Quantitative Evaluation of Drug Resistance Profile of Cells Expressing Wild-Type or Genetic Polymorphic Variants of the Human ABC Transporter ABCC4



Abstract
:
1. Introduction
2. Results
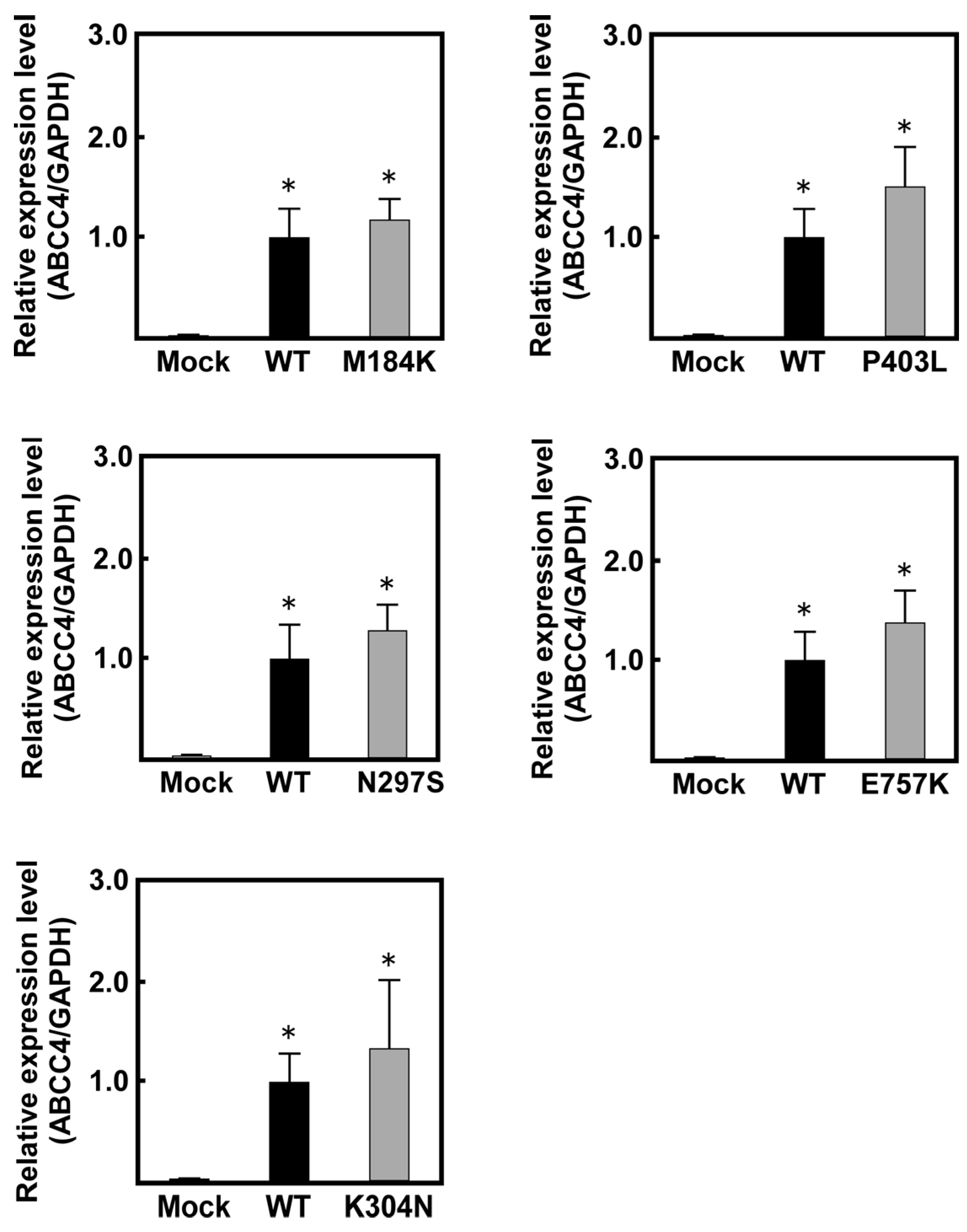
2.1. Levels of ABCC4 mRNA and Protein in Cells Established Using the Flp-In™ System
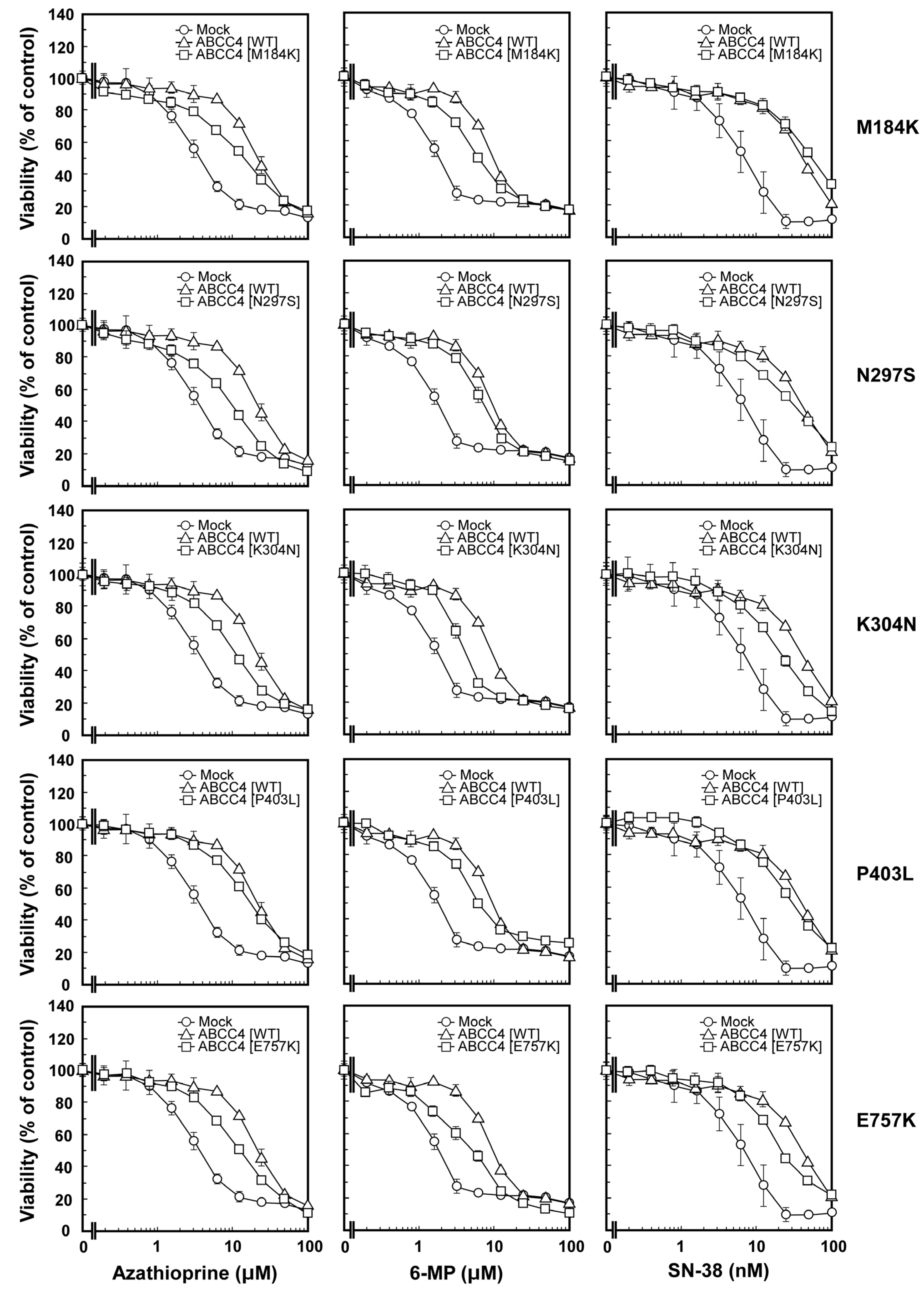
2.2. Anticancer Drug Resistance Properties of Cells Established Using the Flp-In™ System
3. Discussion
3.1. Establishment of Human ABCC4-Expressing Cells Using the Flp-In™ System
3.2. Anticancer Drug Resistance Properties of Cells Established Using the Flp-In™ System
4. Materials and Methods
4.1. Chemicals and Biological Reagents
4.2. Subcloning of Human ABCC4 cDNA
4.3. Preparation of Plasmids Carrying the ABCC4 SNP Variant cDNA
4.4. Cell Culture
4.5. Preparation of Total RNA and Synthesis of First-Strand cDNA
4.6. Quantitative Real-Time PCR
4.7. MTT Assay
4.8. Preparation of Cell Lysates for Sodium Dodecyl Sulfate Poly-Acrylamide Gel Electrophoresis (SDS-PAGE)
4.9. SDS-PAGE and Western Blotting
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| ABCC4 | ATP-binding cassette subfamily C member 4 |
| BSA | bovine serum albumin |
| DMEM | high-glucose Dulbecco’s modified Eagle’s medium |
| DTT | dithiothreitol |
| EC50 | half maximal (50%) effective concentration |
| FBS | fetal bovine serum |
| FRT | Flp recombination target |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| HRP | horseradish peroxidase |
| MTT | 3-(4,5-dimethyl-2-thiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide |
| MRP | multi drug resistance protein |
| NCBI | National Center for Biotechnology Information |
| PBS(−) | phosphate-buffered saline without both Ca2+ and Mg2+ |
| PCR | polymerase chain reaction |
| qPCR | quantitative real-time PCR |
| SDS-PAGE | sodium dodecyl sulfate poly-acrylamide gel electrophoresis |
| SN-38 | 7-Ethyl-10-hydroxy-camptothecin |
| SNP | single-nucleotide polymorphism |
| TBS | Tris-buffered saline |
| TBST | TBS with 0.05% (v/v) Tween 20 |
| WT | wild type |
References
- Imai, Y.; Nakane, M.; Kage, K.; Tsukahara, S.; Ishikawa, E.; Tsuruo, T.; Miki, Y.; Sugimoto, Y. C421A polymorphism in the human breast cancer resistance protein gene is associated with low expression of Q141K protein and low-level drug resistance. Mol. Cancer Ther. 2002, 1, 611–616. [Google Scholar] [PubMed]
