Alteration of Mammary Gland Development and Gene Expression by In Utero Exposure to Cadmium

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Effects of Early Life Exposure to Cadmium on Vaginal Opening in Female Offspring
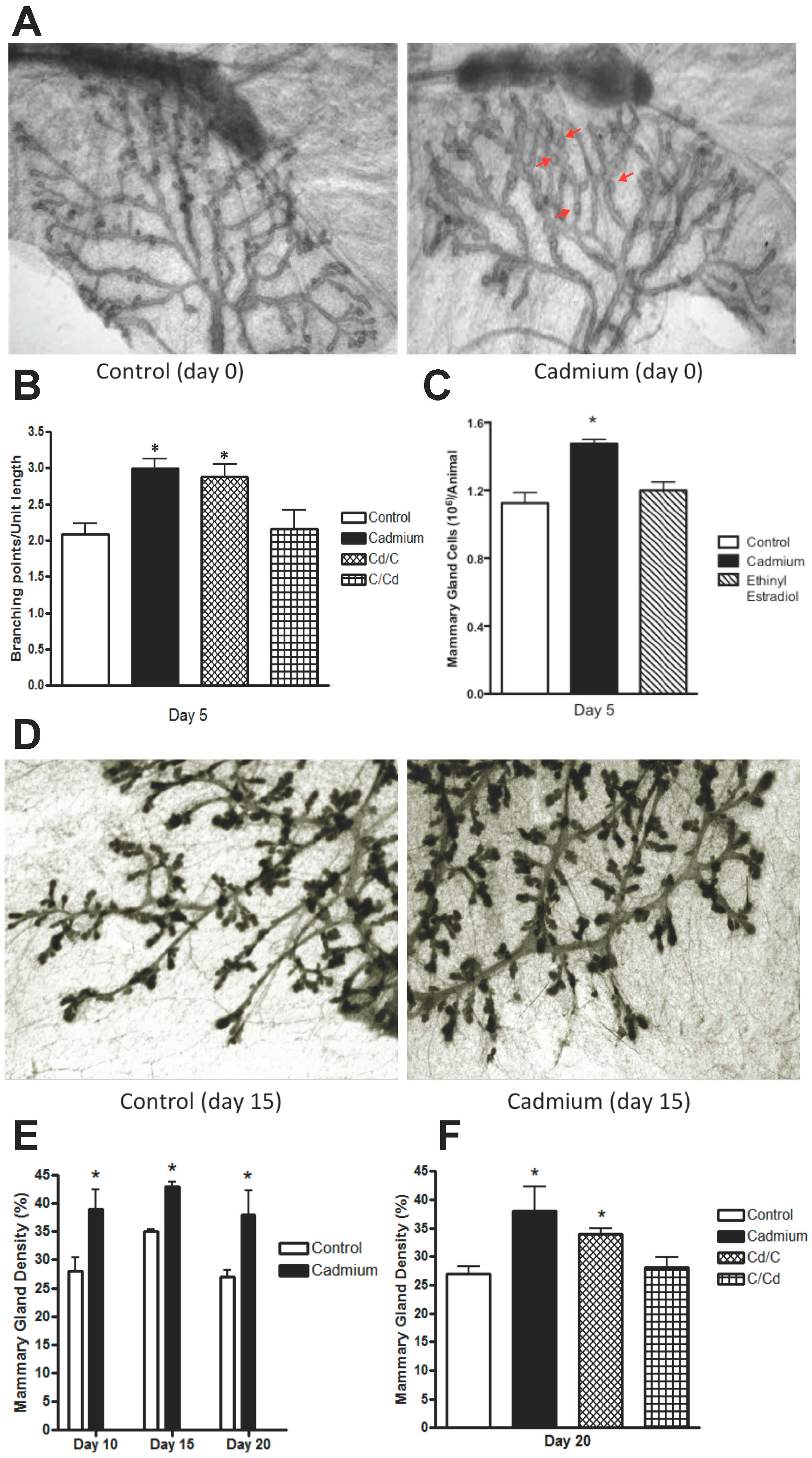
2.2. Effects of In Utero Exposure to Cadmium on the Morphology of the Mammary Gland of Female Offspring
2.3. Effects of In Utero Exposure to Cadmium on Mammosphere-Forming Cells in the Mammary Glands of Female Offspring
2.4. Effects of In Utero Exposure to Cadmium on Gene Expression in the Mammary Gland of Female Offspring
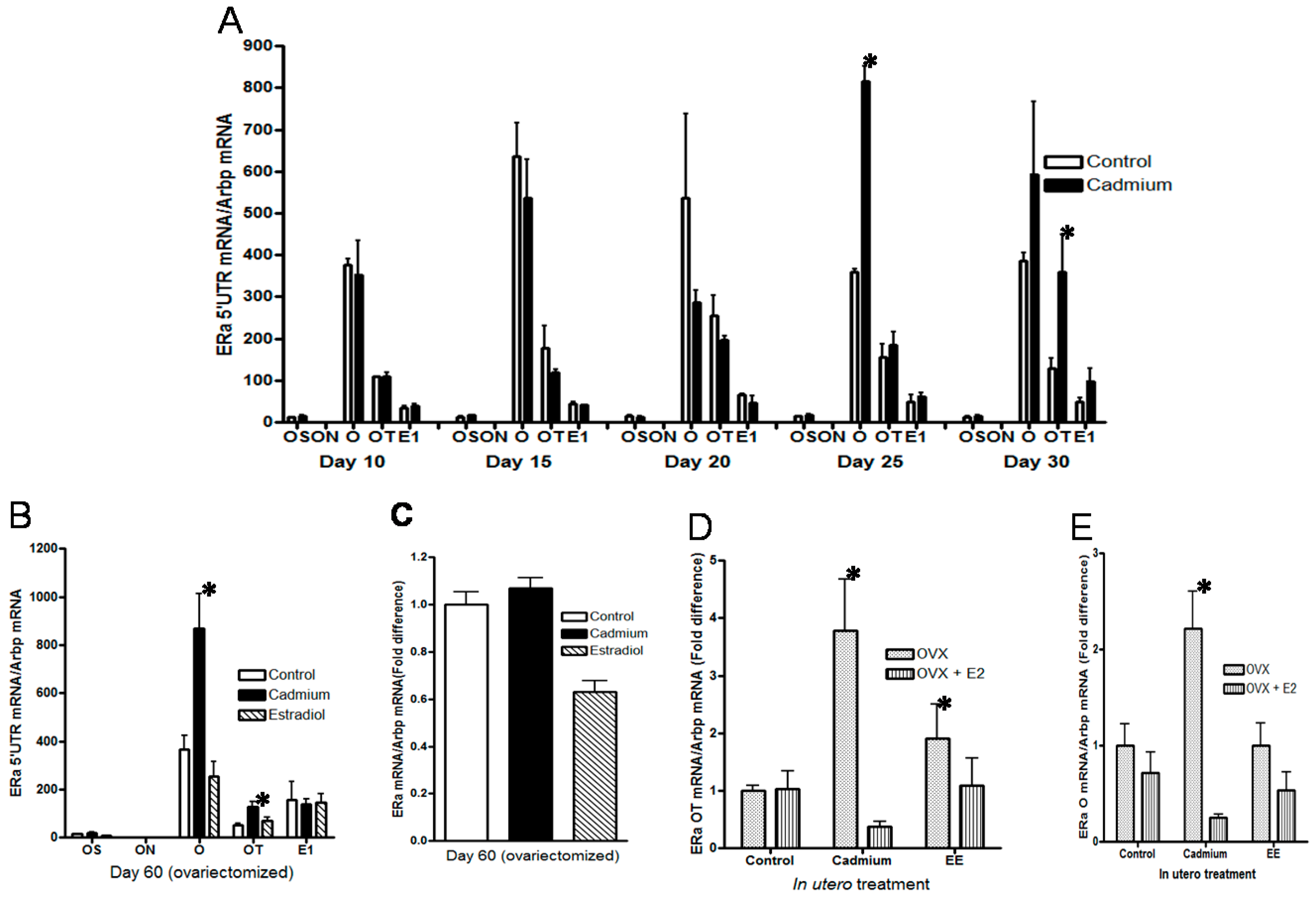
2.5. Effects of In Utero Exposure to Cadmium on the Expression of ERα Transcripts in the Mammary Gland of Female Offspring
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Morphological Analysis of the Mammary Gland
4.3. Mammosphere Culture
4.4. Real Time Reverse Transcriptase-Polymerase Chain Reaction
4.5. Nuclear Run-On Assay
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Delemarre, E.M.; Felius, B.; Delemarre-van de Waal, H.A. Inducing puberty. Eur. J. Endocrinol. 2008, 159, S9–S15. [Google Scholar] [CrossRef] [PubMed]
- Colborn, T.; von Saal, F.S.; Soto, A.M. Developmental effects of endocrine-disrupting chemicals in wildlife and humans. Environ. Health Perspect. 1993, 101, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Fenton, S.E.; Reed, C.; Newbold, R.R. Perinatal environmental exposures affect mammary development, function, and cancer risk in adulthood. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 455–479. [Google Scholar] [CrossRef] [PubMed]
- De Muinck Keizer, S.M.; Mul, D. Trends in pubertal development in Europe. Hum. Reprod. Update 2001, 7, 287–291. [Google Scholar] [CrossRef]
- Herman-Giddens, M.E.; Slora, E.J.; Wasserman, R.C.; Bourdony, C.J.; Bhapkar, M.V.; Koch, G.G.; Hasemeier, C.M. Secondary sexual characteristics and menses in young girls seen in office practice: A study from the Pediatric Research in Office Settings network. Pediatrics 1997, 99, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Delemarre-van de Waal, H.A. Regulation of puberty. Best Pract. Res. Clin. Endocrinol. Metab. 2002, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hens, J.R.; Wysolmerski, J.J. Key stages of mammary gland development: Molecular mechanisms involved in the formation of the embryonic mammary gland. Breast Cancer Res. 2005, 7, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Fendrick, J.L.; Raafat, A.M.; Haslam, S.Z. Mammary gland growth and development from the postnatal period to postmenopause: Ovarian steroid receptor ontogeny and regulation