
Zebrafish as an Alternative Vertebrate Model for Investigating Developmental Toxicity—The Triadimefon Example

Abstract
:1. Introduction
2. Results and Discussion
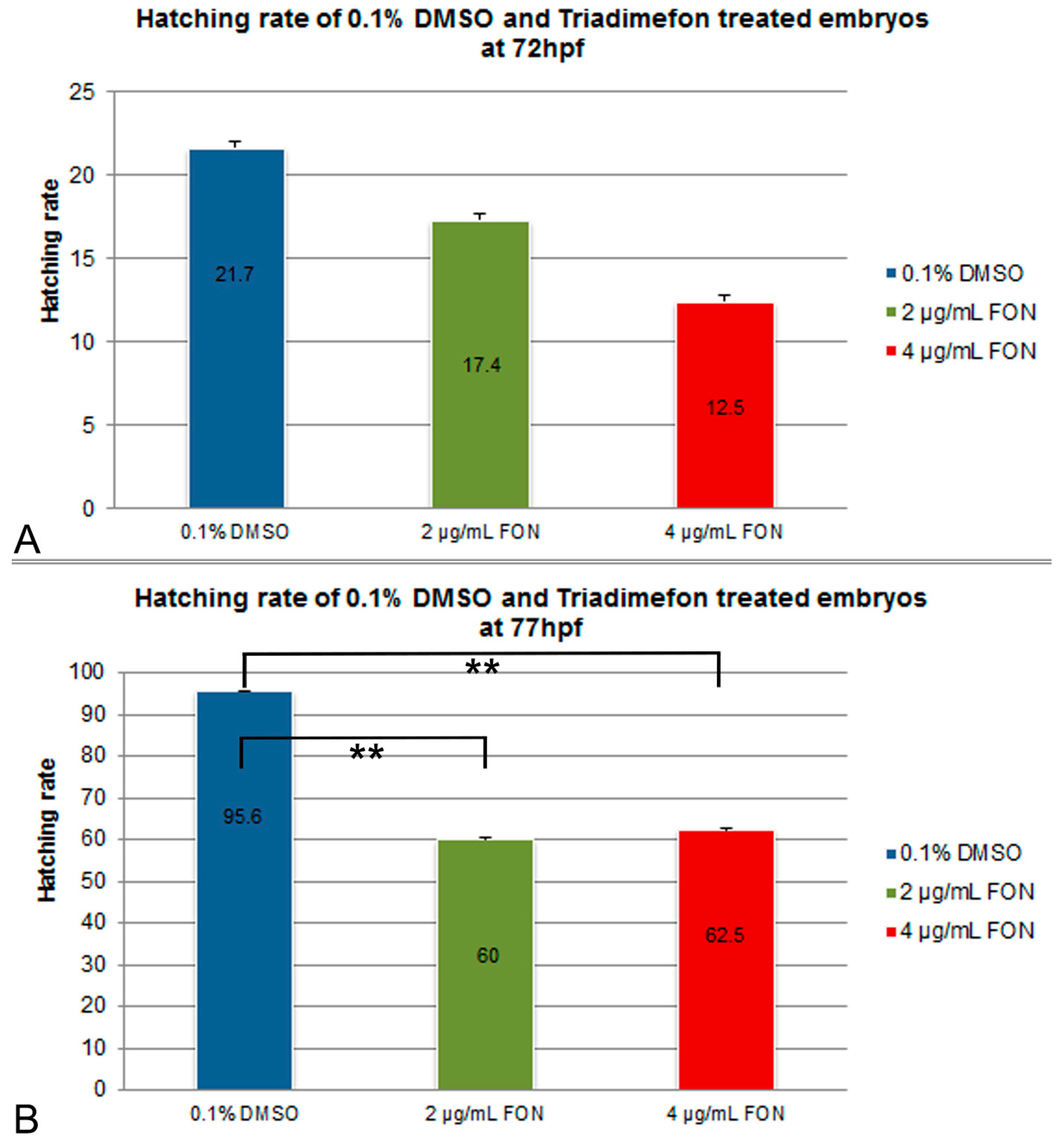
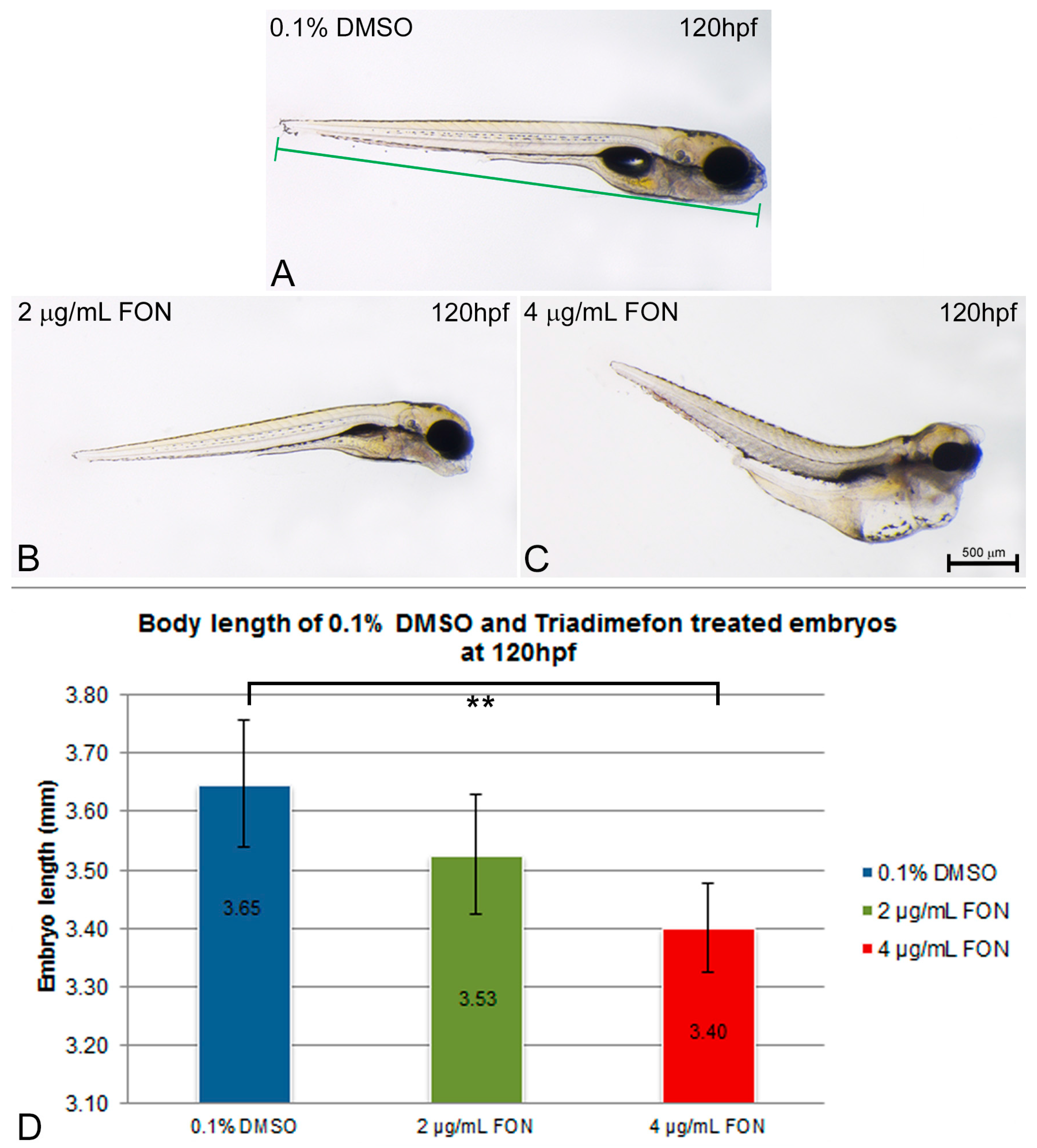
2.1. Observations on Hatching Rate and Length Growth
2.2. Skeletal and Soft Tissue Effects
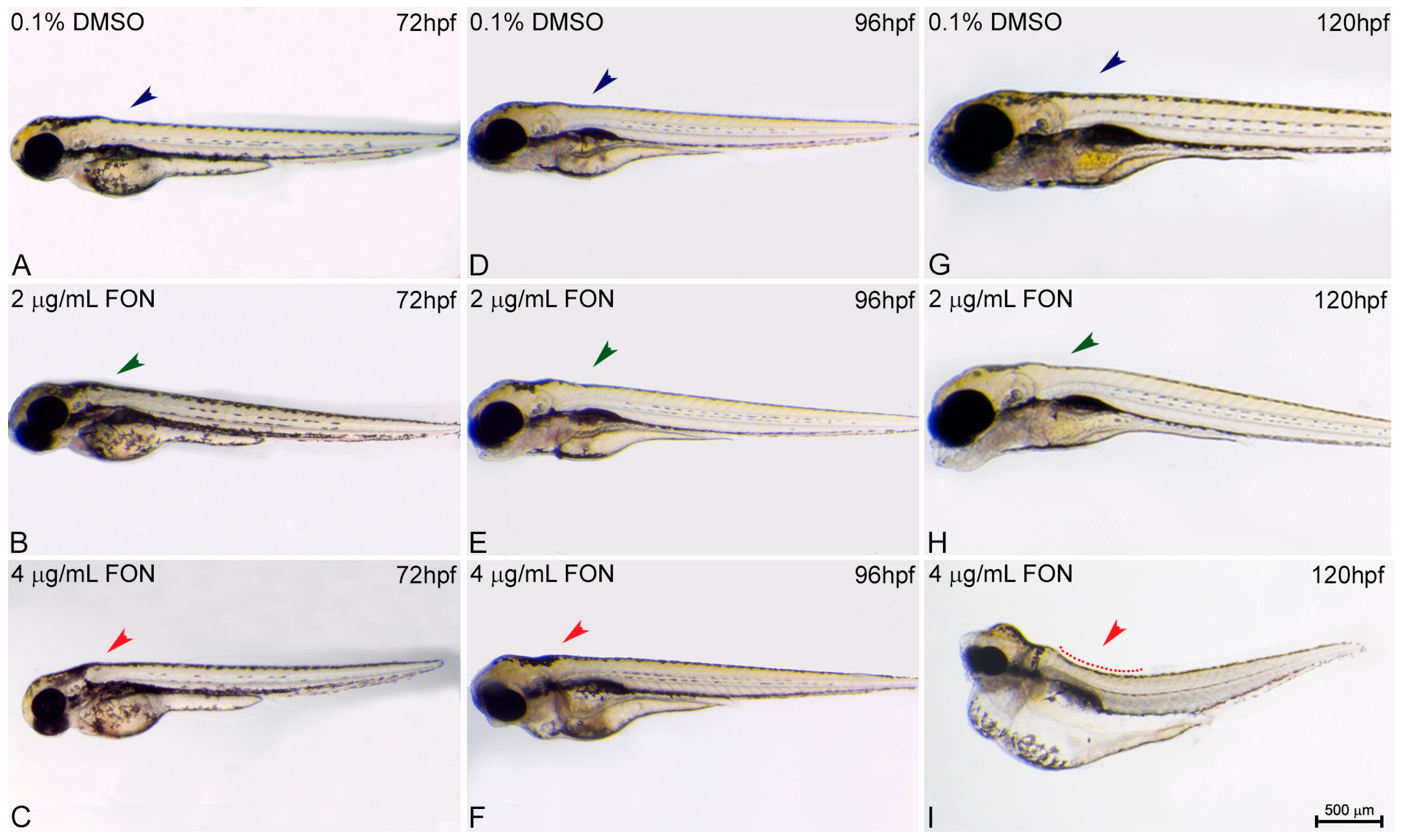
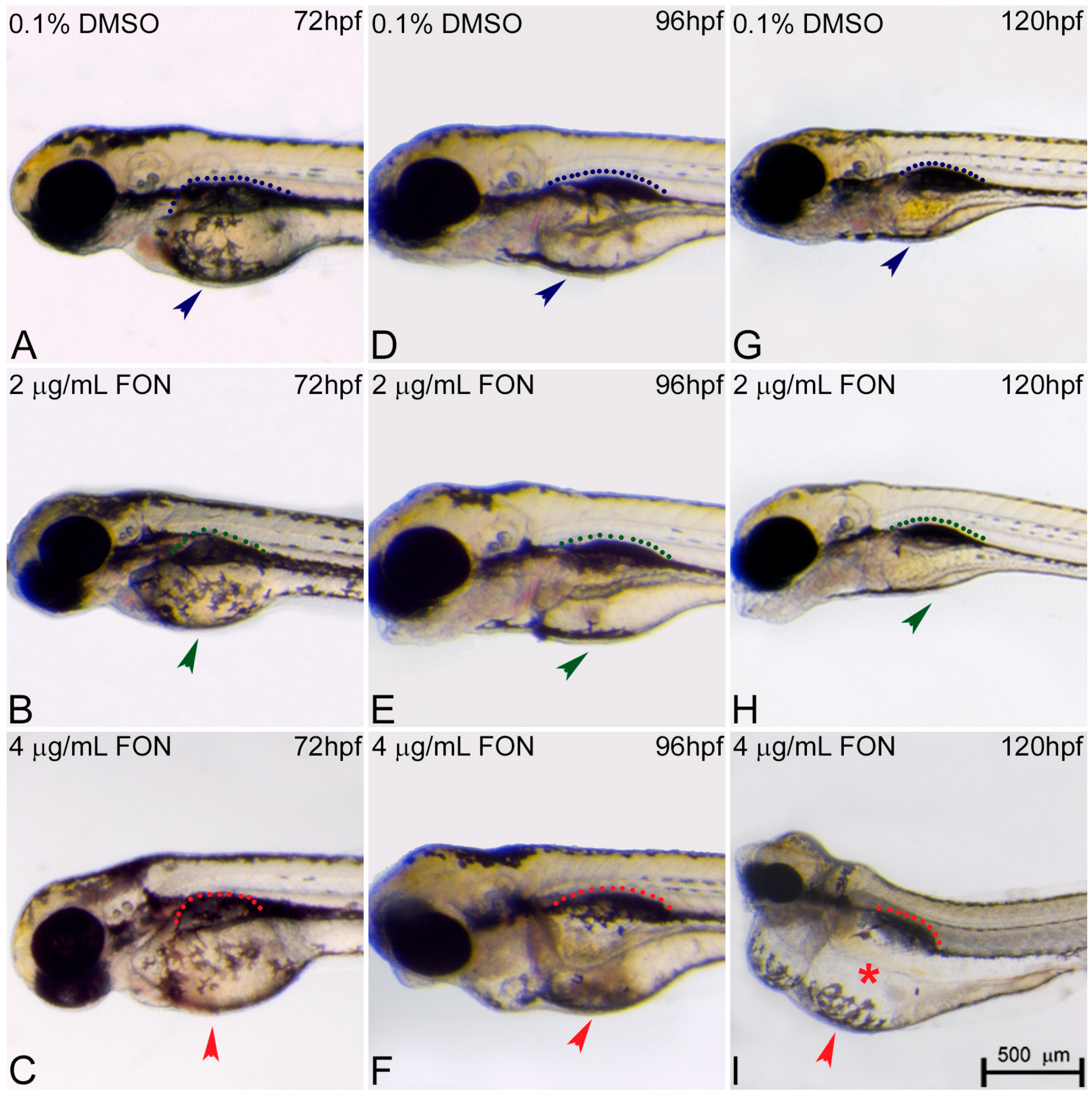
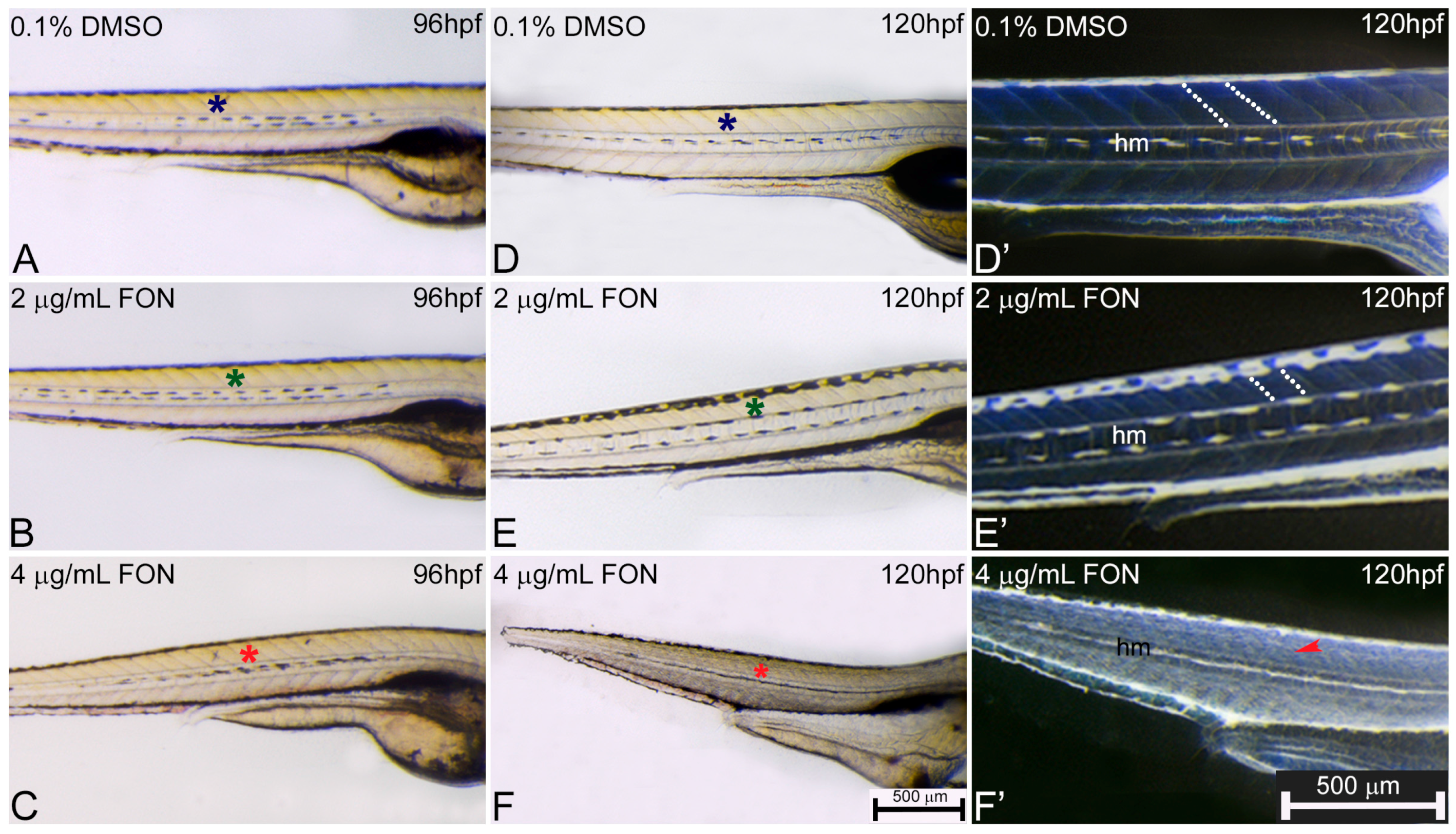
2.3. Alternations in Somitic, Myotome and Horizontal Myoseptum Formation Following Triadimefon Exposure
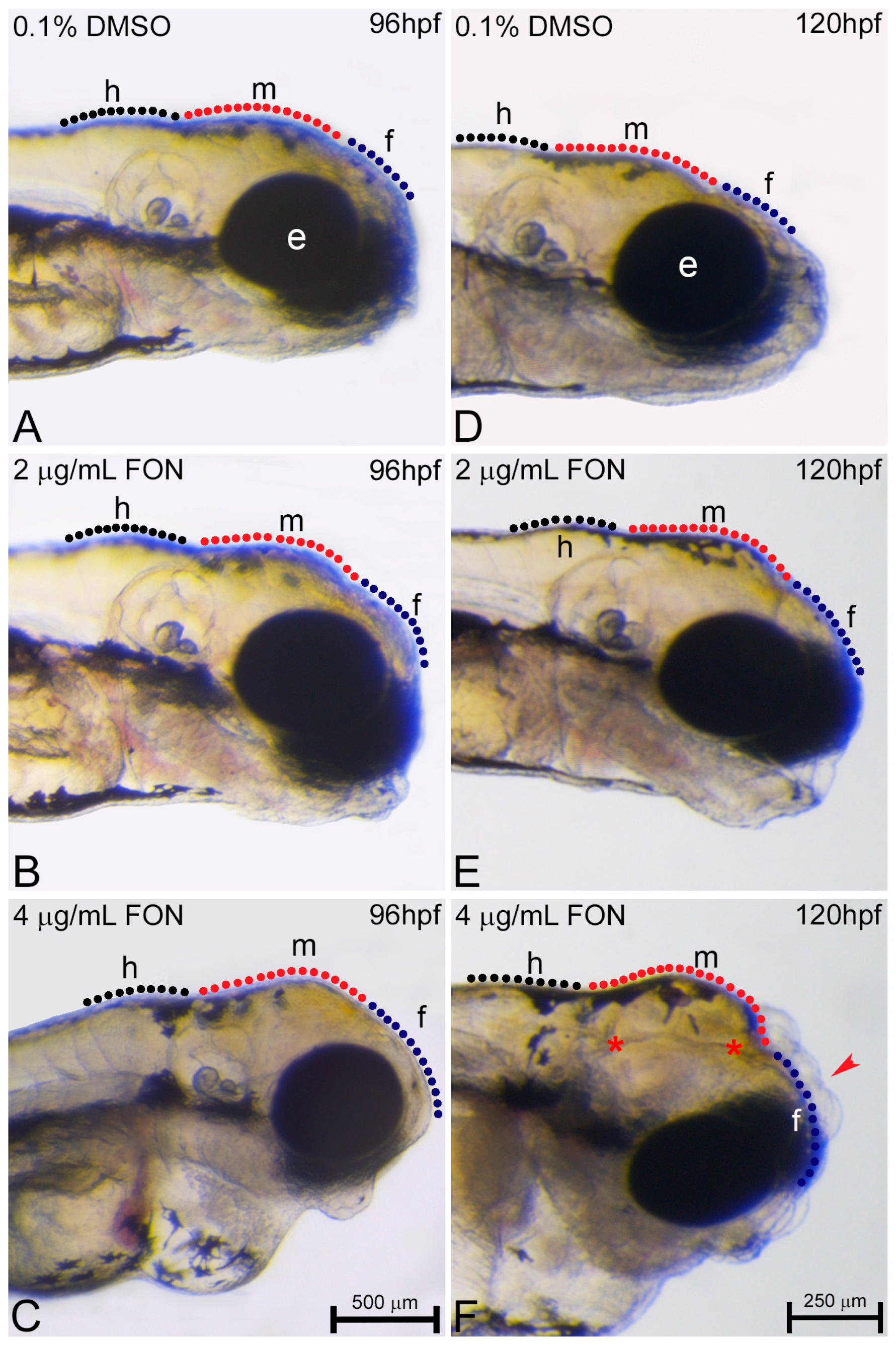
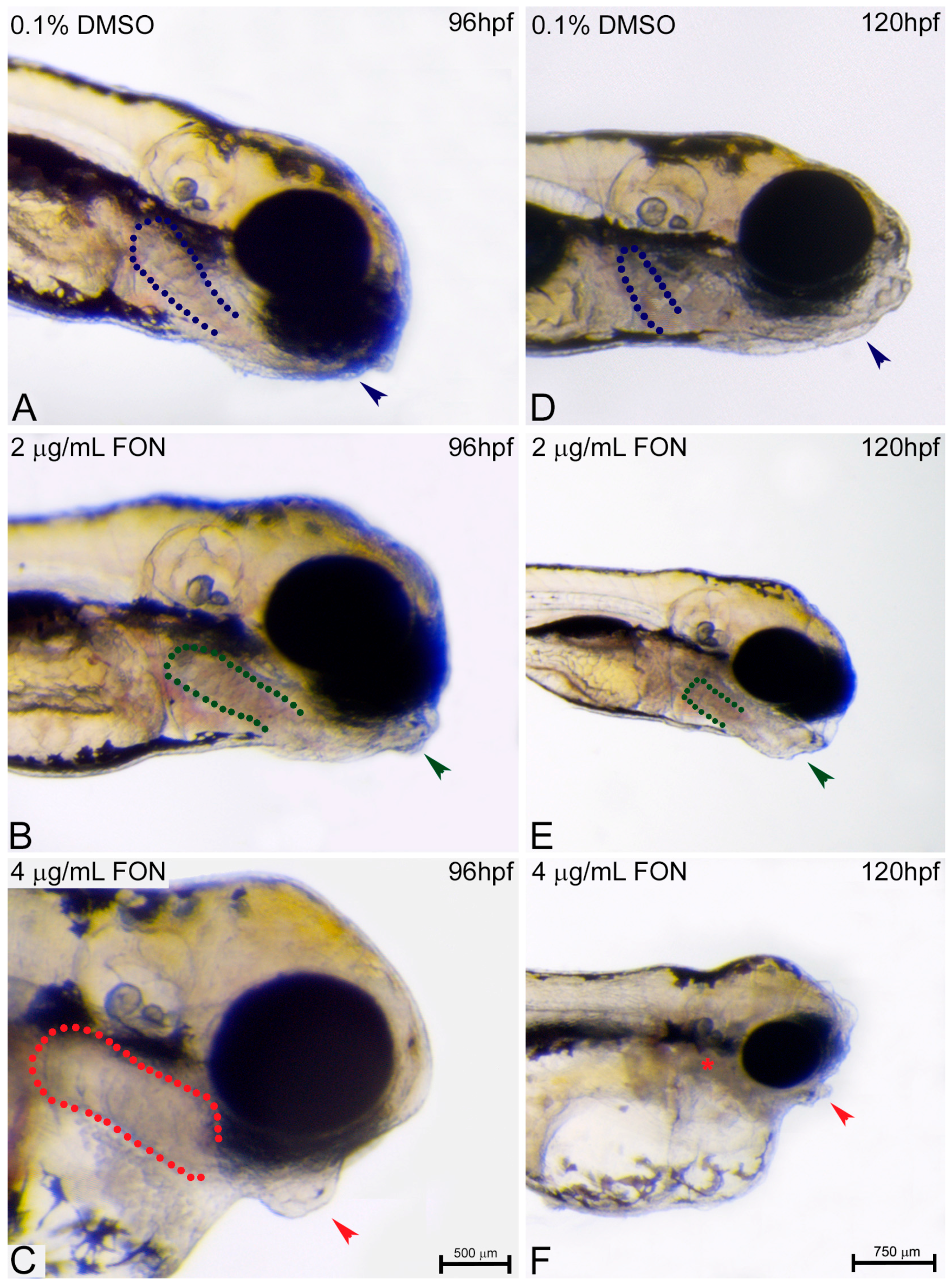
2.4. Triadimefon Leads to Craniofacial and Pharyngeal Arch Defects in Zebrafish Embryos
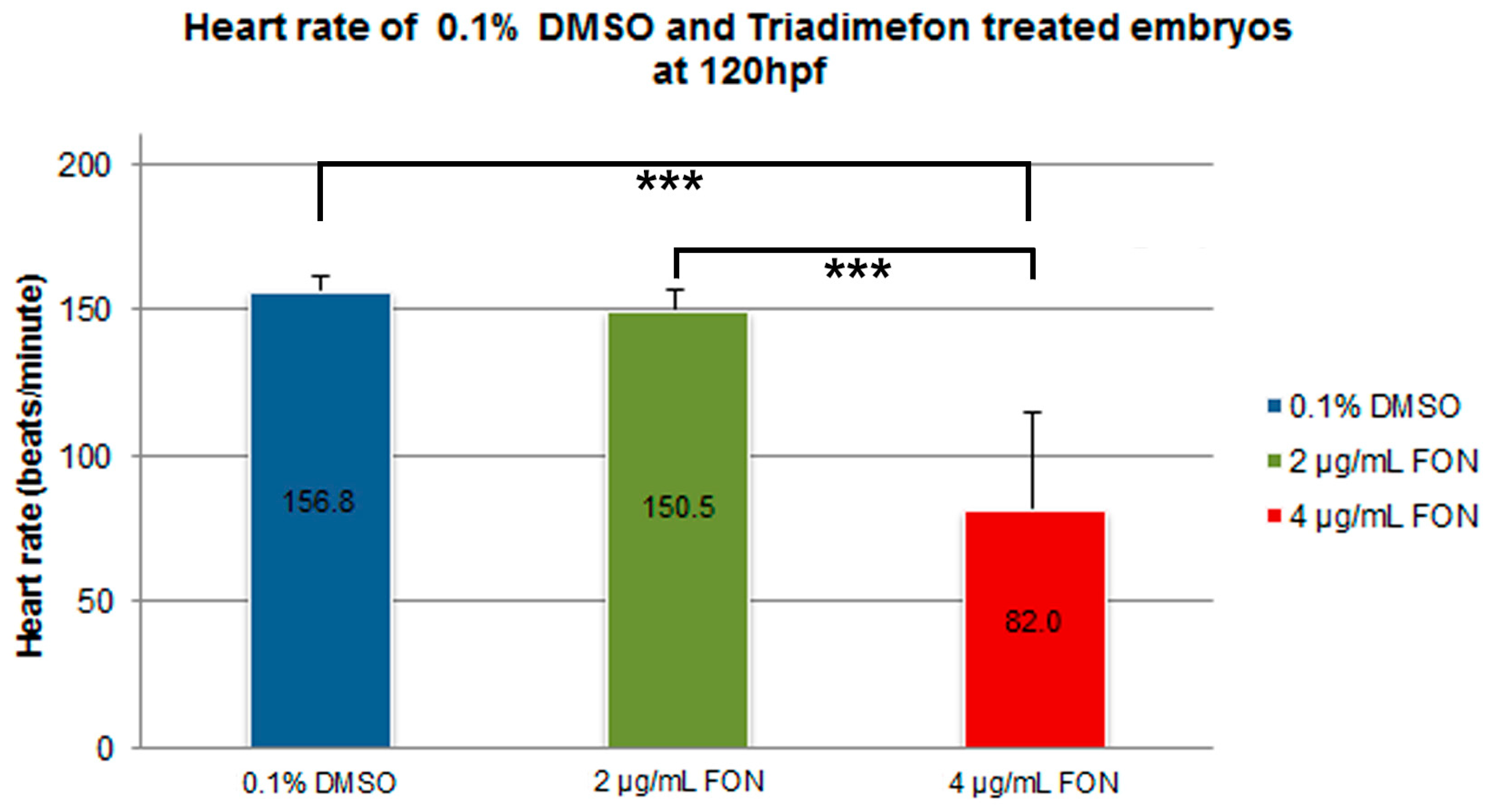
2.5. Defects in Cardiovascular Function of Triadimefon Exposed Embryos
3. Materials and Methods
3.1. Ethics Statement
3.2. Fish Husbandry and Embryo Collection
3.3. Embryo Exposure
3.4. Effect of Triadimefon on Hatching Rate and Body Length Measurement
3.5. Heart Rate Quantification
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| hpf | Hours post-fertilization |
| RA | Retinoic acid |
| WEC | Whole embryo culture |
| NCC | Neural crest cell |
References
- Marotta, F.; Tiboni, G.M. Molecular aspects of azoles-induced teratogenesis. Expert Opin. Drug Metab. Toxicol. 2010, 6, 461–482. [Google Scholar] [CrossRef] [PubMed]
- Ribas, E.R.A.D.; Spolti, P.; del Ponte, E.M.; Donato, K.Z.; Schrekker, H.; Fuentefria, A.M. Is the emergence of fungal resistance to medical triazoles related to their use in the agroecosystems? A mini review. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2016, 47, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Goetz, A.K.; Dix, D.J. Toxicogenomic effects common to triazole antifungals and conserved between rats and humans. Toxicol. Appl. Pharmacol. 2009, 238, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Hester, S.D.; Nesnow, S. Transcriptional responses in thyroid tissues from rats treated with a tumorigenic and a non-tumorigenic triazole conazole fungicide. Toxicol. Appl. Pharmacol. 2008, 227, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Goetz, A.K.; Dix, D.J. Mode of action for reproductive and hepatic toxicity inferred from a genomic study of triazole antifungals. Toxicol. Sci. 2009, 110, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Leavitt, S.A. Analysis of the mutations induced by conazole fungicides in vivo. Mutagenesis 2010, 25, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, F.; Rossi, F.; Prati, M.; Giavini, E.; Menegola, E. Early genetic control of craniofacial development is affected by the in vitro exposure of rat embryos to the fungicide triadimefon. Birth Defects Res. B Dev. Reprod. Toxicol. 2011, 92, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Ward, W.O.; Delker, D.A.; Hester, S.D.; Thai, S.F.; Wolf, D.C.; Allen, J.W.; Nesnow, S. Transcriptional profiles in liver from mice treated with hepatotumorigenic and nonhepatotumorigenic triazole conazole fungicides: Propiconazole, triadimefon, and myclobutanil. Toxicol. Pathol. 2006, 34, 863–878. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.C.; Allen, J.W.; George, M.H.; Hester, S.D.; Sun, G.; Moore, T.; Thai, S.F.; Delker, D.; Winkfield, E.; Leavitt, S.; et al. Toxicity profiles in rats treated with tumorigenic and nontumorigenic triazole conazole fungicides: Propiconazole, triadimefon, and myclobutanil. Toxicol. Pathol. 2006, 34, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Goetz, A.K.; Ren, H.; Schmid, J.E.; Blystone, C.R.; Thillainadarajah, I.; Best, D.S.; Nichols, H.P.; Strader, L.F.; Wolf, D.C.; Narotsky, M.G.; et al. Disruption of testosterone homeostasis as a mode of action for the reproductive toxicity of triazole fungicides in the male rat. Toxicol. Sci. 2007, 95, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Walker, Q.D.; Mailman, R.B. Triadimefon and triadimenol: Effects on monoamine uptake and release. Toxicol. Appl. Pharmacol. 1996, 139, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Menegola, E.; Broccia, M.L.; di Renzo, F.; Massa, V.; Giavini, E. Craniofacial and axial skeletal defects induced by the fungicide triadimefon in the mouse. Birth Defects Res. B Dev. Reprod. Toxicol. 2005, 74, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, F.; Broccia, M.L.; Giavini, E.; Menegola, E. Antifungal triazole derivative triadimefon induces ectopic maxillary cartilage by altering the morphogenesis of the first branchial arch. Birth Defects Res. B Dev. Reprod. Toxicol. 2006, 80, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, F.; Broccia, M.L.; Giavini, E.; Menegola, E. Stage-dependent abnormalities induced by the fungicide triadimefon in the mouse. Reprod. Toxicol. 2011, 31, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Morriss-Kay, G. Retinoic acid and craniofacial development: Molecules and morphogenesis. Bioessays 1993, 15, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Groppelli, S.; Pennati, R.; De Bernardi, F.; Menegola, E.; Giavini, E.; Sotgia, C. Teratogenic effects of two antifungal triazoles, triadimefon and triadimenol, on Xenopus laevis development: Craniofacial defects. Aquat. Toxicol. 2005, 73, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Pennati, R.; Groppelli, S.; Zega, G.; Biggiogero, M.; De Bernardi, F.; Sotgia, C. Toxic effects of two pesticides, Imazalil and Triadimefon, on the early development of the ascidian Phallusia mammillata (Chordata, Ascidiacea). Aquat. Toxicol. 2006, 79, 205–212. [Google Scholar] [CrossRef] [PubMed]
- McCollum, C.W.; Ducharme, N.A.; Bondesson, M.; Gustafsson, J.A. Developmental toxicity screening in zebrafish. Birth Defects Res. C Embryo Today 2011, 93, 67–114. [Google Scholar] [CrossRef] [PubMed]
- Veldman, M.B.; Lin, S. Zebrafish as a developmental model organism for pediatric research. Pediatr. Res. 2008, 64, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chang, J.; Zhu, G. Developmental Toxicity of Triadimefon in Embryo-Larval Stages of Zebrafish. In Proceedings of the 2010 International Conference on Digital Manufacturing and Automation, Changsha, China, 18–20 December 2010; Volume 1, pp. 552–555. [Google Scholar]
- He, J.H.; Gao, J.M.; Huang, C.J.; Li, C.Q. Zebrafish models for assessing developmental and reproductive toxicity. Neurotoxicol. Teratol. 2014, 42, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Hussain, S.; Bano, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.A.; Khan, F.; Chen, Y.; Wu, C.; et al. Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: Consequences for changing environment. Environ. Sci. Pollut. Res. Int. 2015, 22, 4907–4921. [Google Scholar] [CrossRef] [PubMed]