- Sakurai, A.; Tamura, A.; Onishi, Y.; Ishikawa, T. Genetic polymorphisms of ATP-binding cassette transporters ABCB1 and ABCG2: Therapeutic implications. Exp. Opin. Pharmacother. 2005, 6, 2455–2473. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.; Rzhetsky, A.; Allikmets, R. The human ATP-binding cassette (ABC) transporter superfamily. Genome Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Borst, P.; de Wolf, C.; van de Wetering, K. Multidrug resistance-associated proteins 3, 4, and 5. Pflügers Arch. Eur. J. Phy. 2007, 453, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Cascorbi, I. Role of pharmacogenetics of ATP-binding cassette transporters in the pharmacokinetics of drugs. Pharmacol. Ther. 2006, 112, 457–473. [Google Scholar] [CrossRef] [PubMed]
- Fromm, M.F. Importance of P-glycoprotein at blood-tissue barriers. Trends Pharmacol. Sci. 2004, 25, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Ho, R.H.; Kim, R.B. Transporters and drug therapy: Implications for drug disposition and disease. Clin. Pharmacol. Ther. 2005, 78, 260–277. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Suzuki, H.; Horie, T.; Sugiyama, Y. Apical/basolateral surface expression of drug transporters and its role in vectorial drug transport. Pharm. Res. 2005, 22, 1559–1577. [Google Scholar] [CrossRef] [PubMed]
- Keppler, D. Uptake and efflux transporters for conjugates in human hepatocytes. Methods Enzymol. 2005, 400, 531–542. [Google Scholar] [PubMed]
- Klein, I.; Sarkadi, B.; Varadi, A. An inventory of the human ABC proteins. Biochim. Biophys. Acta 1999, 1461, 237–262. [Google Scholar] [CrossRef]
- Chan, L.M.; Lowes, S.; Hirst, B.H. The ABCs of drug transport in intestine and liver: Efflux proteins limiting drug absorption and bioavailability. Eur. J. Pharm. Sci. 2004, 21, 25–51. [Google Scholar] [CrossRef] [PubMed]
- Imaoka, T.; Kusuhara, H.; Adachi, M.; Schuetz, J.D.; Takeuchi, K.; Sugiyama, Y. Functional involvement of multidrug resistance-associated protein 4 (MRP4/ABCC4) in the renal elimination of the antiviral drugs adefovir and tenofovir. Mol. Pharmacol. 2007, 71, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, P.; Schwab, M.; Takenaka, K.; Nachagari, D.; Morgan, J.; Leslie, M.; Du, W.; Boyd, K.; Cheok, M.; Nakauchi, H.; et al. Transporter-mediated protection against thiopurine-induced hematopoietic toxicity. Cancer Res. 2008, 68, 4983–4989. [Google Scholar] [CrossRef] [PubMed]
