
Genome-Wide Analyses of MicroRNA Profiling in Interleukin-27 Treated Monocyte-Derived Human Dendritic Cells Using Deep Sequencing: A Pilot Study


 and
and
Abstract
:
1. Introduction
2. Results
2.1. miRNA Library Analysis
2.2. Highly Expressed miRNAs in DCs
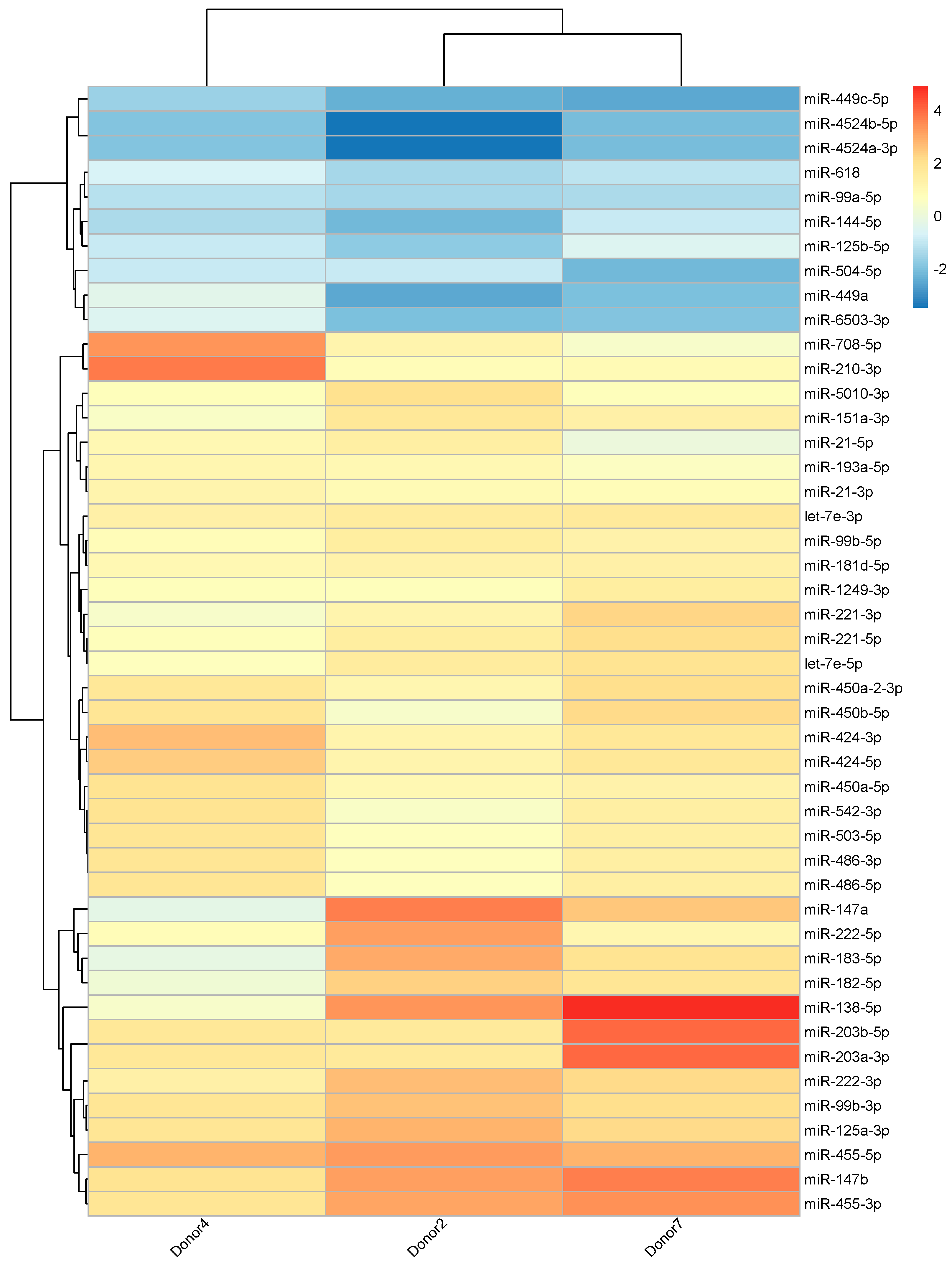
2.3. Differential miRNA Expression in 27DCs
2.4. KEGG Pathway Enrichment Analysis
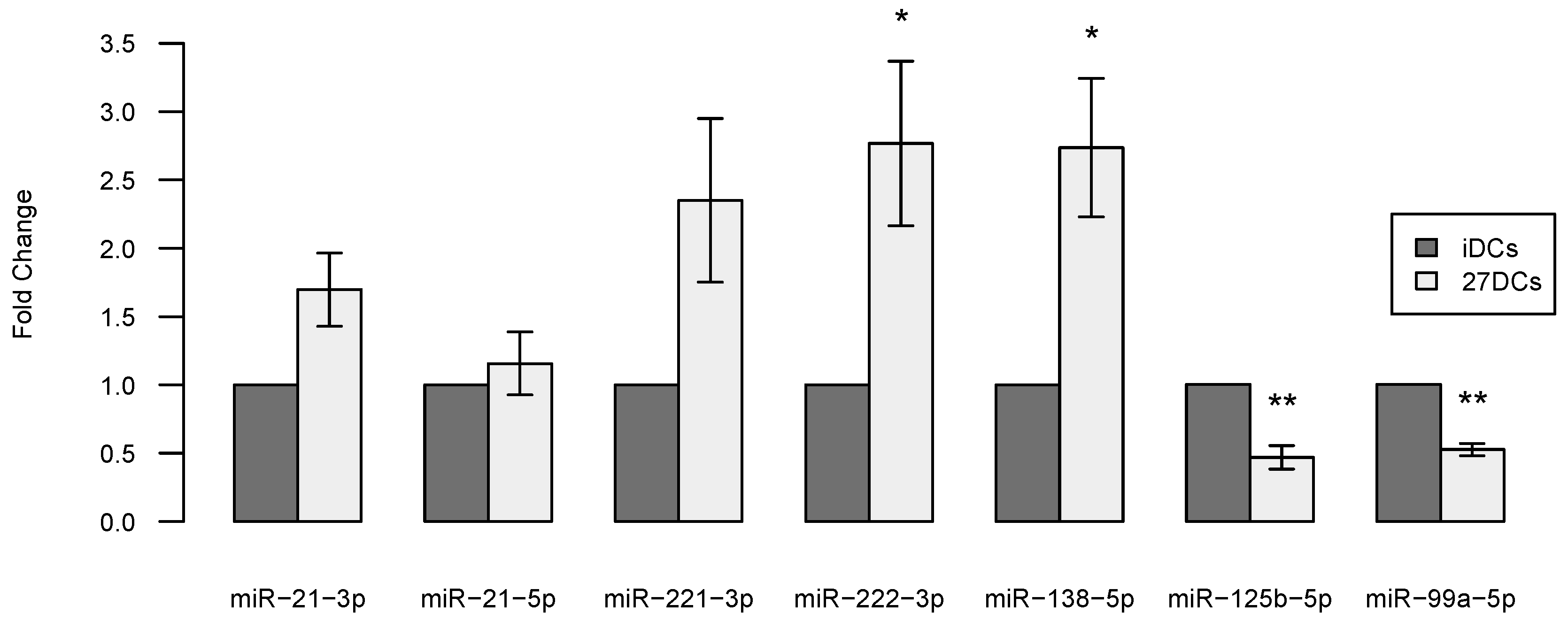
2.5. Validation of Differential miRNAs by RT-qPCR
2.6. Potential Viruses Targeted by Differential miRNAs
2.7. Novel miRNA Discovery
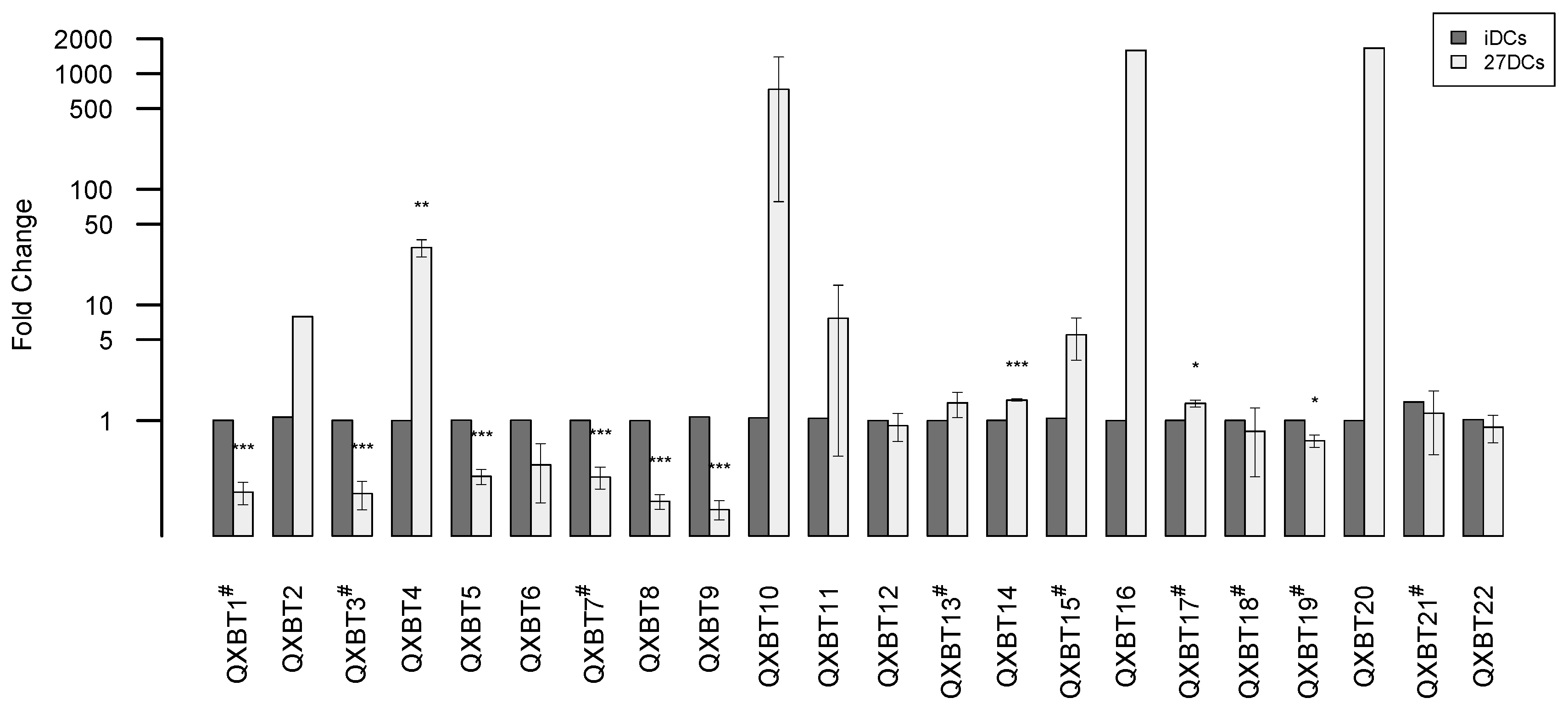
2.8. Validation of Novel miRNAs
3. Discussion
4. Materials and Methods
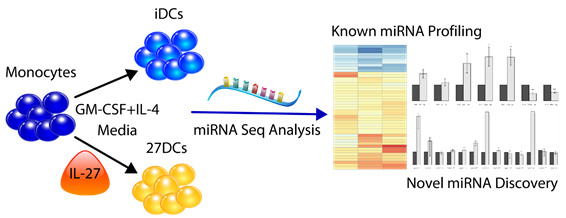
4.1. Generation of DC Subtypes
4.2. miRNA Library Preparation and Sequencing
4.3. Differential Expression Analysis
4.4. KEGG Pathway and GO Term Enrichment Analysis
4.5. Prediction of Viruses Targeted by miRNAs
4.6. Discovery of Candidate Novel miRNAs
4.7. Quantitative RT-PCR for miRNA Validation
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 27DCs | Monocyte-derived IL-27 treated immature dendritic cells |
| APCs | Antigen presenting cells |
| DCs | Dendritic cells |
| EBI-3 | Epstein-Barr virus-induced gene 3 |
| FDR | False discovery rate |
| GO | Gene ontology |
| iDCs | Monocyte-derived immature DCs |
| IL-27 | Interleukin-27 |
| JAK/STAT | Janus kinase/signal transducer and activator of transcription |
| JNK | C-Jun N-terminal kinases |
| KEGG | Kyoto encyclopedia of genes and genomes |
| logCPM | log2 counts per million |
| MAPKs | Mitogen-activated protein kinases |
| MFE | Minimum free energy |
| miRNAs | MicroRNAs |
| PI3K | Phosphoinositide 3-kinase |
| piRNAs | Piwi-interacting RNAs |
| RT-qPCR | Quantitative reverse transcription polymerase chain reaction |
| TLR | toll-like receptors |
References
- Liu, K.; Nussenzweig, M.C. Origin and development of dendritic cells. Immunol. Rev. 2010, 234, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of dendritic cells. Ann. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; KewalRamani, V.N. Dendritic-cell interactions with HIV: Infection and viral dissemination. Nat. Rev. Immunol. 