Age-Related Loss of Cohesion: Causes and Effects

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Mammalian Cohesion
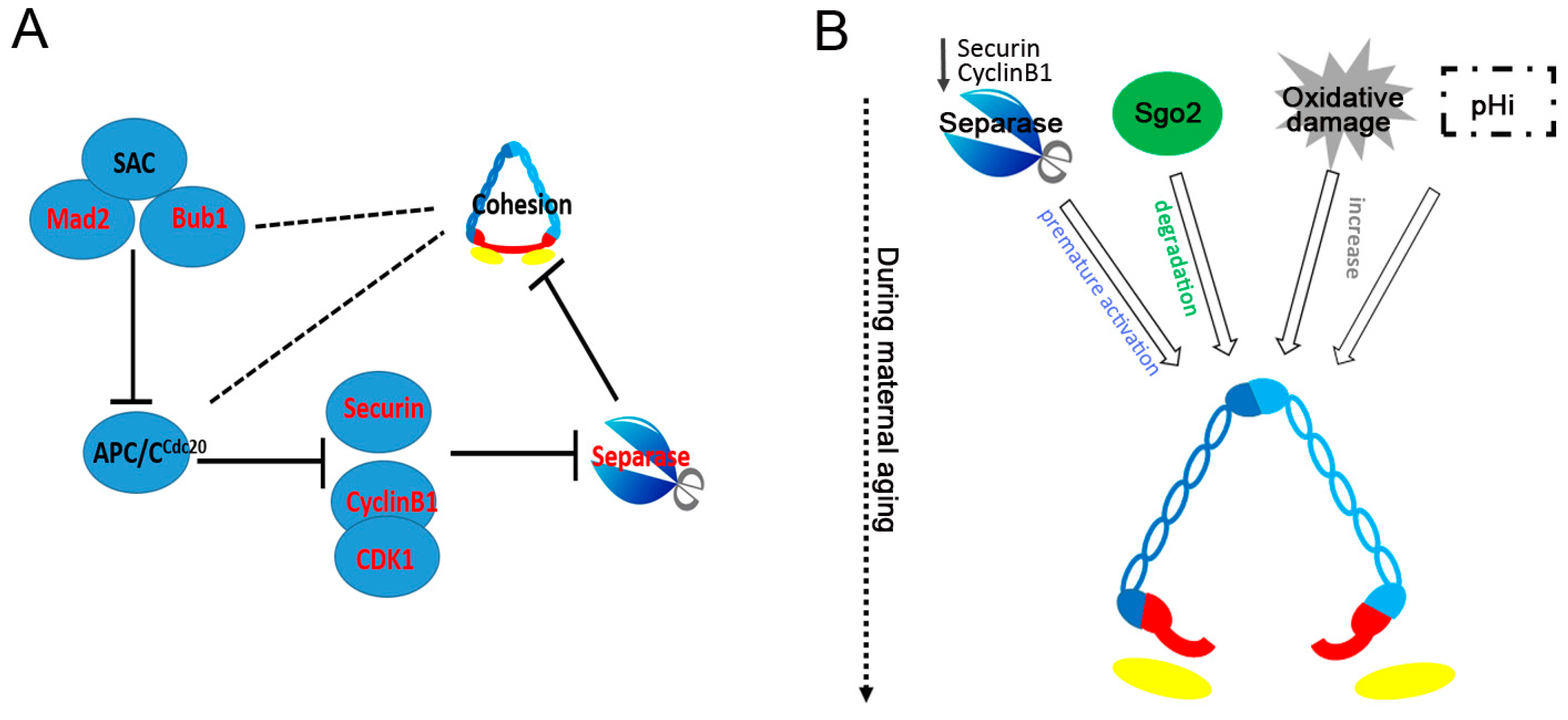
3. The Cause of Age-Related Loss of Cohesion
3.1. Increased Sensibility of Separase
3.2. The SAC and APC/CCdc20
3.3. Premature Shugoshin 2 Degradation
3.4. Oxidative Damage
3.5. Other Factors
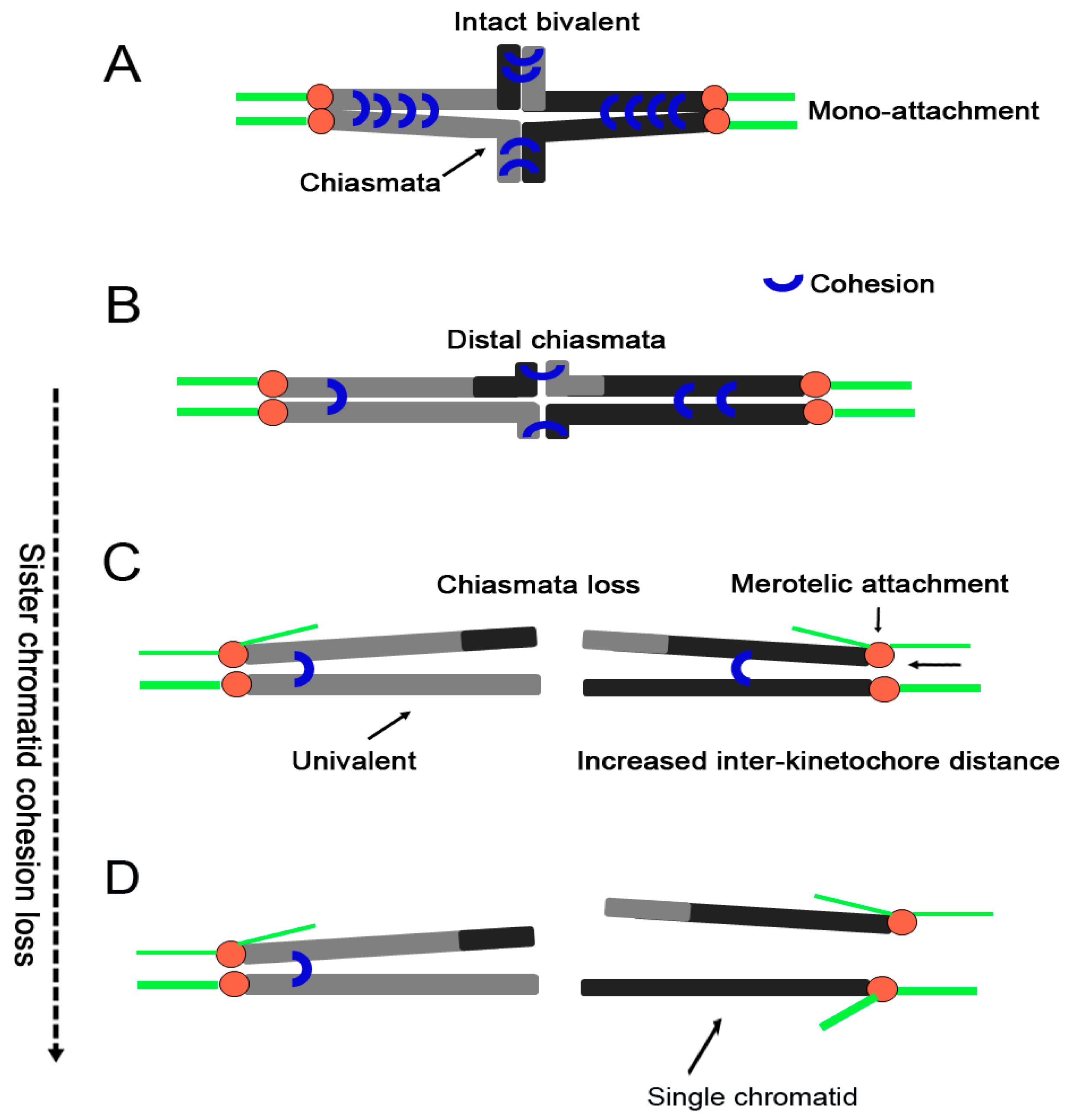
4. The Effects of Age-Related Loss of Cohesion
4.1. Destabilization of Chiasmata
4.2. Bivalents Separation into Univalents
4.3. Kinetochore Orientation Alteration
5. Conclusions and Future Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Anifandis, G.; Markandona, O.; Dafopoulos, K.; Messini, C.; Tsezou, A.; Dimitraki, M.; Georgoulias, P.; Daponte, A.; Messinis, I. Embryological Results of Couples Undergoing ICSI-ET Treatments with Males Carrying the Single Nucleotide Polymorphism rs175080 of the MLH3 Gene. Int. J. Mol. Sci. 2017, 18, 314. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.M.; Li, J.; Tang, J.X.; Cheng, S.R.; Deng, S.L.; Jin, C.; Zhang, Y.; Wang, X.X.; Zhou, C.X.; Liu, Y.X. Elevated intracellular pH appears in aged oocytes and causes oocyte aneuploidy associated with the loss of cohesion in mice. Cell Cycle 2016, 18, 2454–2463. [Google Scholar] [CrossRef] [PubMed]
- Merriman, J.A.; Jennings, P.C.; McLaughlin, E.A.; Jones, K.T. Effect of aging on superovulation efficiency, aneuploidy rates, and sister chromatid cohesion in mice aged up to 15 months. Biol. Reprod. 2011, 49, 1–6. [Google Scholar]
- Camlin, N.J.; McLaughlin, E.A.; Holt, J.E. The use of C57Bl/6 × CBA F1 hybrid cross as a model for human age-related oocyte aneuploidy. Mol. Reprod. Dev. 2017, 84, 6–7. [Google Scholar]
- Fragouli, E.; Alfarawati, S.; Goodall, N.N.; Sanchez-Garcia, J.F.; Colls, P.; Wells, D. The cytogenetics of polar bodies: Insights into female meiosis and the diagnosis of aneuploidy. Mol. Hum. Reprod. 2011, 17, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Kuliev, A.; Zlatopolsky, Z.; Kirillova, I.; Spivakova, J.; Cieslak, J. Meiosis errors in over 20,000 oocytes studied in the practice of preimplantation aneuploidy testing. Reprod. Biomed. Online 2011, 22, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Lister, L.M.; Kouznetsova, A.; Hyslop, L.A.; Kalleas, D.; Pace, S.L.; Barel, J.C.; Nathan, A.; Floros, V.; Adelfalk, C.; Watanabe, Y.; et al. Age-related meiotic segregation errors in mammalian oocytes are preceded by depletion of cohesin and Sgo2. Curr. Biol. 2010, 20, 1511–1521. [Google Scholar] [CrossRef] [PubMed]
- Jessberger, R. Age-related aneuploidy through cohesion exhaustion. EMBO Rep. 2012, 13, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Hodges, C.A.; Revenkova, E.; Jessberger, R.; Hassold, T.J.; Hunt, P.A. SMC1β-deficient female mice provide evidence that cohesins are a missing link in age-related nondisjunction. Nat. Genet. 2005, 37, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Chiang, T.; Schultz, R.M.; Lampson, M.A. Meiotic origins of maternal age-related aneuploidy. Biol. Reprod. 2012, 86, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, Y.; Hashimoto, S.; Nakaoka, Y.; Kouznetsova, A.; Hoog, C.; Kitajima, T.S. Bivalent separation into univalents precedes age-related meiosis I errors in oocytes. Nat. Commun. 2015, 6, 7550. [Google Scholar] [CrossRef] [PubMed]