- Sylvain, N.J.; Brewster, D.L.; Ali, D.W. Zebrafish embryos exposed to alcohol undergo abnormal development of motor neurons and muscle fibers. Neurotoxicol. Teratol. 2010, 32, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.J.; Seok, S.H.; Baek, M.W.; Lee, H.Y.; Na, Y.R.; Park, S.H.; Lee, H.K.; Dutta, N.K.; Kawakami, K.; Park, J.H. Benomyl induction of brain aromatase and toxic effects in the zebrafish embryo. J. Appl. Toxicol. 2009, 29, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.L.; Carten, J.D.; Farber, S.A. Zebrafish lipid metabolism: From mediating early patterning to the metabolism of dietary fat and cholesterol. Methods Cell Biol. 2011, 101, 111–141. [Google Scholar] [PubMed]
- Jomaa, B.; Hermsen, S.A.; Kessels, M.Y.; van den Berg, J.H.; Peijnenburg, A.A.; Aarts, J.M.; Piersma, A.H.; Rietjens, I.M. Developmental toxicity of thyroid-active compounds in a zebrafish embryotoxicity test. ALTEX 2014, 31, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Holtta-Vuori, M.; Salo, V.T.; Nyberg, L.; Brackmann, C.; Enejder, A.; Panula, P.; Ikonen, E. Zebrafish: Gaining popularity in lipid research. Biochem. J. 2010, 429, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Hermsen, S.A.; Pronk, T.E.; van den Brandhof, E.J.; van der Ven, L.T.; Piersma, A.H. Concentration-response analysis of differential gene expression in the zebrafish embryotoxicity test following flusilazole exposure. Toxicol. Sci. 2012, 127, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Hermsen, S.A.; Pronk, T.E.; van den Brandhof, E.J.; van der Ven, L.T.; Piersma, A.H. Triazole-induced gene expression changes in the zebrafish embryo. Reprod. Toxicol. 2012, 34, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Jugan, M.L.; Levi, Y.; Blondeau, J.P. Endocrine disruptors and thyroid hormone physiology. Biochem. Pharmacol. 2010, 79, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chang, J.; Zhao, Y.; Zhu, G. Changes of thyroid hormone levels and related gene expression in zebrafish on early life stage exposure to triadimefon. Environ. Toxicol. Pharmacol. 2011, 32, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wang, Z.; Collins, J.E.; Andrews, R.M.; Stemple, D.; Gong, Z. Comparative transcriptome analyses indicate molecular homology of zebrafish swimbladder and mammalian lung. PLoS ONE 2011, 6, e24019. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.N.; McGee, C.A.; Dumbarton, T.C.; Croll, R.P.; Smith, F.M. Development of the swimbladder and its innervation in the zebrafish, Danio rerio. J. Morphol. 2007, 268, 967–985. [Google Scholar] [CrossRef] [PubMed]
- Zoupa, M. Normal Triadimefon Phenotypes. Ph.D. Thesis, Benaki Phytopathological Institute, Attika, Greece, 2016. [Google Scholar]
- Holley, S.A. The genetics and embryology of zebrafish metamerism. Dev. Dyn. 2007, 236, 1422–1449. [Google Scholar] [CrossRef] [PubMed]
- Stickney, H.L.; Barresi, M.J.; Devoto, S.H. Somite development in zebrafish. Dev. Dyn. 2000, 219, 287–303. [Google Scholar] [CrossRef]