2006, 6, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.M.; Wu, L. HIV interactions with monocytes and dendritic cells: Viral latency and reservoirs. Retrovirology 2009, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Kim, V.N. MicroRNA biogenesis: Coordinated cropping and dicing. Nat. Rev. Mol. Cell Biol. 2005, 6, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, C.C.; Cheng, H.H.; Tewari, M. MicroRNA profiling: Approaches and considerations. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Nishida, N.; Calin, G.A.; Pantel, K. Clinical relevance of circulating cell-free microRNAs in cancer. Nat. Rev. Clin. Oncol. 2014, 11, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.L.; Schnorfeil, F.M.; Brocker, T. MicroRNAs regulate dendritic cell differentiation and function. J. Immunol. 2011, 187, 3911–3917. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z.; Schaffert, S.; Fragoso, R.; Loh, C. Regulation of immune responses and tolerance: The microRNA perspective. Immunol. Rev. 2013, 253, 112–128. [Google Scholar] [CrossRef] [PubMed]
- Montecalvo, A.; Larregina, A.T.; Shufesky, W.J.; Stolz, D.B.; Sullivan, M.L.; Karlsson, J.M.; Baty, C.J.; Gibson, G.A.; Erdos, G.; Morelli, A.E.; et al. Mechanism of transfer of functional microRNAs between mouse dendritic cells via exosomes. Blood 2012, 119, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Mittelbrunn, M.; Gutierrez-Vazquez, C.; Villarroya-Beltri, C.; Gonzalez, S.; Sanchez-Cabo, F.; Gonzalez, M.A.; Bernad, A.; Sanchez-Madrid, F. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat. Commun. 2011, 2, 282. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, G.; Navas-Martin, S.; Martin-Garcia, J. MicroRNAs and HIV-1 infection: Antiviral activities and beyond. J. Mol. Biol. 2014, 426, 1178–1197. [Google Scholar] [CrossRef] [PubMed]
- Stumhofer, J.S.; Hunter, C.A. Advances in understanding the anti-inflammatory properties of IL-27. Immunol. Lett. 2008, 117, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Pflanz, S.; Timans, J.C.; Cheung, J.; Rosales, R.; Kanzler, H.; Gilbert, J.; Hibbert, L.; Churakova, T.; Travis, M.; Kastelein, R.A.; et al. IL-27, a heterodimeric cytokine composed of EBI3 and p28 protein, induces proliferation of naive CD4+ T cells. Immunity 2002, 16, 779–790. [Google Scholar] [CrossRef]