- Shomper, M.; Lappa, C.; FitzHarris, G. Kinetochore microtubule establishment is defective in oocytes from aged mice. Cell Cycle 2014, 13, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Zielinska1, A.P.; Holubcova, Z.; Blayney, M.; Elder, K.; Schuh, M. Sister kinetochore splitting and precocious disintegration of bivalents could explain the maternal age effect. Elife 2015, 4, e11389. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Tan, S.L.; Hartshorne, G.M.; McAinsh, A.D. Unique geometry of sister kinetochores in human oocytes during meiosis I may explain maternal age-associated increases in chromosomal abnormalities. Biol. Open 2015, 5, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.T.; Lane, S.I. Molecular causes of aneuploidy in mammalian eggs. Development 2013, 140, 3719–3730. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.J.; Severson, A.F.; Meyer, B.J. Condensin and cohesin complexity: The expanding repertoire of functions. Nat. Rev. Genet. 2010, 11, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Uhlmann, F. Chromosome cohesion and segregation in mitosis and meiosis. Curr. Opin. Cell. Biol. 2001, 13, 754–761. [Google Scholar] [CrossRef]
- Nasmyth, K.; Haering, C.H. Cohesin: Its roles and mechanisms. Annu. Rev. Genet. 2009, 43, 525–558. [Google Scholar] [CrossRef] [PubMed]
- Revenkova, E.; Eijpe, M.; Heyting, C.; Gross, B.; Jessberger, R. Novel meiosis-specific isoform of mammalian SMC1. Mol. Cell. Biol. 2001, 21, 6984–6998. [Google Scholar] [CrossRef] [PubMed]
- Revenkova, E.; Eijpe, M.; Heyting, C.; Hodges, C.A.; Hunt, P.A.; Liebe, B.; Scherthan, H.; Jessberger, R. Cohesin SMC1β is required for meiotic chromosome dynamics, sister chromatid cohesion and DNA recombination. Nat. Cell. Biol. 2004, 6, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T. At the heart of the chromosome: SMC proteins in action. Nat. Rev. Mol. Cell. Biol. 2006, 7, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cruz, R.; Brieno, M.A.; Roig, I.; Grossmann, M.; Velilla, E.; Pujol, A.; Cabero, L.; Pessarrodona, A.; Barbero, J.L.; Garcia Caldés, M. Dynamics of cohesin proteins REC8, STAG3, SMC1 β and SMC3 are consistent with a role in sister chromatid cohesion during meiosis in human oocytes. Hum. Reprod. 2010, 25, 2316–2332. [Google Scholar] [CrossRef] [PubMed]
- Schleiffer, A.; Kaitna, S.; Maurer-Stroh, S.; Glotzer, M.; Nasmyth, K.; Eisenhaber, F. Kleisins: A superfamily of bacterial and eukaryotic SMC protein partners. Mol. Cell. 2003, 11, 571–575. [Google Scholar] [CrossRef]
- Watanabe, Y.; Nurse, P. Cohesin Rec8 is required for reductional chromosome segregation at meiosis. Nature 1999, 400, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Parisi, S.; McKay, M.J.; Molnar, M.; Thompson, M.A.; van der Spek, P.J.; Drunen Schoenmaker, E.; van Kanaar, R.; Lehmann, E.; Hoeijmakers, J.H.; Kohli, J. Rec8p, a meiotic recombination and sister chromatid cohesion phosphoprotein of the Rad21p family conserved from fission yeast to humans. Mol. Cell. Biol. 1999, 19, 3515–3528. [Google Scholar] [CrossRef] [PubMed]
- Prieto, I.; Suja, J.A.; Pezzi, N.; Kremer, L.; Martínez-A, C.; Rufas, J.S.; Barbero, J.L. Mammalian STAG3 is a cohesin specific to sister chromatid arms in meiosis I. Nat. Cell. Biol. 2001, 3, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Pezzi, N.; Prieto, I.; Kremer, L.; Perez Jurado, L.A.; Valero, C.; Del Mazo, J.; Martinez, A.C.; Barbero, J.L. STAG3, a novel gene encoding a protein involved in meiotic chromosome pairing and location of STAG3-related genes flanking the Williams-Beuren syndrome deletion. FASEB J. 2000, 14, 581–592. [Google Scholar] [PubMed]