- Lenkowski, J.R.; Sanchez-Bravo, G.; McLaughlin, K.A. Low concentrations of atrazine, glyphosate, 2,4-dichlorophenoxyacetic acid, and triadimefon exposures have diverse effects on Xenopus laevis organ morphogenesis. J. Environ. Sci. 2010, 22, 1305–1308. [Google Scholar] [CrossRef]
- Papis, E.; Bernardini, G.; Gornati, R.; Menegola, E.; Prati, M. Gene expression in Xenopus laevis embryos after Triadimefon exposure. Gene Expr. Patterns 2007, 7, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Van der Meer, D.L.; Marques, I.J.; Leito, J.T.; Besser, J.; Bakkers, J.; Schoonheere, E.; Bagowski, C.P. Zebrafish cypher is important for somite formation and heart development. Dev. Biol. 2006, 299, 356–372. [Google Scholar] [CrossRef] [PubMed]
- Van Eeden, F.J.; Granato, M.; Schach, U.; Brand, M.; Furutani-Seiki, M.; Haffter, P.; Hammerschmidt, M.; Heisenberg, C.P.; Jiang, Y.J.; Kane, D.A.; et al. Mutations affecting somite formation and patterning in the zebrafish, Danio rerio. Development 1996, 123, 153–164. [Google Scholar] [PubMed]
- Julich, D.; Geisler, R.; Holley, S.A. Integrinα5 and delta/notch signaling have complementary spatiotemporal requirements during zebrafish somitogenesis. Dev. Cell 2005, 8, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Koshida, S.; Kishimoto, Y.; Ustumi, H.; Shimizu, T.; Furutani-Seiki, M.; Kondoh, H.; Takada, S. Integrinα5-dependent fibronectin accumulation for maintenance of somite boundaries in zebrafish embryos. Dev. Cell 2005, 8, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Giavini, E.; Menegola, E. Are azole fungicides a teratogenic risk for human conceptus? Toxicol. Lett. 2010, 198, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Menegola, E.; di Renzo, F.; Broccia, M.L. Cleft palate and exposure to an azole derivative: Triadimefon. Reprod. Toxicol. 2009, 28, 123. [Google Scholar] [CrossRef]
- Kazy, Z.; Puho, E.; Czeizel, A.E. Teratogenic potential of vaginal metronidazole treatment during pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2005, 123, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Czeizel, A.E.; Rockenbauer, M. A population based case-control teratologic study of oral metronidazole treatment during pregnancy. Br. J. Obstet. Gynaecol. 1998, 105, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Machera, K. Developmental toxicity of cyproconazole, an inhibitor of fungal ergosterol biosynthesis, in the rat. Bull. Environ. Contam. Toxicol. 1995, 54, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Menegola, E.; Di Renzo, F.; Metruccio, F.; Moretto, A.; Giavini, E. Effects of mixtures of azole fungicides in postimplantation rat whole-embryo cultures. Arch. Toxicol. 2013, 87, 1989–1997. [Google Scholar] [CrossRef] [PubMed]
- Menegola, E.; Broccia, M.L.; Di Renzo, F.; Prati, M.; Giavini, E. In vitro teratogenic potential of two antifungal triazoles: Triadimefon and triadimenol. In Vitro Cell. Dev. Biol. Anim. 2000, 36, 88–95. [Google Scholar] [CrossRef]
- Menegola, E.; Broccia, M.L.; Di Renzo, F.; Giavini, E. Antifungal triazoles induce malformations in vitro. Reprod. Toxicol. 2001, 15, 421–427. [Google Scholar] [CrossRef]