- Lucas, S.; Ghilardi, N.; Li, J.; de Sauvage, F.J. IL-27 regulates IL-12 responsiveness of naive CD4+ T cells through Stat1-dependent and -independent mechanisms. Proc. Natl. Acad. Sci. USA 2003, 100, 15047–15052. [Google Scholar] [CrossRef] [PubMed]
- Hibbert, L.; Pflanz, S.; de Waal Malefyt, R.; Kastelein, R.A. IL-27 and IFN-α signal via Stat1 and Stat3 and induce T-Bet and IL-12Rβα2 in naive T cells. J. Interferon Cytokine Res. 2003, 23, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Vignali, D.A.; Kuchroo, V.K. IL-12 family cytokines: Immunological playmakers. Nat. Immunol. 2012, 13, 722–778. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Swaminathan, S.; Yang, D.; Dai, L.; Sui, H.; Yang, J.; Hornung, R.L.; Wang, Y.; Hu, X.; Imamichi, T.; et al. Interleukin-27 is a potent inhibitor of cis HIV-1 replication in monocyte-derived dendritic cells via a type I interferon-independent pathway. PLoS ONE 2013, 8, e59194. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Lidie, K.B.; Chen, Q.; Adelsberger, J.W.; Zheng, X.; Huang, D.; Yang, J.; Lempicki, R.A.; Lane, H.C.; Imamichi, T.; et al. IL-27 inhibits HIV-1 infection in human macrophages by down-regulating host factor SPTBN1 during monocyte to macrophage differentiation. J. Exp. Med. 2013, 210, 517–534. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Dilger, P.; Bird, C.; Wadhwa, M. IL-27 Promotes proliferation of human leukemic cell lines through the MAPK/ERK signaling pathway and suppresses sensitivity to chemotherapeutic drugs. J. Interferon Cytokine Res. 2016, 36, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Diegelmann, J.; Olszak, T.; Goke, B.; Blumberg, R.S.; Brand, S. A novel role for interleukin-27 (IL-27) as mediator of intestinal epithelial barrier protection mediated via differential signal transducer and activator of transcription (STAT) protein signaling and induction of antibacterial and anti-inflammatory proteins. J. Biol. Chem. 2012, 287, 286–298. [Google Scholar] [PubMed]
