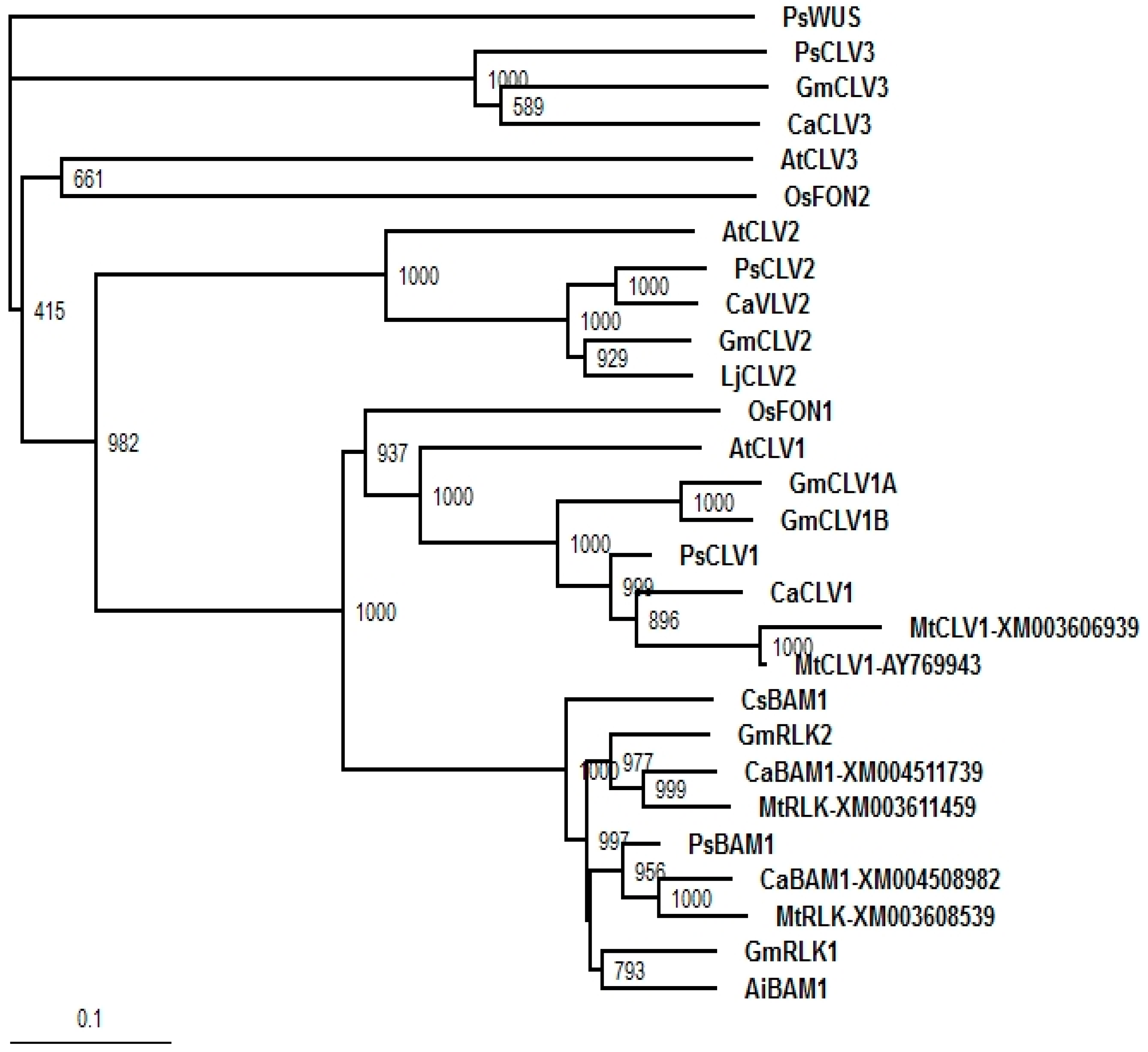
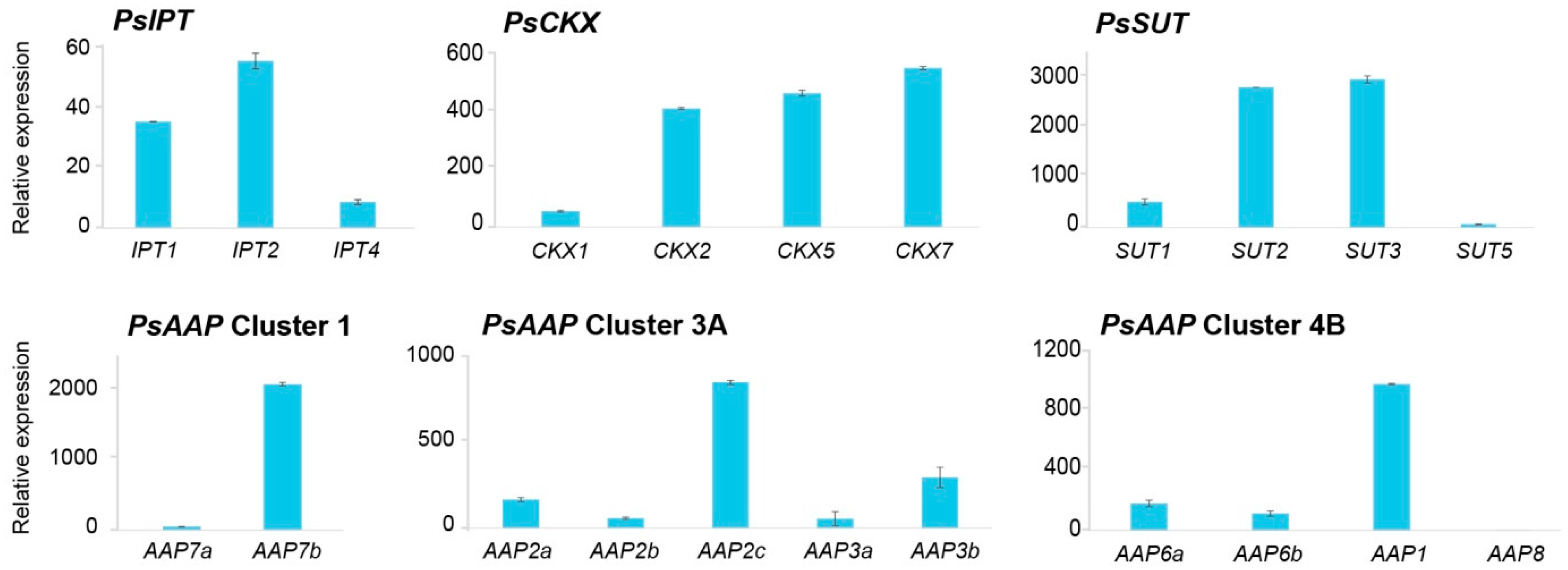
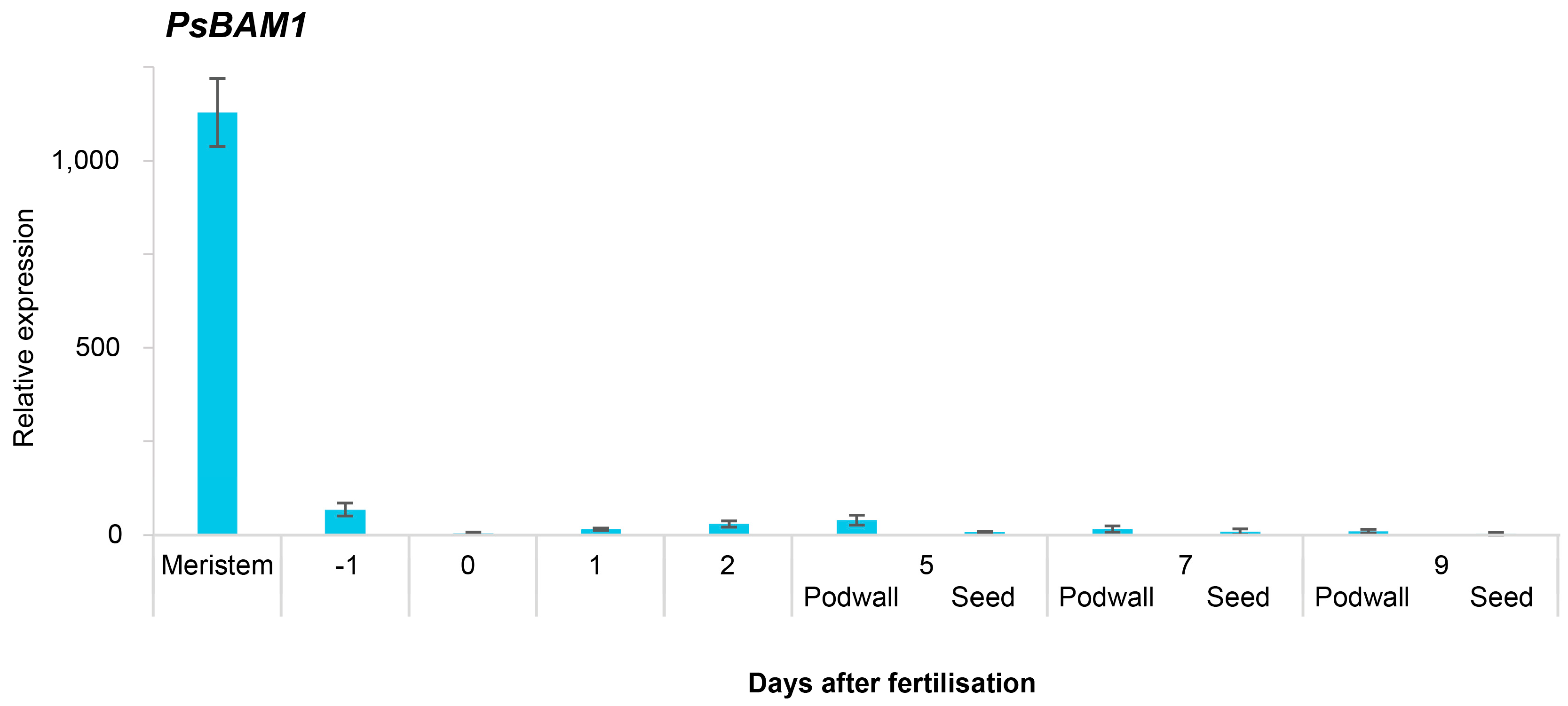
2.2. Gene Expression in the Shoot Apical Meristem
3. Discussion
4. Materials and Methods
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zohary, D.H.M. Domestication of Plants in the Old World; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Wikipedia Page: Pea. Available online: https://en.wikipedia.org/wiki/Pea (accessed on 9 August 2016).
- Tegeder, M. Transporters involved in source to sink partitioning of amino acids and ureides: Opportunities for crop improvement. J. Exp. Bot. 2014, 65, 1865–1878. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Garneau, M.G.; Majumdar, R.; Gtant, J.; Tegeder, M. Improvement of pea biomass and seed productivity by simultaneous increase of phloem and embryo loading with amino acids. Plant J. 2015, 81, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Yadav, U.P.; Ayre, B.G.; Bush, D.R. Transgenic approaches to altering carbon and nitrogen partitioning in whole plants: Assessing the potential to improve crop yields and nutritional quality. Front. Plant Sci. 2015, 6, 275. [Google Scholar] [CrossRef] [PubMed]
- Jameson, P.E.; Song, J. Cytokinin: A key driver of seed yield. J. Exp. Bot. 2016, 67, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Crosby, K.E.; Aung, L.H.; Buss, G.R. Influence of 6-benzylaminopurine on fruit-set and seed development in two soybean, Glycine max (L.) Merr. genotypes. Plant Physiol. 1981, 68, 985–988. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.R.; Dyer, D.J.; Cotterman, C.D.; Durley, R.C. The physiological-basis for cytokinin induced increases in pod set in IX93-100 soybeans. Plant Physiol. 1987, 84, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Dyer, D.J.; Carlson, D.R.; Cotterman, C.D.; Sikorski, J.A.; Ditson, S.L. Soybean pod set enhancement with synthetic cytokinin analogs. Plant Physiol. 1987, 84, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.M.; Williams, J.C.; Kuang, A.X. Increased pod set of determinate cultivars of soybean, Glycine max, with 6-benzylaminopurine. Bot. Gaz. 1990, 151, 322–330. [Google Scholar] [CrossRef]