- Xu, H.; Beasley, M.D.; Warren, W.D.; van der Horst, G.T.; McKay, M.J. Absence of mouse REC8cohesin promotes synapsis of sister chromatids in meiosis. Dev. Cell. 2005, 8, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Klein, F.; Mahr, P.; Galova, M.; Buonomo, S.B.; Michaelis, C.; Nairz, K.; Nasmyth, K. A central role for cohesins in sister chromatid cohesion, formation of axial elements, and recombination during yeast meiosis. Cell 1999, 1, 91–103. [Google Scholar] [CrossRef]
- Eijpe, M.; Offenberg, H.; Jessberger, R.; Revenkova, E.; Heyting, C. Meiotic cohesin REC8 marks the axial elements of rat synaptonemal complexes before cohesins SMC1beta and SMC3. J. Cell. Biol. 2003, 160, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Kudo, N.R.; Anger, M.; Peters, A.H.F.M.; Stemmann, O.; Theussl, H.C.; Helmhart, W.; Kudo, H.; Heyting, C.; Nasmyth, K. Role of cleavage by separase of the Rec8 kleisin subunit of cohesin during mammalian meiosis I. J. Cell. Sci. 2009, 122, 2686–2698. [Google Scholar] [CrossRef] [PubMed]
- Chiang, T.; Duncan, F.E.; Schindler, K.; Schultz, R.M.; Lampson, M.A. Evidence that weakened centromere cohesion is a leading cause of age-related aneuploidy in oocytes. Curr. Biol. 2010, 20, 1522–1528. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Keefe, D.L. Defective cohesin is associated with age-dependent misaligned chromosomes in oocytes. Reprod. Biomed. Online 2008, 1, 103–112. [Google Scholar] [CrossRef]
- Tachibana-Konwalski, K.; Godwin, J.; van der Weyden, L.; Champion, L.; Kudo, N.R.; Adams, D.J.; Nasmyth, K. Rec8-containing cohesin maintains bivalents without turnover during the growing phase of mouse oocytes. Genes Dev. 2010, 24, 2505–2516. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, S.; Borsos, M.; Szydlowska, A.; Hirota, T.; Saitou, M.; Tachibana-Konwalski, K. Chromosome Cohesion Established by Rec8-Cohesin in Fetal Oocytes Is Maintained without Detectable Turnover in Oocytes Arrested for Months in Mice. Curr. Biol. 2016, 26, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Revenkova, E.; Herrmann, K.; Adelfalk, C.; Jessberger, R. Oocyte cohesion expression restricted to predictyate stages provides full fertility and prevents aneuploidy. Curr. Biol. 2010, 20, 1529–1533. [Google Scholar] [CrossRef] [PubMed]
- Weng, K.A.; Jeffreys, C.A.; Bickel, S.E. Rejuvenation of meiotic cohesion in oocytes during prophase I is required for chiasma maintenance and accurate chromosome segregation. PLoS Genet. 2014, 10, e1004607. [Google Scholar] [CrossRef] [PubMed]
- Hauf, S.; Waizenegger, I.C.; Peters, J.M. Cohesin Cleavage by Separase Required for Anaphase and Cytokinesis in Human Cells. Science 2001, 5533, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Waizenegger, I.C.; Hauf, S.; Meinke, A.; Peters, J.M. Two distinct pathways remove mammalian cohesin from chromosome arms in prophase and from centromeres in anaphase. Cell 2000, 103, 399–410. [Google Scholar] [CrossRef]
- Kudo, N.R.; Wassmann, K.; Anger, M.; Schuh, M.; Wirth, K.G.; Xu, H.; Helmhart, W.; Kudo, H.; McKay, M.; Maro, B.; et al. Resolution of chiasmata in oocytes requires separase-mediated proteolysis. Cell 2006, 126, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Herbert, M.; Levasseur, M.; Homer, H.; Yallop, K.; Murdoch, A.; McDougall, A. Homologue disjunction in mouse oocytes requires proteolysis of securin and cyclin B1. Nat. Cell. Biol. 2003, 5, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Stemmann, O.; Zou, H.; Gerber, S.A.; Gygi, S.P.; Kirschner, M.W. Dual inhibition of sister chromatid separation at metaphase. Cell 2001, 107, 715–726. [Google Scholar] [CrossRef]