- Sharma, G.; Dutta, R.K.; Khan, M.A.; Ishaq, M.; Sharma, K.; Malhotra, H.; Majumdar, S. IL-27 inhibits IFN-γ induced autophagy by concomitant induction of JAK/PI3 K/Akt/mTOR cascade and up-regulation of Mcl-1 in Mycobacterium tuberculosis H37Rv infected macrophages. Int. J. Biochem. Cell Biol. 2014, 55, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Fakruddin, J.M.; Lempicki, R.A.; Gorelick, R.J.; Yang, J.; Adelsberger, J.W.; Garcia-Pineres, A.J.; Pinto, L.A.; Lane, H.C.; Imamichi, T. Noninfectious papilloma virus-like particles inhibit HIV-1 replication: Implications for immune control of HIV-1 infection by IL-27. Blood 2007, 109, 1841–1849. [Google Scholar] [CrossRef] [PubMed]
- Sowrirajan, B.; Saito, Y.; Poudyal, D.; Chen, Q.; Sui, H.; DeRavin, S.S.; Imamichi, H.; Malech, H.L.; Lane, H.L.; Imamichi, T.; et al. Interleukin-27 Enhances the potential of reactive oxygen species generation from monocyte-derived macrophages and dendritic cells by induction of p47phox. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Kostoulas, N.; Vergoulis, T.; Georgakilas, G.; Reczko, M.; Maragkakis, M.; Paraskevopoulou, M.D.; Prionidis, K.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA miRPath v.2.0: Investigating the combinatorial effect of microRNAs in pathways. Nucleic Acids Res. 2012, 40, W498–W504. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, A.E.; Li, Q.; Xie, L.; Xie, J. Biology of IL-27 and its role in the host immunity against Mycobacterium tuberculosis. Int. J. Biol. Sci. 2015, 11, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, S.; Owaki, T.; Morishima, N.; Fukai, F.; Mizuguchi, J.; Yoshimoto, T. An indispensable role for STAT1 in IL-27-induced T-bet expression but not proliferation of naive CD4+ T cells. J. Immunol. 2004, 173, 3871–3877. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Hamano, S.; Yamanaka, A.; Hanada, T.; Ishibashi, T.; Mak, T.W.; Yoshimura, A.; Yoshida, H. Cutting edge: Role of IL-27/WSX-1 signaling for induction of T-bet through activation of STAT1 during initial Th1 commitment. J. Immunol. 2003, 170, 4886–4890. [Google Scholar] [CrossRef] [PubMed]
- Blahoianu, M.A.; Rahimi, A.A.; Kozlowski, M.; Angel, J.B.; Kumar, A. IFN-γ-induced IL-27 and IL-27p28 expression are differentially regulated through JNK MAPK and PI3K pathways independent of Jak/STAT in human monocytic cells. Immunobiology 2014, 219, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Heikkilä, O.; Nygårdas, M.; Paavilainen, H.; Ryödi, E.; Hukkanen, V. Interleukin-27 