- Aikins, C.A.; Pigeaire, A. Applications of cytokinins to flowers to increase pod set in Lupinus angustifolius. Aust. J. Agric. Res. 1993, 44, 1799–1819. [Google Scholar] [CrossRef]
- Nagel, L.; Brewster, R.; Riedell, W.E.; Reese, R.N. Cytokinin regulation of flower and pod set in soybeans (Glycine max (L.) Merr.). Ann. Bot. 2001, 88, 27–31. [Google Scholar] [CrossRef]
- Cho, Y.; Suh, S.K.; Park, H.K.; Wood, A. Impact of 2,4-DP and BAP upon pod set and seed yield of soybean treated at reproductive stages. Plant Growth Regul. 2002, 36, 215–221. [Google Scholar] [CrossRef]
- Aikins, C.A.; Emery, R.J.N.; Smith, P.M.C. Consequences of transforming narrow leafed lupin (Lupinus angustifolius L.) with an ipt gene under control of a flower-specific promoter. Transgenic Res. 2011, 20, 1321–1332. [Google Scholar] [CrossRef] [PubMed]
- Ashikari, M.; Sakakibara, H.; Lin, S.Y.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin regulates the activity of reproductive meristems, flower organ size, ovule formation, and thus seed yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Hirose, N.; Takei, K.; Huroha, T.; Kamada-Nobusada, T.; Hayashi, H.; Sakakibara, H. Regulation of cytokinin biosynthesis, compartmentalization and translocation. J. Exp. Bot. 2008, 59, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Jiang, L.; Jameson, P.E. Co-ordinate regulation of cytokinin gene family members during flag leaf and reproductive development in wheat. BMC Plant Biol. 2012, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Jiang, L.; Jameson, P.E. Expression patterns of Brassica napus genes implicate IPT, CKX, sucrose transporter, cell wall invertase and amino acid permease gene family members in leaf, flower, silique and seed development. J. Exp. Bot. 2015, 66, 5067–5082. [Google Scholar] [CrossRef] [PubMed]
- Leibfried, A.; To, J.P.C.; Busch, W.; Stehling, S.; Kehle, A.; Demar, M.; Kieber, J.J.; Lohmann, J.U. Wuschel controls meristem function by direct regulation of cytokinin-inducible response regulators. Nature 2005, 438, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.P.; Chickarmane, V.S.; Ohno, C.; Meyerowitz, E.M. Multiple feedback loops through cytokinin signaling control stem cell number within the Arabidopsis shoot meristem. Proc. Natl. Acad. Sci. USA 2009, 106, 16529–16534. [Google Scholar] [CrossRef] [PubMed]