- Gorr, I.H.; Boos, D.; Stemmann, O. Mutual inhibition of separase and Cdk1 by two-step complex formation. Mol. Cell. 2005, 19, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Nabti, I.; Reis, A.; Levasseur, M.; Stemmann, O.; Jones, K.T. Securin and not CDK1/cyclin B1 regulates sister chromatid disjunction during meiosis II in mouse eggs. Dev. Biol. 2008, 321, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Chiang, T.; Schultz, R.M.; Lampson, M.A. Age-dependent susceptibility of chromosome cohesion to premature separase activation in mouse oocytes. Biol. Reprod. 2011, 85, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.M. The anaphase promoting complex/cyclosome: A machine designed to destroy. Nat. Rev. Mol. Cell. Biol. 2006, 7, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.M. The anaphase-promoting complex: Proteolysis in mitosis and beyond. Mol. Cell 2002, 9, 931–943. [Google Scholar] [CrossRef]
- Musacchio, A.; Salmon, E.D. The spindle-assembly checkpoint in space and time. Nat. Rev. Mol. Cell. Biol. 2007, 8, 379–393. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, B.E.; Anger, M.; Kouznetsova, A.; Gil-Bernabe´, A.M.; Helmhart, W.; Kudo, N.R.; Wuensche, A.; Taylor, S.; Hoog, C.; Novak, B.; et al. Regulation of APC/C activity in oocytes by a Bub1-dependent spindle assembly checkpoint. Curr. Biol. 2009, 19, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Wassmann, K.; Benezra, R. Mitotic checkpoints: From yeast to cancer. Curr. Opin. Genet. Dev. 2001, 11, 83–90. [Google Scholar] [CrossRef]
- Fang, G.; Yu, H.; Kirschner, M.W. The checkpoint protein MAD2 and the mitotic regulator CDC20 form a ternary complex with the anaphase-promoting complex to control anaphase initiation. Genes Dev. 1998, 12, 1871–1883. [Google Scholar] [CrossRef] [PubMed]
- Homer, H.A.; McDougall, A.; Levasseur, M.; Yallop, K.; Murdoch, A.P.; Herbert, M. Mad2 prevents aneuploidy and premature proteolysis of cyclin B and securin during meiosis I in mouse oocytes. Genes Dev. 2005, 19, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Wassmann, K.; Niault, T.; Maro, B. Metaphase I arrest upon activation of the Mad2-dependent spindle checkpoint in mouse oocytes. Curr. Biol. 2003, 18, 1596–1608. [Google Scholar] [CrossRef]
- Perera, D.; Tilston, V.; Hopwood, J.A.; Barchi, M.; Boot-Handford, R.P.; Taylor, S.S. Bub1 maintains centromeric cohesion by activation of the spindle checkpoint. Dev. Cell 2007, 13, 566–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, P.; Maure, J.F.; Javerzat, J.P. Fission yeast Bub1 is essential in setting up the meiotic pattern of chromosome segregation. Nat. Cell. Biol. 2001, 3, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, T.S.; Kawashima, S.A.; Watanabe, Y. The conserved kinetochore protein shugoshinprotects centromeric cohesion during meiosis. Nature 2004, 427, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Steuerwald, N.M.; Bermúdez, M.G.; Wells, D.; Munne, S.; Cohen, J. Maternal age-related differential global expression profiles observed in human oocytes. Reprod. BioMed. Online. 2007, 14, 700–708. [Google Scholar] [CrossRef]