inhibits herpes simplex virus type 1 infection by activating STAT1 and 3, interleukin-6, and chemokines IP-10 and MIG. J. Interferon Cytokine Res. 2016, 36, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, S.; Hu, X.; Zheng, X.; Kriga, Y.; Shetty, J.; Zhao, Y.; Stephens, R.; Tran, B.; Lane, H.C.; Imamichi, T.; et al. Interleukin-27 treated human macrophages induce the expression of novel microRNAs which may mediate anti-viral properties. Biochem. Biophys. Res. Commun. 2013, 434, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Lecellier, C.H.; Dunoyer, P.; Arar, K.; Lehmann-Che, J.; Eyquem, S.; Himber, C.; Saib, A.; Voinnet, O. A cellular microRNA mediates antiviral defense in human cells. Science 2005, 308, 557–560. [Google Scholar] [CrossRef] [PubMed]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human microRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Huang, X.; Lu, C.; Cairo, M.S.; Zhou, X. MicroRNA-146a and microRNA-146b regulate human dendritic cell apoptosis and cytokine production by targeting TRAF6 and IRAK1 proteins. J. Biol. Chem. 2015, 290, 2831–2841. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, F.; Jia, H.; Zhang, Q.; Yin, L.; Liu, W.; Li, H.; Yu, B.; Wu, J. Inhibition of microRNA let-7i depresses maturation and functional state of dendritic cells in response to lipopolysaccharide stimulation via targeting suppressor of cytokine signaling 1. J. Immunol. 2011, 187, 1674–1683. [Google Scholar] [CrossRef] [PubMed]
- Krichevsky, A.M.; Gabriely, G. miR-21: A small multi-faceted RNA. J. Cell. Mol. Med. 2009, 13, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Sheedy, F.J. Turning 21: Induction of miR-21 as a Key Switch in the Inflammatory Response. Front. Immunol. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Smyth, L.A.; Boardman, D.A.; Tung, S.L.; Lechler, R.; Lombardi, G. MicroRNAs affect dendritic cell function and phenotype. Immunology 2015, 144, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Hashimi, S.T.; Fulcher, J.A.; Chang, M.H.; Gov, L.; Wang, S.; Lee, B. MicroRNA profiling identifies miR-34a and miR-21 and their target genes JAG1 and WNT1 in the coordinate regulation of dendritic cell differentiation. Blood 2009, 114, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Huang, X.; Zhang, X.; Roensch, K.; Cao, Q.; Nakayama, K.I.; Blazar, B.R.; Zeng, Y.; Zhou, X. miR-221 and miR-155 regulate human dendritic cell development, apoptosis, and IL-12 production through targeting of p27kip1, KPC1, and SOCS-1. Blood 2011, 117, 4293–4303. [Google Scholar] [CrossRef] [PubMed]