- Schuetz, J.D.; Connelly, M.C.; Sun, D.; Paibir, S.G.; Flynn, P.M.; Srinivas, R.V.; Kumar, A.; Fridland, A. MRP4: A previously unidentified factor in resistance to nucleoside-based antiviral drugs. Nat. Med. 1999, 5, 1048–1051. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Klein-Szanto, A.J.; Kruh, G.D. Analysis of the MRP4 drug resistance profile in transfected NIH3T3 cells. J. Natl. Cancer Inst. 2000, 92, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Adachi, M.; Sampath, J.; Lan, L.B.; Sun, D.; Hargrove, P.; Flatley, R.; Tatum, A.; Edwards, M.Z.; Wezeman, M.; Matherly, L.; et al. Expression of MRP4 confers resistance to ganciclovir and compromises bystander cell killing. J. Biol. Chem. 2002, 277, 38998–39004. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.S.; Lee, K.; Walther, S.; Raftogianis, R.B.; Kuwano, M.; Zeng, H.; Kruh, G.D. Analysis of methotrexate and folate transport by multidrug resistance protein 4 (ABCC4): MRP4 is a component of the methotrexate efflux system. Cancer Res. 2002, 62, 3144–3150. [Google Scholar] [PubMed]
- Wielinga, P.R.; Reid, G.; Challa, E.E.; van der Heijden, I.; van Deemter, L.; de Haas, M.; Mol, C.; Kuil, A.J.; Groeneveld, E.; Schuetz, J.D.; et al. Thiopurine metabolism and identification of the thiopurine metabolites transported by MRP4 and MRP5 overexpressed in human embryonic kidney cells. Mol. Pharmacol. 2002, 62, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Leggas, M.; Adachi, M.; Scheffer, G.L.; Sun, D.; Wielinga, P.; Du, G.; Mercer, K.E.; Zhuang, Y.; Panetta, J.C.; Johnston, B.; et al. Mrp4 confers resistance to topotecan and protects the brain from chemotherapy. Mol. Cell. Biol. 2004, 24, 7612–7621. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Zhang, J.; Tan, T.M.; Chan, E.; Duan, W.; Chan, S.Y.; Boelsterli, U.A.; Ho, P.C.; Yang, H.; Bian, J.S.; et al. Human multidrug resistance associated protein 4 confers resistance to camptothecins. Pharm. Res. 2005, 22, 1837–1853. [Google Scholar] [CrossRef] [PubMed]
- Norris, M.D.; Smith, J.; Tanabe, K.; Tobin, P.; Flemming, C.; Scheffer, G.L.; Wielinga, P.; Cohn, S.L.; London, W.B.; Marshall, G.M.; et al. Expression of multidrug transporter MRP4/ABCC4 is a marker of poor prognosis in neuroblastoma and confers resistance to irinotecan in vitro. Mol. Cancer Ther. 2005, 4, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Zhang, J.; Chan, S.Y.; Tan, T.M.; Duan, W.; Huang, M.; Zhu, Y.Z.; Chan, E.; Yu, Q.; Nie, Y.Q.; et al. Topotecan is a substrate for multidrug resistance associated protein 4. Curr. Drug Metab. 