- Pan, H.; Ma, P.P.; Zhu, W.T.; Schultz, R.M. Age-associated increase in aneuploidy and changes in gene expression in mouse eggs. Dev. Biol. 2008, 316, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Steuerwald, N.; Cohen, J.; Herrera, R.J.; Sandalinas, M.; Brenner, C.A. Association between spindle assembly checkpoint expression and maternal age in human oocytes. Mol. Hum. Reprod. 2001, 7, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Holt, J.E.; Lane, S.I.; McLaughlin, E.A.; Merriman, J.A.; Jones, K.T. Reduced ability to recover from spindle disruption and loss of kinetochore spindle assembly checkpoint proteins in oocytes from aged mice. Cell Cycle 2014, 13, 1938–1947. [Google Scholar] [CrossRef] [PubMed]
- Lagirand-Cantaloube, J.; Ciabrini, C.; Charrasse, S.; Ferrieres, A.; Castro, A.; Anahory, T.; Lorca, T. Loss of Centromere Cohesion in Aneuploid Human Oocytes Correlates with Decreased Kinetochore Localization of the Sac Proteins Bub1 and Bubr1. Sci. Rep. 2017, 7, 44001. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zhou, C.; Cui, Z.; Dai, X.; Zhang, M.; Lu, Y.; Xiong, B. Smc1β is required for activation of SAC during mouse oocyte meiosis. Cell Cycle 2017, 16, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Nabti, I.; Grimes, R.; Sarna, H.; Marangos, P.; Carroll, J. Maternal age-dependent APC/C-mediated decrease in securin causes premature sister chromatid separation in meiosis II. Nat. Commun. 2017, 8, 15346. [Google Scholar] [CrossRef]
- Lee, J.; Kitajima, T.S.; Tanno, Y.; Yoshida, K.; Morita, T.; Miyano, T.; Miyake, M.; Watanabe, Y. Unified mode of centromeric protection by shugoshin in mammalian oocytes and somatic cells. Nat. Cell. Biol. 2008, 10, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y. Shugoshin: Guardian spirit at the centromere. Curr. Opin. Cell. Biol. 2005, 17, 590–595. [Google Scholar] [CrossRef]
- Llano, E.; Gomez, R.; Gutierrez-Caballero, C.; Herran, Y.; Sanchez-Martin, M.; Vazquez-Quinones, L.; Hernandez, T.; de Alava, E.; Cuadrado, A.; Barbero, J.L.; et al. Shugoshin-2 is essential for the completion of meiosis but not for mitotic cell division in mice. Genes. Dev. 2008, 22, 2400–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera, T.; Losada, A. Shugoshin and PP2A, shared duties at the centromere. BioEssays 2006, 8, 775–779. [Google Scholar] [CrossRef]
- Kitajima, T.S.; Sakuno, T.; Ishiguro, K.; Iemura, S.; Natsume, T.; Kawashima, S.A.; Watanabe, Y. Shugoshin collaborates with protein phosphatase 2A to protect cohesin. Nature 2006, 441, 46–52. [Google Scholar] [CrossRef]
- Yun, Y.; Lane, S.I.; Jones, K.T. Premature dyad separation in meiosis II is the major segregation error with maternal age in mouse oocytes. Development 2014, 141, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Hamant, O.; Golubovskaya, I.; Meeley, R.; Fiume, E.; Timofejeva, L.; Schleiffer, A.; Nasmyth, K.; Cande, W.Z. A REC8-dependent plant Shugoshin is required for maintenance of centromeric cohesion during meiosis and has no mitotic functions. Curr. Biol. 2005, 15, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, R.; Gillespie, P.J.; Hirano, T. Human Wapl is a cohesin-binding protein that promotes sister-chromatid resolution in mitotic prophase. Curr. Biol. 2006, 24, 2406–2417. [Google Scholar] [CrossRef]
- Kueng, S.; Hegemann, B.; Peters, B.H.; Lipp, J.J.; Schleiffer, A.; Mechtler, K.; Peters, J.M. Wapl controls the dynamic association of cohesin with chromatin. Cell 2006, 127, 955–967. [Google Scholar] [CrossRef]