- Galli, M.; Gallavotti, A. Expanding the regulatory network for meristem size in plants. Trends Genet. 2016, 32, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer, K.F.; Jürgens, G.; Laux, T. The stem cell population of Arabidopsis shoot meristems in maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 2000, 100, 635–644. [Google Scholar] [CrossRef]
- Buechel, S.; Leibfried, A.; To, J.P.C.; Zhao, Z.; Andersen, S.U.; Kieber, J.J.; Lohmann, J.U. Role of A-type Arabidopsis response regulators in meristem maintenance and regeneration. Eur. J. Cell Biol. 2010, 89, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Hazak, O.; Hardtke, C.S. CLAVATA 1-type receptors in plant development. J. Exp. Bot. 2016, 67, 4827–4833. [Google Scholar] [CrossRef] [PubMed]
- Deyoung, B.J.; Clark, S.E. BAM receptors regulate stem cell specification and organ development through complex interactions with CLAVATA signaling. Genetics 2008, 180, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Nimchuk, Z.L.; Zhou, Y.; Tarr, P.L.; Peterson, B.A.; Meyerowitz, E.M. Plant stem cell maintenance by transcriptional cross-regulation of related receptor kinases. Development 2015, 142, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, H.; Matsuayashi, Y. Reevaluation of the CLV3-receptor interaction in the shoot apical meristem: Dissection of the LV3 signaling pathway from a direct ligand-binding point of view. Plant J. 2015, 82, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Wong, C.E.; Singh, M.B.; Beveridge, C.A.; Phipson, B.; Smyth, G.K.; Bhalla, P.L. Molecular dissection of the pea shoot apical meristem. J. Exp. Bot. 2009, 60, 4201–4213. [Google Scholar] [CrossRef] [PubMed]
- Emery, R.J.N.; Ma, Q.; Atkins, C.A. The forms and sources of cytokinins in developing white lupine seeds and fruits. Plant Physiol. 2000, 123, 1593–1604. [Google Scholar] [CrossRef] [PubMed]
- De Jong, J.W.K.K.; Schuurmans, M.J.; Borstlap, A.C. The mechanism of amino acid efflux from seed coats of developing pea seeds as revealed by uptake experiments. Plant Physiol. 1997, 114, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Sundaresan, V. Control of seed size in plants. Proc. Natl. Acad. Sci. USA 2005, 102, 17887–17888. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, M.; Naganuma, T.; Tsutsumi, K.