in the mouse. J. Mammary Gland Biol. Neoplasia 1998, 3, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Morales, P.; Saceda, M.; Kenney, N.; Kim, N.; Salomon, D.S.; Gottardis, M.M.; Solomon, H.B.; Sholler, P.F.; Jordan, V.C.; Martin, M.B. Effect of cadmium on estrogen receptor levels and estrogen-induced responses in human breast cancer cells. J. Biol. Chem. 1994, 269, 16896–16901. [Google Scholar] [PubMed]
- Stoica, A.; Katzenellenbogen, B.S.; Martin, M.B. Activation of estrogen receptor-α by the heavy metal cadmium. Mol. Endocrinol. 2000, 14, 545–553. [Google Scholar] [PubMed]
- Stoica, A.; Pentecost, E.; Martin, M.B. Effect of arsenite on estrogen receptor-a expression and activity in MCF-7 breast cancer cells. Endocrinology 2000, 141, 3595–3602. [Google Scholar] [CrossRef] [PubMed]
- Stoica, A.; Pentecost, E.; Martin, M.B. Effects of selenite on estrogen receptor-a expression and activity in MCF-7 breast cancer cells. J. Cell. Biochem. 2000, 79, 282–292. [Google Scholar] [CrossRef]
- Veselik, D.J.; Divekar, S.; Dakshanamurthy, S.; Storchan, G.B.; Turner, J.M.; Graham, K.L.; Huang, L.; Stoica, A.; Martin, M.B. Activation of estrogen receptor-α by the anion nitrite. Cancer Res. 2008, 68, 3950–3958. [Google Scholar] [CrossRef] [PubMed]
- Divekar, S.D.; Storchan, G.B.; Sperle, K.; Veselik, D.J.; Johnson, E.; Dakshanamurthy, S.; Lajiminmuhip, Y.N.; Nakles, R.E.; Huang, L.; Martin, M.B. The role of calcium in the activation of estrogen receptor-α. Cancer Res. 2011, 71, 1658–1668. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.B.; Reiter, R.; Pham, T.; Avellanet, Y.R.; Camara, J.; Lahm, M.; Pentecost, E.; Pratap, K.; Gilmore, B.; Divekar, S.; et al. Estrogen like activity of metals in MCF-7 breast cancer cells. Endocrinology 2003, 144, 2425–2436. [Google Scholar] [CrossRef] [PubMed]
- Freni-Titulaer, L.W.; Cordero, J.F.; Haddock, L.; Lebron, G.; Martinez, R.; Mills, J.L. Premature thelarche in Puerto Rico. A search for environmental factors. Am. J. Dis. Child. 1986, 140, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Blanck, H.M.; Marcus, M.; Tolbert, P.E.; Rubin, C.; Henderson, A.K.; Hertzberg, V.S.; Zhang, R.H.; Cameron, L. Age at menarche and tanner stage in girls exposed in utero and postnatally to polybrominated biphenyl. Epidemiology 2000, 11, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L.; Cho, E.; Onojafe, I.; Raygada, M.; Clarke, R. Maternal exposure to genistein during pregnancy increases carcinogen-induced mammary tumorigenesis in female rat offspring. Oncol. Rep. 1999, 6, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L.; Cho, E.; Clarke, R. Maternal genistein exposure