- Menegola, E.; Broccia, M.L.; Di Renzo, F.; Giavini, E. Dysmorphogenic effects of some fungicides derived from the imidazole on rat embryos cultured in vitro. Reprod. Toxicol. 2006, 21, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Ito, Y.; Han, J. TGF-β signaling and its functional significance in regulating the fate of cranial neural crest cells. Crit. Rev. Oral Biol. Med. 2003, 14, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Sanford, L.P.; Ormsby, I.; Gittenberger-de Groot, A.C.; Sariola, H.; Friedman, R.; Boivin, G.P.; Cardell, E.L.; Doetschman, T. TGFβ2 knockout mice have multiple developmental defects that are non-overlapping with other TGFβ knockout phenotypes. Development 1997, 124, 2659–2670. [Google Scholar] [PubMed]
- Di Renzo, F.; Corsini, E.; Broccia, M.L.; Marinovich, M.; Galli, C.L.; Giavini, E.; Menegola, E. Molecular mechanism of teratogenic effects induced by the fungicide triadimefon: Study of the expression of TGF-β mRNA and TGF-β and CRABPI proteins during rat in vitro development. Toxicol. Appl. Pharmacol. 2009, 234, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Van Wauwe, J.P.; Coene, M.C.; Goossens, J.; Cools, W.; Monbaliu, J. Effects of cytochrome P-450 inhibitors on the in vivo metabolism of all-trans-retinoic acid in rats. J. Pharmacol. Exp. Ther. 1990, 252, 365–369. [Google Scholar] [PubMed]
- Vanier, K.L.; Mattiussi, A.J.; Johnston, D.L. Interaction of all-trans-retinoic acid with fluconazole in acute promyelocytic leukemia. J. Pediatr. Hematol. Oncol. 2003, 25, 403–404. [Google Scholar] [CrossRef] [PubMed]
- Menegola, E.; Broccia, M.L.; Di Renzo, F.; Massa, V.; Giavini, E. Relationship between hindbrain segmentation, neural crest cell migration and branchial arch abnormalities in rat embryos exposed to fluconazole and retinoic acid in vitro. Reprod. Toxicol. 2004, 18, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Papis, E.; Bernardini, G.; Gornati, R.; Prati, M. Triadimefon causes branchial arch malformations in Xenopus laevis embryos. Environ. Sci. Pollut. Res. Int. 2006, 13, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Carter, T.C.; Druschel, C.M.; Romitti, P.A.; Bell, E.M.; Werler, M.M.; Mitchell, A.A. Antifungal drugs and the risk of selected birth defects. Am. J. Obstet. Gynecol. 2008, 198, 191.e1–191.e7. [Google Scholar] [CrossRef] [PubMed]
- Pursley, T.J.; Blomquist, I.K.; Abraham, J.; Andersen, H.F.; Bartley, J.A. Fluconazole-induced congenital anomalies in three infants. Clin. Infect. Dis. 1996, 22, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Van den Bulck, K.; Hill, A.; Mesens, N.; Diekman, H.; De Schaepdrijver, L.; Lammens, L. Zebrafish developmental toxicity assay: A fishy solution to reproductive toxicity screening, or just a red herring? Reprod. Toxicol. 2011, 32, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, J.V.; McArthur, A.G.; Kubota, A.; Zanette, J.; Parente, T.; Jonsson, M.E.; Nelson, D.R.; Stegeman, J.J. Identification and developmental expression of the full complement of Cytochrome P450 genes in Zebrafish. BMC Genom. 2010, 11, 643. [Google Scholar] [CrossRef] [PubMed]