- Woltman, A.M.; van der Kooij, S.W.; Coffer, P.J.; Offringa, R.; Daha, M.R.; van Kooten, C. Rapamycin specifically interferes with GM-CSF signaling in human dendritic cells, leading to apoptosis via increased p27KIP1 expression. Blood 2003, 101, 1439–1445. [Google Scholar] [CrossRef] [PubMed]
- Mayoral, R.J.; Pipkin, M.E.; Pachkov, M.; van Nimwegen, E.; Rao, A.; Monticelli, S. MicroRNA-221–222 regulate the cell cycle in mast cells. J. Immunol. 2009, 182, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, H.; Schnorfeil, F.M.; Brocker, T. Differentially expressed microRNAs regulate plasmacytoid vs. conventional dendritic cell development. Mol. Immunol. 2010, 48, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Pflanz, S.; Hibbert, L.; Mattson, J.; Rosales, R.; Vaisberg, E.; Bazan, J.F.; Phillips, J.H.; McClanahan, T.K.; de Waal Malefyt, R.; Kastelein, R.A. WSX-1 and glycoprotein 130 constitute a signal-transducing receptor for IL-27. J. Immunol. 2004, 172, 2225–2231. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Cao, Z.; Chen, J.; Li, R.; Cao, Y.; Zhu, C.; Wu, K.; Wu, J.; Liu, F.; Zhu, Y. Influenza A virus induces interleukin-27 through cyclooxygenase-2 and protein kinase A signaling. J. Biol. Chem. 2012, 287, 11899–11910. [Google Scholar] [CrossRef] [PubMed]
- Frank, A.C.; Zhang, X.; Katsounas, A.; Bharucha, J.P.; Kottilil, S.; Imamichi, T. Interleukin-27, an anti-HIV-1 cytokine, inhibits replication of hepatitis C virus. J. Interferon Cytokine Res. 2010, 30, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Ten Oever, B.R. RNA viruses and the host microRNA machinery. Nat. Rev. Microbiol. 2013, 11, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Tam, S.; Tsao, M.S.; McPherson, J.D. Optimization of miRNA-seq data preprocessing. Brief. Bioinform. 2015, 16, 950–963. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: MicroRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Kolde, R. Pheatmap: Pretty Heatmaps, R Package Version 1.0.8, 2015. Available online: https://cran.r-project.org/web/packages/pheatmap/index.html (accessed on 27 April 2017).