2006, 7, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.L.; Lamba, J.; Aquilante, C.L.; Schuetz, E.; Fletcher, C.V. Pharmacogenetic characteristics of indinavir, zidovudine, and lamivudine therapy in HIV-infected adults: A pilot study. J. Acquir. Immune Defic. Syndr. 2006, 42, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Janke, D.; Mehralivand, S.; Strand, D.; Godtel-Armbrust, U.; Habermeier, A.; Gradhand, U.; Fischer, C.; Toliat, M.R.; Fritz, P.; Zanger, U.M.; et al. 6-mercaptopurine and 9-(2-phosphonyl-methoxyethyl) adenine (PMEA) transport altered by two missense mutations in the drug transporter gene ABCC4. Hum. Mutat. 2008, 29, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Ban, H.; Andoh, A.; Imaeda, H.; Kobori, A.; Bamba, S.; Tsujikawa, T.; Sasaki, M.; Saito, Y.; Fujiyama, Y. The multidrug-resistance protein 4 polymorphism is a new factor accounting for thiopurine sensitivity in Japanese patients with inflammatory bowel disease. J. Gastroenterol. 2010, 45, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Marensi, V.; Conseil, G.; Le, X.C.; Cole, S.P.; Leslie, E.M. Polymorphic variants of MRP4/ABCC4 differentially modulate the transport of methylated arsenic metabolites and physiological organic anions. Biochem. Pharmacol. 2016, 120, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Tamura, A.; Wakabayashi, K.; Onishi, Y.; Nakagawa, H.; Tsuji, M.; Matsuda, Y.; Ishikawa, T. Genetic polymorphisms of human ABC transporter ABCG2: Development of the standard method for functional validation of SNPs by using the Flp recombinase system. J. Exp. Ther. Oncol. 2006, 6, 1–11. [Google Scholar] [PubMed]
- Tamura, A.; Watanabe, M.; Saito, H.; Nakagawa, H.; Kamachi, T.; Okura, I.; Ishikawa, T. Functional validation of the genetic polymorphisms of human ATP-binding cassette (ABC) transporter ABCG2: Identification of alleles that are defective in porphyrin transport. Mol. Pharmacol. 2006, 70, 287–296. [Google Scholar] [PubMed]
- Furukawa, T.; Wakabayashi, K.; Tamura, A.; Nakagawa, H.; Morishima, Y.; Osawa, Y.; Ishikawa, T. Major SNP (Q141K) variant of human ABC transporter ABCG2 undergoes lysosomal and proteasomal degradations. Pharm. Res. 2009, 26, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, K.; Nakagawa, H.; Ishikawa, T.; Kubo, M.; Mushiroda, T. ABCB1 polymorphism is associated with atorvastatin-induced liver injury in Japanese population. BMC Genet. 2016, 17, 79. [Google Scholar] [CrossRef] [PubMed]