mimics the effects of estrogen on mammary gland development in female mouse offspring. Oncol. Rep. 1998, 5, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.D.; Kenney, N.; Stoica, A.; Hilakivi-Clarke, L.; Singh, B.; Chepko, G.; Clarke, R.; Sholler, P.F.; Lirio, A.A.; Foss, C.; et al. Cadmium mimics the in vivo effects of estrogen in the uterus and mammary gland. Nat. Med. 2003, 9, 1081–1084. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.Y.; Kim, S.J.; Kim, H.G.; Lee, J.H.; Choi, Y.; Lee, H.; Kim, Y. Evaluation of estrogenicity of major heavy metals. Sci. Total Environ. 2003, 312, 15–21. [Google Scholar] [CrossRef]
- Martinez-Campa, C.; Alonso-Gonzalez, C.; Mediavilla, M.D.; Cos, S.; Gonzalez, A.; Ramos, S.; Sanchez-Barcelo, E.J. Melatonin inhibits both ERα activation and breast cancer cell proliferation induced by a metalloestrogen, cadmium. J. Pineal Res. 2006, 40, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Siewit, C.L.; Gengler, B.; Vegas, E.; Puckett, R.; Louie, M.C. Cadmium promotes breast cancer cell proliferation by potentiating the interaction between ERα and c-Jun. Mol. Endocrinol. 2010, 24, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Brama, M.; Gnessi, L.; Basciani, S.; Cerulli, N.; Politi, L.; Spera, G.; Mariani, S.; Cherubini, S.; d’Abusco, A.S.; Scandurra, R.; et al. Cadmium induces mitogenic signaling in breast cancer cell by an ERα -dependent mechanism. Mol. Cell. Endocrinol. 2007, 264, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Wilson, V.S.; Bobseine, K.; Gray, L.E. Development and characterization of a cell line that stably expresses an estrogen-responsive luciferase reporter for the detection of estrogen receptor agonist and antagonists. Toxicol. Sci. 2004, 81, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yu, X.; Shaikh, Z.A. Rapid activation of ERK1/2 and AKT in human breast cancer cells by cadmium. Toxicol. Appl. Pharmacol. 2008, 228, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Odwin-Dacosta, S.; Yager, J.D. Effects of cadmium on estrogen receptor mediated signaling and estrogen induced DNA synthesis in T47D human breast cancer cells. Toxicol. Lett. 2009, 184, 134–138. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Cadmium and cadmium compounds. In International Agency for Research on Cancer Monographs; IARC Press: Lyon, France, 1993; Volume 58, pp. 119–238. [Google Scholar]
- Gartell, M.J.; Craun, J.C.; Podrebarae, D.S.; Gunderson, E.R. Pesticides, selected elements and other chemicals in adult total diet samples. October 1980–March 1982. J. Assoc. Anal. Chem. 1986, 69, 146–161. [Google Scholar]