- Rankin, S.; Ayad, N.G.; Kirschner, M.W. Sororin, a substrate of the anaphase-promoting complex, is required for sister chromatid cohesion in vertebrates. Mol. Cell 2005, 18, 185–200. [Google Scholar] [CrossRef]
- Lafont, A.L.; Song, J.; Rankin, S. Sororin cooperates with the acetyltransferase Eco2 to ensure DNA replication-dependent sister chromatid cohesion. Proc. Natl. Acad. Sci. USA 2010, 107, 20364–20369. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T.; Ladurner, R.; Schmitz, J.; Kreidl, E.; Schleiffer, A.; Bhaskara, V.; Bando, M.; Shirahige, K.; Hyman, A.A.; Mechtler, K.; et al. Sororin mediates sister chromatid cohesion by antagonizing Wapl. Cell 2010, 143, 737–749. [Google Scholar] [CrossRef]
- Hara, K.; Zheng, G.; Qu, Q.; Liu, H.; Ouyang, Z.; Chen, Z.; Tomchick, D.R.; Yu, H. Structure of cohesin subcomplex pinpoints direct shugoshin-Wapl antagonism in centromeric cohesion. Nat. Struct. Mol. Biol. 2014, 21, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Tarín, J.J. Potential effects of age-associated oxidative stress on mammalian oocytes/embryos. Mol. Hum. Reprod. 1996, 2, 717–724. [Google Scholar] [CrossRef]
- Tian, L.; Cai, Q.; Wei, H. Alterations of antioxidant enzymes and oxidative damage to macromolecules in different organs of rats during aging. Free. Radic. Biol. Med. 1998, 24, 1477–1484. [Google Scholar] [CrossRef]
- Martin, I.; Grotewiel, M.S. Oxidative damage and age-related functional declines. Mech. Ageing Dev. 2006, 127, 411–423. [Google Scholar] [CrossRef]
- Muller, F.L.; Lustgarten, M.S.; Jang, Y.; Richardson, A.; Van Remmen, H. Trends in oxidative aging theories. Free Radic. Biol. Med. 2007, 43, 477–503. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Aponte-Mellado, A.; Premkumar, B.J.; Shaman, A.; Gupta, S. The effects of oxidative stress on female reproduction: A review. Reprod. Biol. Endocrinol. 2012, 10, 49. [Google Scholar] [CrossRef]
- Devine, P.J.; Perreault, S.D.; Luderer, U. Roles of reactive oxygen species and antioxidants in ovarian toxicity. Biol. Reprod. 2012, 27, 1–10. [Google Scholar]
- Tarín, J.J.; Vendrell, F.J.; Ten, J.; Cano, A. Antioxidant therapy counteracts the disturbing effects of diamide and maternal ageing on meiotic division and chromosomal segregation in mouse oocytes. Mol. Hum. Reprod. 1998, 4, 281–288. [Google Scholar] [CrossRef]
- Perkins, A.T.; Das, T.M.; Panzera, L.C.; Bickel, S.E. Oxidative stress in oocytes during midprophase induces premature loss of cohesion and chromosome segregation errors. Proc. Natl. Acad. Sci. 2016, 113, E6823–E6830. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Boukari, H.; Banerjee, I.; Sbalzarini, I.F.; Horvath, P.; Helenius, A. Histone deacetylase 8 is required for centrosome cohesion andinfluenza A virus entry. PLoS Pathog. 2011, 7, e1002316. [Google Scholar] [CrossRef] [PubMed]
- Deardoff, M.A.; Bando, M.; Nakato, R.; Watrin, E.; Itoh, T.; Minamino, M.; Saitoh, K.; Komata, M.; Katou, Y.; Clark, D. HDAC8 mutations in Cornelia de Lange syndrome affect the cohesin acetylation cycle. Nature 2012, 489, 313–317. [Google Scholar] [CrossRef]
- Nasmyth, K. Cohesin: A catenase with separate entry and exitgates? Nat. Cell. Biol. 2011, 13, 1170–1177. [Google Scholar] [CrossRef]
- Carpenter, A.T.C. Chiasma function. Cell 1994, 7, 959–962. [Google Scholar] [CrossRef]
- Martinez-Perez, E.; Schvarzstein, M.; Barroso, C.; Lightfoot, J.; Dernburg, A.F.; Villeneuve, A.M. Crossovers trigger a remodeling of meiotic chromosome axis composition that is linked to two-step loss of sister chromatid cohesion. Genes Dev. 2008, 22, 2886–2901. [Google Scholar] [CrossRef] [PubMed]