- Campa, D.; Muller, P.; Edler, L.; Knoefel, L.; Barale, R.; Heussel, C.P.; Thomas, M.; Canzian, F.; Risch, A. A comprehensive study of polymorphisms in ABCB1, ABCC2 and ABCG2 and lung cancer chemotherapy response and prognosis. Int. J. Cancer 2012, 131, 2920–2928. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, K.; Nakagawa, H.; Adachi, T.; Kii, I.; Kobatake, E.; Kudo, A.; Ishikawa, T. Identification of cysteine residues critically involved in homodimer formation and protein expression of human ATP-binding cassette transporter ABCG2: A new approach using the flp recombinase system. J. Exp. Ther. Oncol. 2006, 5, 205–222. [Google Scholar] [PubMed]
- Kosztyu, P.; Dolezel, P.; Mlejnek, P. Can P-glycoprotein mediate resistance to nilotinib in human leukaemia cells? Pharmacol. Res. 2013, 67, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Kosztyu, P.; Bukvova, R.; Dolezel, P.; Mlejnek, P. Resistance to daunorubicin, imatinib, or nilotinib depends on expression levels of ABCB1 and ABCG2 in human leukemia cells. Chem. Biol. Interact. 2014, 5, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Tamura, A.; Wakabayashi, K.; Hoshijima, K.; Komada, M.; Yoshida, T.; Kometani, S.; Matsubara, T.; Mikuriya, K.; Ishikawa, T. Ubiquitin-mediated proteasomal degradation of non-synonymous SNP variants of human ABC transporter ABCG2. Biochem. J. 2008, 411, 623–631. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant | rsNumber | Nucleotide Position | Nucleotide Change | Amino Acid Position | Amino Acid Change |
|---|---|---|---|---|---|
| M184K | rs45454092 | 551 | t > a | 184 | Met > Lys |
| N297S | rs200387797 | 890 | a > g | 297 | Asn > Ser |
| K304N | rs2274407 | 912 | g > t | 304 | Lys > Asn |
| P403L | rs11568705 | 1208 | c > t | 403 | Pro > Leu |
| E757K | rs3765534 | 2269 | g > a | 757 | Glu > Lys |
| Compounds | EC50 | ||||||
|---|---|---|---|---|---|---|---|
| Mock | WT | M184K | N297S | K304N | P403L | E757K | |
| ATRA (μM) | 40.0 ± 10.1 | 42.1 ± 4.8 | 43.4 ± 3.2 | 48.1 ± 8.0 | 40.3 ± 4.8 | 41.0 ± 2.9 | 42.0 ± 4.3 |
| Azathioprine (μM) | 4.2 ± 1.0 | 21.5 ± 1.0 * | 14.3 ± 1.8 *,** | 12.6 ± 2.4 *,** | 12.6 ± 2.2 *,** | 17.9 ± 1.0 * | 15.2 ± 3.3 *,** |
| Etoposide (nM) | 204.9 ± 16.2 | 256.6 ± 31.7 | 210.3 ± 13.2 | 254.1 ± 28.2 | 326.3 ± 33.6 * | 321.0 ± 14.6 * | 208.4 ± 17.9 |
| 5-FU (μM) | 7.7 ± 1.4 | 6.3 ± 2.2 | 11.1 ± 1.4 | 4.5 ± 1.2 | 11.0 ± 2.7 | 4.3 ± 1.0 | 5.9 ± 1.2 |