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef] [PubMed]
- Reiche, K.; Stadler, P.F. RNAstrand: Reading direction of structured RNAs in multiple sequence alignments. Algorithms Mol. Biol. AMB 2007, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG Pathway | p-Values | Gene Count | miRNA Count |
|---|---|---|---|
| ErbB signaling pathway | 8.98 × 10−6 | 58 | 35 |
| Wnt signaling pathway | 4.60 × 10−5 | 84 | 36 |
| TGF-β signaling pathway | 4.99 × 10−5 | 50 | 33 |
| MAPK signaling pathway | 1.30 × 10−3 | 143 | 38 |
| PI3K-Akt signaling pathway | 1.60 × 10−2 | 177 | 41 |
| Novel miRNA | GenBank Accession Number | Mature Sequence | Precursor Genomic Location (hg38) | Precursor Minimum Free Energy (kcal/mol) | Encoding Gene | Gene Function |
|---|---|---|---|---|---|---|
| QXBT1 | KY994043 | ugucuguucccugucucucuag | chr10:48966435-48966526 | −38.7 | WDFY4 | exonic |
| QXBT2 | KY994044 | ugugucccuaugaaucucaugu | chr2:94772066-94772132 | −22.9 | ANKRD20A8P | ncRNA_intronic |
| QXBT3 | KY994045 | uaccucucccaaaacucaugugga | chr9:40297854-40297928 | −33.4 | ANKRD20A3; FAM95B1 | intergenic |
| QXBT4 | KY994046 | uucccucacuguaaacagagu | chr16:21647236-21647297 | −24.1 | IGSF6; METTL9 | exonic |
| QXBT5 | KY994047 | ucugucccaggcccagacuu | chr4:77048486-77048550 | −24.7 | CCNI | exonic |
| QXBT6 | KY994048 | aacaggccuugcucugcucacaga | chr3:52523356-52523428 | −44.7 | STAB1 | exonic |
| QXBT7 | KY994049 | ucucucucucuccgugucagugu | chr10:48827482-48827544 | −34.3 | WDFY4 | intronic |
| QXBT8 | KY994050 | agggagcggagaagcgagcgcag | chr15:45167265-45167327 | −40.2 | SHF | 3′UTR |
| QXBT9 | KY994051 | gaagcagcgccugucgcaacucg | chr17:78140754-78140815 | −30.6 | TMC8 | intronic |
| QXBT10 | KY994052 | uaauguaguugccacuaggaga | chr1:19910424-19910514 | −24.5 | OTUD3 | 3′UTR |
| QXBT11 | KY994053 | aaaagcuguccacuguagaguu | chr9:32456301-32456370 | −34.6 | DDX58 | 3′UTR |
| QXBT12 | KY994054 | cucccacugcuucacuugacuag | chr4:165400665-165400731 | −23.6 | CPE | intronic |
| QXBT13 | KY994055 | ccugucugagcgucgcu | chr14:16057563-16057625 | −21.7 | NA | intergenic |