- Gartell, M.J.; Craun, J.C.; Podrebarae, D.S.; Gunderson, E.R. Pesticides, selected elements and other chemicals in infant and toddler total diet samples. October 1980–March 1982. J. Assoc. Anal. Chem. 1986, 69, 123–145. [Google Scholar]
- Jarup, L.; Berglund, M.; Elinder, C.G.; Nordberg, G.; Vahter, M. Health effects of cadmium exposure—A review of the literature and a risk estimate. Scand. J. Work Environ. Health 1998, 24, 1–51. [Google Scholar] [PubMed]
- Ysart, G.; Miller, P.; Croasdale, M.; Crews, H.; Robb, P.; Baxter, M.; De L’Argy, C.; Harrison, N. 1997 UK Total Diet Study—Dietary exposures to aluminium, arsenic, cadmium, chromium, copper, lead, mercury, nickel, selenium, tin and zinc. Food Addit. Contam. 2000, 17, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Moschandreas, D.J.; Karuchit, S.; Berry, M.R.; O’Rourke, M.K.; Lo, D.; Lebowitz, M.D.; Robertson, G. Exposure apportionment: Ranking food items by their contribution to dietary exposure. J. Expo. Anal. Environ. Epidemiol. 2002, 12, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.; Anke, M.; Illing-Gunther, H.; Thiel, C. Oral cadmium exposure of adults in Germany. 2: Market basket calculations. Food Addit. Contam. 1998, 15, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, M.; Wittsiepe, J.; Schrey, P.; Budde, U.; Idel, H. Dietary intake of cadmium by children and adults from Germany using duplicate portion sampling. Sci. Total Environ. 2002, 285, 11–19. [Google Scholar] [CrossRef]
- Dabeka, R.W.; McKenzie, A.D. Lead and Cadmium levels in commercial infant foods and dietary intake by infants 0–1 year old. Food Addit. Contam. 1988, 5, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Sternowsky, H.J.; Wessolowske, R. Lead and cadmium in breast milk. Higher levels in urban vs. rural mothers during the first 3 months of lactation. Arch. Toxicol. 1985, 57, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Turan, S.; Saygi, S.; Kilic, Z.; Acar, O. Determination of heavy metal contents in human colostrum samples by electrothermal atomic absorption spectroscopy. J. Trop. Pediatr. 2001, 47, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, M.; Finch, H. Cadmium content of umbilical cord blood. Environ. Res. 1984, 34, 120–122. [Google Scholar] [CrossRef]
- Honda, R.; Tawara, K.; Nishijo, M.; Nakagawa, H.; Tanebe, K.; Saito, S. Cadmium exposure and trace elements in human breast milk. Toxicology 2003, 186, 255–259. [Google Scholar] [CrossRef]
- Petersson, G.K.; Oskarsson, A. Cadmium in milk and mammary gland in rats and mice. Arch. Toxicol. 2000, 73, 519–527. [Google Scholar] [CrossRef]