- Tiboni, G.M.; Giampietro, F. Murine teratology of fluconazole: Evaluation of developmental phase specificity and dose dependence. Pediatr. Res. 2005, 58, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Tiboni, G.M.; Marotta, F.; Castigliego, A.P. Teratogenic effects in mouse fetuses subjected to the concurrent in utero exposure to miconazole and metronidazole. Reprod. Toxicol. 2008, 26, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Carey, J.C.; Martinez, L.; Balken, E.; Leen-Mitchell, M.; Robertson, J. Determination of human teratogenicity by the astute clinician method: Review of illustrative agents and a proposal of guidelines. Birth Defects Res. A Clin. Mol. Teratol. 2009, 85, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Farag, A.T.; Ibrahim, H.H. Developmental toxic effects of antifungal flusilazole administered by gavage to mice. Birth Defects Res. B Dev. Reprod. Toxicol. 2007, 80, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Tiboni, G.M.; Marotta, F.; Del Corso, A.; Giampietro, F. Defining critical periods for itraconazole-induced cleft palate, limb defects and axial skeletal malformations in the mouse. Toxicol. Lett. 2006, 167, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Amaral, V.C.; Nunes, G.P., Jr. Prednisone reduces ketoconazole-induced skeletal defects in rat fetuses. Arch. Toxicol. 2009, 83, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Menegola, E.; Broccia, M.L.; Di Renzo, F.; Massa, V.; Giavini, E. Study on the common teratogenic pathway elicited by the fungicides triazole-derivatives. Toxicol. In Vitro 2005, 19, 737–748. [Google Scholar] [CrossRef] [PubMed]
- De Jong, E.; Barenys, M.; Hermsen, S.A.; Verhoef, A.; Ossendorp, B.C.; Bessems, J.G.; Piersma, A.H. Comparison of the mouse Embryonic Stem cell Test, the rat Whole Embryo Culture and the Zebrafish Embryotoxicity Test as alternative methods for developmental toxicity testing of six 1,2,4-triazoles. Toxicol. Appl. Pharmacol. 2011, 253, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, F.; Broccia, M.L.; Giavini, E.; Menegola, E. Citral, an inhibitor of retinoic acid synthesis, attenuates the frequency and severity of branchial arch abnormalities induced by triazole-derivative fluconazole in rat embryos cultured in vitro. Reprod. Toxicol. 2007, 24, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio Rerio), 5th ed.; University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Hermsen, S.A.; van den Brandhof, E.J.; van der Ven, L.T.; Piersma, A.H. Relative embryotoxicity of two classes of chemicals in a modified zebrafish embryotoxicity test and comparison with their in vivo potencies. Toxicol. In Vitro 2011, 25, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Hoage, T.; Ding, Y.; Xu, X. Quantifying cardiac functions in embryonic and adult zebrafish. Methods Mol. Biol. 2012, 843, 11–20. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotype | 72 hpf | 96 hpf | 120 hpf | |||
|---|---|---|---|---|---|---|
| 2 µg/mL | 4 µg/mL | 2 µg/mL | 4 µg/mL | 2 µg/mL | 4 µg/mL | |
| Hatching | -- | -- | delayed | delayed | delayed | delayed |
| Length | na | na | na | na | N | shorter |
| Spine shape | N | N | N | N | N | bent |
| Yolk sac | N | N | minor enlargement | enlarged | minor enlargement | enlarged/misshaped |
| Swim bladder | N | N | minor inhibition | minor inhibition | uniflated | uniflated |
| Somites | N | N | N | N | N | severely affected |
| Horizontal myoseptum | N | N | N | N | N | reduced |
| Mandible | N | N | delayed | hypoplastic | severely hypoplastic | severely hypo plastic/absent |
| Maxilla | N | N | minor delay | minor delay | minor delay | minor delay |