-I.; Saitoh, Y. The syncytium-specific expression of the Orysa; KRP3 CDK inhibitor: Implication of its involvement in the cell cycle control in the rice (Oryza sativa L.) syncytial endosperm. J. Exp. Bot. 2010, 61, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Weber, H. Molecular physiology of legume seed development. Annu. Rev. Plant Biol. 2005, 56, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W-H.; Zhou, Y.; Dibley, K.E.; Tyerman, S.D.; Furbank, R.T.; Patrick, J.W. Nutrient loading of developing seeds. Func. Plant Biol. 2007, 34, 314–331. [Google Scholar] [CrossRef]
- Tan, Q.; Zhang, L.; Grant, J.; Cooper, P.; Tegeder, M. Increased phloem transport of S-methylmethionine positively affects sulfur and nitrogen metabolism and seed development in pea plants. Plant Physiol. 2010, 154, 1886–1896. [Google Scholar] [CrossRef] [PubMed]
- Rosche, E.G.; Blackmore, D.; Offler, C.E.; Patrick, J.W. Increased capacity for sucrose uptake leads to earlier onset of protein accumulation in developing pea seeds. Funct. Plant Biol. 2005, 32, 997–1007. [Google Scholar] [CrossRef]
- Rolletschek, H.; Hosein, F.; Miranda, M.; Heim, U.; Götz, K.-P.; Schlereth, A.; Borisjuk, L.; Saalbach, I.; Wobus, U.; Weber, H. Ectopic expression of an amino acid transporter (VfAAP1) in seeds of Vicia narbonensis and pea increases storage proteins. Plant Physiol. 2005, 137, 1236–1249. [Google Scholar] [CrossRef] [PubMed]
- Gotz, K.-P.; Staroske, N.; Radchuk, R.; Emery, R.J.N.; Wutze, K.-D.; Herzog, H.; Weber, H. Uptake and allocation of carbon and nitrogen in Vicia narbonensis plants with increased seed sink strength achieved by seed-specific expression of an amino acid permease. J. Exp. Bot. 2007, 58, 3183–3195. [Google Scholar] [CrossRef] [PubMed]
- Dhandapani, P.; Song, J.; Novak, O.; Jameson, P.E. Infection by Rhodococcus fascians maintains cotyledons as a sink tissue for the pathogen. Ann. Bot. 2016. [Google Scholar] [CrossRef] [PubMed]
- Mayar, K.F.X.; Schoof, H.; Haecker, A.; Lenhard, M.; Jürgens, G.; Lauz, T. Role of WUSCHEL in regulating stem cell fate in Arabidopsis shoot meristem. Cell 1998, 95, 805–815. [Google Scholar] [CrossRef]