- Bickel, S.E.; Orr-Weaver, T.L.; Balicky, E.M. The sister-chromatid cohesion protein ORD is required for chiasma maintenance in Drosophila oocytes. Curr. Biol. 2002, 12, 925–929. [Google Scholar] [CrossRef]
- Henderson, S.A.; Edwards, R.G. Chiasma frequency and maternal age in mammals. Nature 1968, 218, 22–28. [Google Scholar] [CrossRef]
- Nilsson, N.O.; Sall, T. A model of chiasma reduction of closely formed crossovers. J. Theor. Biol. 1995, 7, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Broman, K.W.; Murray, J.C.; Sheffield, V.C.; White, R.L.; Weber, J.L. Comprehensive human genetic maps: Individual and sex-specific variation in recombination. Am. J. Hum. Genet. 1998, 63, 861–869. [Google Scholar] [CrossRef]
- Gruhn, J.R.; Al-Asmar, N.; Fasnacht, R.; Maylor-Hagen, H.; Peinado, V.; Rubio, C.; Broman, K.W.; Hunt, P.A.; Hassold, T. Correlations between synaptic initiation and meiotic recombination: A study of humans and mice. Am. J. Hum. Genet. 2016, 98, 102–115. [Google Scholar] [CrossRef]
- Lamb, N.E.; Feingold, E.; Savage, A.; Avramopoulos, D.; Freeman, S.; Gu, Y.C.; Hallberg, A.; Hersey, J.; Karadima, G.; Pettay, D.; et al. Characterization of susceptible chiasma configurations that increase the risk for maternal nondisjunction of chromosome 21. Hum. Mol. Genet. 1997, 6, 1391–1399. [Google Scholar] [CrossRef]
- Oliver, T.R.; Tinker, S.W.; Allen, E.G.; Hollis, N.; Locke, A.E.; Bean, L.J.; Chowdhury, R.; Begum, F.; Marazita, M.; Cheung, V.; et al. Altered patterns of multiple recombinant events are associated with nondisjunction of chromosome 21. Hum. Genet. 2012, 13, 1039–1046. [Google Scholar] [CrossRef]
- Orr-Weaver, T. Meiotic nondisjunction does the two-step. Nat Genet. 1996, 14, 374–376. [Google Scholar] [CrossRef]
- LeMaire-Adkins, R.; Radke, K.; Hunt, P.A. Lack of checkpoint control at the metaphase/anaphase transition: A mechanism of meiotic nondisjunction in mammalian females. J. Cell. Biol. 1997, 139, 1611–1619. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, T.S.; Ohsugi, M.; Ellenberg, J. Complete kinetochore tracking reveals error-prone homologous chromosome biorientation in mammalian oocytes. Cell 2011, 146, 568–581. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y. Geometry and force behind kinetochore orientation: Lessons from meiosis. Nat Rev. Mol. Cell. Biol. 2012, 13, 370–382. [Google Scholar] [CrossRef]
- Wittmann, T.; Hyman, A.; Desai, A. The spindle: A dynamic assembly of microtubules and motors. Nat. Cell. Biol. 2001, 3, E28–E34. [Google Scholar] [CrossRef]
- Salmon, E.D.; Cimini, D.; Cameron, L.A.; Deluca, J.G. Merotelic kinetochores in mammalian tissue cells. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2005, 360, 553–568. [Google Scholar] [CrossRef]
- Cimini, D.; Howell, B.; Maddox, P.; Khodjakov, A.; Degrassi, F.; Salmon, E.D. Merotelic kinetochore orientation is a major mechanism of aneuploidy in mitotic mammalian tissue cells. J. Cell. Biol. 2001, 153, 517–527. [Google Scholar] [CrossRef]
- Gregan, J.; Polakova, S.; Zhang, L.J.; Tolic´-Nørrelykke, I.M.; Cimini, D. Merotelic kinetochore attachment: Causes and effects. Trends Cell Biol. 2011, 21, 374–381. [Google Scholar]
- Cimini, D.; Fioravanti, D.; Salmon, E.D.; Degrassi, F. Merotelic kinetochore orientation versus chromosome mono-orientation in the origin of lagging chromosomes in human primary cells. J. Cell. Sci. 2002, 115, 507–515. [Google Scholar]