| Lower pharyngeal arches | N | N | N | possible delay | slightly hypoplastic | severely hypo plastic/absent |
| Neurocranim | N | N | flattened forebrain | flattened forebrain | mild delay of midbrain/hindbrain | severely reduced size/flattened forebrain hypolastic mid-brain/ no clear forebrain mid-brain boundary mild delay of hindbrain cleft of primary neuronal divisions |
| Heart shape | N | N | N | linear/delayed | N | massive pericardial edemas severe decrease in size linear shape |
| Heart rate | na | na | na | na | N | brachycardia |
| Blood circulation | N | N | N | N | N | severely decrease/absent |
| Phenotype (Identified/Published) | Zebrafish | Mammalian and Other In Vivo Models | In Vitro Models/Embryo Cultures | Human Studies |
|---|---|---|---|---|
| Hatching | Inhibited/delayed: could be a result of decrease movement activity due to somitic abnormalities | |||
| Body length | Shorter: may arise from skeletal defects, irregularities in somitic boundaries and disorganized muscle fibers | Malformations at the level of axial skeleton (FON) [13] Fusions, duplications or morphological transformations of mouse axial segments (FON) [12] Reduced humeral length in mouse (FLUC) [63] Axial skeletal defects in mouse (MIC and MET) [64] Reduced body length in ascidian (IM) [17] | Skeletal defects (FLUC) [60] Skeletal manifestations of humeral radial synostosis and femoral bowing (FLUC) [65] Limb defects (IT) | |
| Spine | Bent: phenotype associated with somitic defects that may lead to muscle and skeletal phenotypes | Axial skeletal and limb defects in mouse (FLUS) [66] Axial skeletal defects in mouse (IT) [67] Limb anomalies in rat (KET) [68] Bent spine in ascidian (IM) [17] Axial defects (homeotic respe-cification/lumbar rib) (FON) [44] Short tail in ascidian (IM, FON) [17] | ||
| Yolk sac | Enlarged/misshaped: associated with lipid metabolism defects | |||
| Swim bladder | Uninflated: secondary effects of the curved spine/muscle/skeletal phenotype, a delay in hatching that didn’t allow inflation to occur on time; reduced movement as a result of somitic defects | |||
| Somites | Irregular somitic formation, disorganized muscle fibers, hypolastic horizontal myoseptum | Somite segmentation defects in X. laevis (FON, NOL) [16] | Fusion of the I and II branchial arches [7] Romboencephalic cleft, abnormal somites (FON) [49] Abnormal somitic develop-ment (KET and EN) [51] | |
| Upper branchial arches (facial structures/mandible/maxillae) | Severely hypoplastic or even absent, Minor delay in development | Micrognathia, microtia, short/fused mandible with zygomatic, joined mandible and maxillae, malformed I and II branchial arches (FON) [13] Abnormal shape, fusions, and agenesis of craniofacial structures originated from branchial ectomesenchyme (palatine, basisphenoid, alisphenoid, pterygoid, squamosal, zygomatic, maxilla, mandible, Meckel’s cartilage, tympanic ring, ear ossicles) during mouse development (FON) [12] Absence of mandibular and maxillary cartilages in X. laevis (FON) [16] Absence of mandibular cartilage and fusion of mandibular cartilage with adjacent structures in X. laevis (NOL) [16] Craniofacial defects in mice (the migration of encephalic neural crest cells) (FON) [44] | Branchial apparatus * reduction/absence, increased cell death of branchial