- Wong, C.E.; Khor, S.Y.; Bhalla, P.L.; Singh, M.B. Novel spatial expression of soybean WUSCHEL in the incipient floral primordia. Planta 2011, 233, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.E.; Singh, M.; Bhalla, P.L. Spatial expression of CLAVATA3 in the shoot apical meristem suggests it is not a stem cell marker in soybean. J. Exp. Bot. 2013, 64, 5641–5649. [Google Scholar] [CrossRef] [PubMed]
- Sturm, A. Invertases. Primary structures, functions, and roles in plant development and sucrose partitioning. Plant Physiol. 1999, 121, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kühn, C.; Grof, C.P.L. Sucrose transporters of higher plants. Curr. Opin. Plant Biol. 2010, 13, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.P.; Tegeder, M. Connecting source with sink: The role of Arabidpsis AAP8 in phloem loading of amino acids. Plant Physiol. 2016, 171, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Okumoto, S.; Schmidt, R.; Tegeder, M.; Fischer, W.N.; Rentsch, D.; Frommer, W.B.; Koch, W. High affinity amino acid transporters specifically expressed in xylem parenchyma and developing seeds of Arabidopsis. J. Biol. Chem. 2002, 277, 45338–45346. [Google Scholar] [CrossRef] [PubMed]
- Jameson, P.E.; Dhandapani, P.; Novak, O.; Song, J. Cytokinins and expression of SWEET, SUT, CWINV and AAP genes increase as pea seeds germinate. Int. J. Mol. Sci. 2016, 17, 2013. [Google Scholar] [CrossRef] [PubMed]
- Ando, S.; Asano, T.; Tsushima, S.; Kamachi, S.; Hagio, T.; Tabei, Y. Changes in gene expression of putative isopentenyl transferase during clubroot development in Chinese cabbage (Brassica rapa L.). Physiol. Mol. Plant Pathol. 2005, 67, 59–67. [Google Scholar] [CrossRef]
- O’Keefe, D.; Song, J.; Jameson, P.E. Isopentenyl transferase and cytokinin oxidase/dehydrogenase gene family members are differentially expressed during pod and seed development in Rapid-cycling Brassica rapa L. J. Plant Growth Regul. 2011, 30, 92–99. [Google Scholar] [CrossRef]
- Liu, Z.; Lv, Y.; Zhang, M.; Liu, Y.; Kong, L.; Zou, M.; Lu, G.; Cao, J.; Yu, X. Identification, expression, and comparative genomic analysis of the IPT and CKX genes families in Chinese cabbage (Brassica rapa ssp. pekinensis). BMC Genom. 2013, 14, 594. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Vankova, R.; Tanaka, M.; Seki, M.; le Ham, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Identification and expression analysis of cytokinin metabolic genes in soybean under normal and drought conditions in relation to cytokinin levels. PLoS ONE 2012, 7, e42411. [Google Scholar] [CrossRef] [PubMed]