| 6-Mercaptopurine (μM) | 1.9 ± 0.4 | 9.1 ± 1.8 * | 6.4 ± 0.7 * | 6.6 ± 0.9 * | 4.6 ± 0.3 ** | 6.4 ± 0.5 * | 5.1 ± 1.4 ** |
| SN-38 (nM) | 8.7 ± 1.1 | 45.5 ± 10.2 * | 50.2 ± 6.6 * | 29.5 ± 8.5 * | 22.5 ± 0.3 ** | 24.4 ± 7.4 ** | 21.6 ± 1.6 ** |
| Vincristine (nM) | 2.9 ± 1.1 | 1.6 ± 0.5 | 2.1 ± 0.7 | 2.1 ± 0.8 | 2.3 ± 0.4 | 1.9 ± 0.4 | 1.7 ± 0.3 |
| Variant | Forward/Reverse (F/R) Primers | Primer Sequence (5′→3′) | Primer Length (Bases) | % GC | Tm (°C) |
|---|---|---|---|---|---|
| M184K | F | CACTTCGTCTTAGTAACAAGGCCATGGGGAAGACAAC | 37 | 48.6 | 80.5 |
| R | GTTGTCTTCCCCATGGCCTTGTTACTAAGACGAAGTG | ||||
| N297S | F | GCCTGGGAAAAGTCATTTTCAAGTCTTATTACCAATTTGAGAAAG | 45 | 35.6 | 78.9 |
| R | CTTTCTCAAATTGGTAATAAGACTTGAAAATGACTTTTCCCAGGC | ||||
| K304N | F | TCTTATTACCAATTTGAGAAATAAGGAGATTTCCAAGATTCTGAG | 45 | 31.1 | 77.9 |
| R | CTCAGAATCTTGGAAATCTCCTTATTTCTCAAATTGGTAATAAGA | ||||
| P403L | F | CGCAACCGTCAGCTGCTGTCAGATGGTAAAAAGATG | 36 | 50.0 | 80.5 |
| R | CATCTTTTTACCATCTGACAGCAGCTGACGGTTGCG | ||||
| E757K | F | AAATGGAGGAGGAAATGTAACCAAGAAGCTAGATCTTAACTGGTA | 45 | 37.8 | 79.7 |
| R | TACCAGTTAAGATCTAGCTTCTTGGTTACATTTCCTCCTCCATTT |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsukamoto, M.; Sato, S.; Satake, K.; Miyake, M.; Nakagawa, H. Quantitative Evaluation of Drug Resistance Profile of Cells Expressing Wild-Type or Genetic Polymorphic Variants of the Human ABC Transporter ABCC4. Int. J. Mol. Sci. 2017, 18, 1435. https://doi.org/10.3390/ijms18071435
Tsukamoto M, Sato S, Satake K, Miyake M, Nakagawa H. Quantitative Evaluation of Drug Resistance Profile of Cells Expressing Wild-Type or Genetic Polymorphic Variants of the Human ABC Transporter ABCC4. International Journal of Molecular Sciences. 2017; 18(7):1435. https://doi.org/10.3390/ijms18071435
Chicago/Turabian StyleTsukamoto, Megumi, Shiori Sato, Kazuhiro Satake, Mizuki Miyake, and Hiroshi Nakagawa. 2017. "Quantitative Evaluation of Drug Resistance Profile of Cells Expressing Wild-Type or Genetic Polymorphic Variants of the Human ABC Transporter ABCC4" International Journal of Molecular Sciences 18, no. 7: 1435. https://doi.org/10.3390/ijms18071435
APA StyleTsukamoto, M., Sato, S., Satake, K., Miyake, M., & Nakagawa, H. (2017). Quantitative Evaluation of Drug Resistance Profile of Cells Expressing Wild-Type or Genetic Polymorphic Variants of the Human ABC Transporter ABCC4. International Journal of Molecular Sciences, 18(7), 1435. https://doi.org/10.3390/ijms18071435