mesenchyme (FLUC and FLU) [50] I branchial arch reduction/I and II branchial arch fusion (FLUC) [57] Reduction of I branchial arch, absence of II branchial arch (FON) [49] Dorso-ventral reduction of I and II branchial arches (NOL) [49] Fusion between I and II branchial arches (FLU, FON and NOL) [69] II branchial arch reduction, I/II branchial arch fusion (KET) [51] I and II branchial arch reduction and fusion (EN) [51] Fusion of the I and II branchial arches (FON) [7] Reduction/ fusion of I and II branchial arches (FON, HEX, FLUS, CYP, MYC, TRI) [70] | |
| Lower branchial arches | Severely hypoplastic hypoplastic or even absent lower pharyngeal arches | Altered morphogenesis of the branchial apparatus (FLUC) [71] II/III branchial arch fusion (KET) [51] | ||
| Clefts | Cleft palate in rat (FLUC, FON, CYP, KET) [12,47,63,68] Cleft palate in mouse (FLUC, IT, FON) [44,45,63,66] | Cleft forebrain and midbrain (KET) [51] | Cleft palate (FLUC) [60,65] Oral cleft (MIC, TER and KET) [59] | |
| Neurocranium | Reduced size/flat forebrain, hypoplastic midbrain, with no clear forebrain midbrain boundaries | Short anterio-dorsal cranial region, slight mouth protrusion of X. laevis (FON, NOL) [16] Alterations of anterior end trunk in ascidian (FON) [17] Abnormal neural crest cell migration from hindbrain to the branchial arches of mice (FLUC) [57] | Reduction of prosencephalon, encephalic schisis (FON) [49] Alterations of hindbrain segmentation (FLU, FON and NOL) [69] Asymmetric forebrain and hindbrain (IMI) [51] Microcephaly, asymmetric forebrain, reduced hindbrain (KET) [51] Reduced, swollen, asymmetric hindbrain (EN) [51] | Craniosynostosis, brachycephaly (FLUC) [60] Craniosynostosis (MIC, TER and KET) [59] Craniosynostosis (FLUC) [65] Craniofacial ossification defects, hypolastic facial bones, small face, brachy-cephaly (FLUC) [60] Microcephalia (IT) |
| Cardiovascular | Pericardial edema, decrease of heart size, linear heart shape/brachycardia, severely decreased or even absent circulation | Cardiac edema of X. laevis (FON, NOL) [16] | Tetralogy of fallot, ventricular septal defects, pulmonary artery hypoplasia (FLUC) [60] Hypoplastic left heart (MIC, TER and KET) [59] Several cardiovascular malformations (FLUC) [65] Hypoplastic left heart syndrome (IT) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoupa, M.; Machera, K. Zebrafish as an Alternative Vertebrate Model for Investigating Developmental Toxicity—The Triadimefon Example. Int. J. Mol. Sci. 2017, 18, 817. https://doi.org/10.3390/ijms18040817
Zoupa M, Machera K. Zebrafish as an Alternative Vertebrate Model for Investigating Developmental Toxicity—The Triadimefon Example. International Journal of Molecular Sciences. 2017; 18(4):817. https://doi.org/10.3390/ijms18040817
Chicago/Turabian StyleZoupa, Maria, and Kyriaki Machera. 2017. "Zebrafish as an Alternative Vertebrate Model for Investigating Developmental Toxicity—The Triadimefon Example" International Journal of Molecular Sciences 18, no. 4: 817. https://doi.org/10.3390/ijms18040817
APA StyleZoupa, M., & Machera, K. (2017). Zebrafish as an Alternative Vertebrate Model for Investigating Developmental Toxicity—The Triadimefon Example. International Journal of Molecular Sciences, 18(4), 817. https://doi.org/10.3390/ijms18040817