- Cheng, J.M.; Li, J.; Tang, J.X.; Hao, X.X.; Wang, Z.P.; Sun, T.C.; Wang, X.X.; Zhang, Y.; Chen, S.R.; Liu, Y.X. Merotelic kinetochore attachment in oocyte meiosis II causes sister chromatids segregation errors in aged mice. Cell Cycle 2017. accepted. [Google Scholar]
- Kouznetsova, A.; Hernandez-Hernandez, A.; Hoog, C. Merotelic attachments allow alignment and stabilization of chromatids in meiosis II oocytes. Nat. Commun. 2014, 5, 4409. [Google Scholar] [CrossRef] [PubMed]
- Bakhoum, S.F.; Genovese, G.; Compton, D.A. Deviant kinetochore microtubule dynamics underlie chromosomal instability. Curr. Biol. 2009, 19, 1937–1942. [Google Scholar] [CrossRef]
- Barber, T.D.; McManus, K.; Yuen, K.W.; Reis, M.; Parmigiani, G.; Shen, D.; Barrett, I.; Nouhi, Y.; Spencer, F.; Markowitz, S.; et al. Chromatid cohesion defects may underlie chromosome instability in human colorectal cancers. Proc. Natl. Acad. Sci. 2008, 105, 3443–3448. [Google Scholar] [CrossRef]
- Orr, B.; Godek, K.M.; Compton, D. Aneuploidy. Curr. Biol. 2015, 25, R538–R542. [Google Scholar] [CrossRef] [PubMed]
- Duncan, F.E.; Hornick, J.E.; Lampson, M.A.; Schultz, R.M.; Shea, L.D.; Woodruff, T.K. Chromosome cohesion decreases in human eggs with advanced maternal age. Aging Cell 2012, 11, 1121–1124. [Google Scholar] [CrossRef] [PubMed]
- Selesniemi, K.; Lee, H.J.; Tilly, J.L. Moderate caloric restriction initiated in rodents during adulthood sustains function of the female reproductive axis into advanced chronological age. Aging Cell 2008, 7, 622–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selesniemi, K.; Lee, H.J.; Muhlhauser, A.; Tilly, J.L. Prevention of maternal aging-associated oocyte aneuploidy and meiotic spindle defects in mice by dietary and genetic strategies. Proc. Natl. Acad. Sci. 2011, 108, 12319–12324. [Google Scholar] [CrossRef]
- Wang, T.; Gao, Y.Y.; Chen, L.; Nie, Z.W.; Cheng, W.; Liu, X.; Schatten, H.; Zhang, X.; Miao, Y.L. Melatonin prevents postovulatory oocyte aging and promotes subsequent embryonic development in the pig. Aging 2017, 9, 1552–1564. [Google Scholar] [CrossRef] [PubMed]
- Lord, T.; Nixon, B.; Jones, K.T.; Aitken, R.J. Melatonin prevents postovulatory oocyte aging in the mouse and extends the window for optimal fertilization in vitro. Biol. Reprod. 2013, 88, 67. [Google Scholar] [CrossRef]
- Park, Y.S.; You, S.Y.; Cho, S.; Jeon, H.J.; Lee, S.; Cho, D.H.; Kim, J.S.; Oh, J.S. Eccentric localization of catalase to protect chromosomes from oxidative damages during meiotic maturation in mouse oocytes. Histochem. Cell Boil. 2016, 146, 281–288. [Google Scholar] [CrossRef] [PubMed]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, J.-M.; Liu, Y.-X. Age-Related Loss of Cohesion: Causes and Effects. Int. J. Mol. Sci. 2017, 18, 1578. https://doi.org/10.3390/ijms18071578
Cheng J-M, Liu Y-X. Age-Related Loss of Cohesion: Causes and Effects. International Journal of Molecular Sciences. 2017; 18(7):1578. https://doi.org/10.3390/ijms18071578
Chicago/Turabian StyleCheng, Jin-Mei, and Yi-Xun Liu. 2017. "Age-Related Loss of Cohesion: Causes and Effects" International Journal of Molecular Sciences 18, no. 7: 1578. https://doi.org/10.3390/ijms18071578
APA StyleCheng, J. -M., & Liu, Y. -X. (2017). Age-Related Loss of Cohesion: Causes and Effects. International Journal of Molecular Sciences, 18(7), 1578. https://doi.org/10.3390/ijms18071578