| QXBT14 | KY994056 | acugugguaauucuagagcu | chr22:11629695-11629789 | −8.4 | NA; LOC102723769 | intergenic |
| QXBT15 | KY994057 | aggacuggaugucgggcugcau | chr4:141707942-141708053 | −22.6 | IL15 | intronic |
| QXBT16 | KY994058 | uuuugugugucagggugcaggu | chr14:94113640-94113698 | −21.9 | IFI27 | intronic |
| QXBT17 | KY994059 | gcgggagaggcggguggac | chr2:136117376-136117461 | −45.5 | CXCR4 | intronic |
| QXBT18 | KY994060 | acuccucauuuguaaacucagg | chrX:129790441-129790501 | −36.8 | SASH3 | intronic |
| QXBT19 | KY994061 | uccgcugcagccccucgacgu | chr8:55880664-55880722 | −33.1 | LYN | intronic |
| QXBT20 | KY994062 | acguggacuccagacucucugu | chr17:42311190-42311260 | −30.5 | STAT5A | 3′UTR |
| QXBT21 | KY994063 | acccucaguccguauuggucucu | chr17:7306830-7306888 | −32.8 | EIF5A | upstream |
| QXBT22 | KY994064 | aaagcccgugggggaccuguuc | chr22:17108635-17108691 | −22.2 | IL17RA | exonic |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.; Chen, Q.; Sowrirajan, B.; Bosche, M.; Imamichi, T.; Sherman, B.T. Genome-Wide Analyses of MicroRNA Profiling in Interleukin-27 Treated Monocyte-Derived Human Dendritic Cells Using Deep Sequencing: A Pilot Study. Int. J. Mol. Sci. 2017, 18, 925. https://doi.org/10.3390/ijms18050925
Hu X, Chen Q, Sowrirajan B, Bosche M, Imamichi T, Sherman BT. Genome-Wide Analyses of MicroRNA Profiling in Interleukin-27 Treated Monocyte-Derived Human Dendritic Cells Using Deep Sequencing: A Pilot Study. International Journal of Molecular Sciences. 2017; 18(5):925. https://doi.org/10.3390/ijms18050925
Chicago/Turabian StyleHu, Xiaojun, Qian Chen, Bharatwaj Sowrirajan, Marjorie Bosche, Tomozumi Imamichi, and Brad T. Sherman. 2017. "Genome-Wide Analyses of MicroRNA Profiling in Interleukin-27 Treated Monocyte-Derived Human Dendritic Cells Using Deep Sequencing: A Pilot Study" International Journal of Molecular Sciences 18, no. 5: 925. https://doi.org/10.3390/ijms18050925
APA StyleHu, X., Chen, Q., Sowrirajan, B., Bosche, M., Imamichi, T., & Sherman, B. T. (2017). Genome-Wide Analyses of MicroRNA Profiling in Interleukin-27 Treated Monocyte-Derived Human Dendritic Cells Using Deep Sequencing: A Pilot Study. International Journal of Molecular Sciences, 18(5), 925. https://doi.org/10.3390/ijms18050925