- Motyka, V.; Vaňková, R.; Čapková, V.; Petrášek, J.; Kamínek, M.; Schmülling, T. Cytokinin-induced upregulation of cytokinin oxidase activity in tobacco includes changes in enzyme glycosylation and secretion. Physiol. Plant 2003, 117, 11–21. [Google Scholar] [CrossRef]
- Brugière, N.; Jiao, S.; Hanke, S.; Zinselmeier, C.; Roessler, J.A.; Niu, X.; Jones, R.J.; Habben, J.E. Cytokinin oxidase gene expression in maize is localized to the vasculature, and is induced by cytokinins, abscisic acid, and abiotic stress. Plant Physiol. 2003, 132, 1228–1240. [Google Scholar] [CrossRef] [PubMed]
- Zalewski, W.; Gasparis, S.; Boczkowska, M.; Rajchel, I.; Orczyk, W.; Nadolska-Orczyk, A. Expression patterns of HvCKX genes indicate their role in growth and reproductive development of barley. PLoS ONE 2014, 9, e115729. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smitha Ninan, A.; Shah, A.; Song, J.; Jameson, P.E. Differential Gene Expression in the Meristem and during Early Fruit Growth of Pisum sativum L. Identifies Potential Targets for Breeding. Int. J. Mol. Sci. 2017, 18, 428. https://doi.org/10.3390/ijms18020428
Smitha Ninan A, Shah A, Song J, Jameson PE. Differential Gene Expression in the Meristem and during Early Fruit Growth of Pisum sativum L. Identifies Potential Targets for Breeding. International Journal of Molecular Sciences. 2017; 18(2):428. https://doi.org/10.3390/ijms18020428
Chicago/Turabian StyleSmitha Ninan, Annu, Anish Shah, Jiancheng Song, and Paula E. Jameson. 2017. "Differential Gene Expression in the Meristem and during Early Fruit Growth of Pisum sativum L. Identifies Potential Targets for Breeding" International Journal of Molecular Sciences 18, no. 2: 428. https://doi.org/10.3390/ijms18020428
APA StyleSmitha Ninan, A., Shah, A., Song, J., & Jameson, P. E. (2017). Differential Gene Expression in the Meristem and during Early Fruit Growth of Pisum sativum L. Identifies Potential Targets for Breeding. International Journal of Molecular Sciences, 18(2), 428. https://doi.org/10.3390/ijms18020428