- Korpela, H.; Loueniva, R.; Yrjanheikki, E.; Kauppila, A. Lead and cadmium concentrations in maternal and umbilical cord blood, aminiotic fluid, placenta, and amniotic membranes. Am. J. Obstet. Gynecol. 1986, 155, 1086–1089. [Google Scholar] [CrossRef]
- Cantor, K.P.; Stewart, P.A.; Brinton, L.A.; Dosemeci, M. Occupational exposures and female breast cancer mortality in the United States. J. Occup. Med. 1994, 37, 336–348. [Google Scholar] [CrossRef]
- Ursin, G.; Bernstein, L.; Pike, M.C. Breast cancer. In Cancer Surveys; Dol, R., Fraumeni, J.F., Eds.; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1994; pp. 241–264. [Google Scholar]
- McElroy, J.A.; Shafer, M.M.; Trentham-Dietz, A.; Hampton, J.M.; Newcomb, P.A. Cadmium exposure and breast cancer risk. J. Natl. Cancer Inst. 2006, 98, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, C.M.; Chen, J.J.; Kovach, J.S. Environmental cadmium and breast cancer risk. Aging (Albany NY) 2010, 2, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, I.; Clayton, P. The genetic basis for the timing of human puberty. J. Neuroendocrinol. 2007, 19, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Parodi, D.A.; Greenfield, M.; Evans, C.; Chichura, A.; Alpaugh, A.; Williams, J.; Martin, M.B. Alteration of mammary gland development and gene expression by in utero exposure to arsenic. Reprod. Toxicol. 2015, 54, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Welm, B.E.; Freeman, K.W.; Chen, M.; Contreras, A.; Spencer, D.M.; Rosen, J.M. Inducible dimerization of FGFR1: Development of a mouse model to analyze progressive transformation of the mammary gland. J. Cell Biol. 2002, 157, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Kenney, N.J.; Smith, G.H.; Lawrence, E.; Barrett, J.C.; Salomon, D.S. Identification of Stem Cell Units in the Terminal End Bud and Duct of the Mouse Mammary Gland. J. Biomed. Biotechnol. 2001, 1, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honeth, G.; Lombardi, S.; Ginestier, C.; Hur, M.; Marlow, R.; Buchupalli, B.; Shinomiya, I.; Gazinska, P.; Bombelli, S.; Ramalingam, V.; et al. Aldehyde dehydrogenase and estrogen receptor define a hierarchy of cellular differentiation in the normal human mammary epithelium. Breast Cancer Res. 2014, 16, R52. [Google Scholar] [CrossRef] [PubMed]
- Dontu, G.; Abdallah, W.M.; Foley, J.M.; Jackson, K.W.; Clarke, M.F.; Kawamura, M.J.; Wicha, M.S. In vitro propagation and transcriptional profiling of human mammary stem/progenitor cells. Genes Dev. 2003, 17, 1253–1270. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.; Vlachou, T.; Carminati, M.; Pelicci, P.G.; Mapelli, M. Molecular mechanisms of asymmetric divisions in mammary stem cells. EMBO Rep. 2016, 17, 1700–1720. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.B. Isolation and characterization of human mammary stem cells. Cell Prolif. 2005, 38, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Stingl, J. Detection and analysis of mammary gland stem cells. J. Pathol. 2009, 217, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Ma, I.; Allan, A.L. The role of human aldehyde dehydrogenase in normal and cancer stem cells. Stem Cell Rev. 2011, 7, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Visvader, J.E. Keeping abreast of the mammary epithelial hierarchy and breast tumorigenesis. Genes Dev. 2009, 23, 2563–2577. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.B.; Spence, K.; Anderson, E.; Howell, A.; Okano, H.; Potten, C.S. A putative human breast stem cell population is enriched for steroid receptor-positive cells. Dev. Biol. 2005, 277, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Booth, B.W.; Smith, G.H. Estrogen receptor-α and progesterone receptor are expressed in label-retaining mammary epithelial cells that divide asymmetrically and retain their template DNA strands. Breast Cancer Res. 2006, 8, R49. [Google Scholar] [CrossRef] [PubMed]
- Stingl, J. Estrogen and progesterone in normal mammary gland development and in cancer. Horm. Cancer 2011, 2, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Hiyama, E.; Hiyama, K. Telomere and telomerase in stem cells. Br. J. Cancer 2007, 96, 1020–1024. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.; Saxena, M.; Paranjape, A.N.; Krishnan, V.; Giraddi, R.; Kumar, M.V.; Mukherjee, G.; Rangarajan, A. Phenotypic and functional characterization of human mammary stem/progenitor cells in long term culture. PLoS ONE 2009, 4, e5329. [Google Scholar] [CrossRef] [PubMed]
- McCoy, E.L.; Iwanaga, R.; Jedlicka, P.; Abbey, N.S.; Chodosh, L.A.; Heichman, K.A.; Welm, A.L.; Ford, H.L. Six1 expands the mouse mammary epithelial stem/progenitor cell pool and induces mammary tumors that undergo epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 2663–2677. [Google Scholar] [CrossRef] [PubMed]
- Coletta, R.D.; McCoy, E.L.; Burns, V.; Kawakami, K.; McManaman, J.L.; Wysolmerski, J.J.; Ford, H.L. Characterization of the Six1 homeobox gene in normal mammary gland morphogenesis. BMC Dev. Biol. 2010, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Ford, H.L.; Kabingu, E.N.; Bump, E.A.; Mutter, G.L.; Pardee, A.B. Abrogation of the G2 cell cycle checkpoint associated with overexpression of HSIX1: A possible mechanism of breast carcinogenesis. Proc. Natl. Acad. Sci. USA 1998, 95, 12608–12613. [Google Scholar] [CrossRef] [PubMed]
- El Sheikh, S.H.; Toullec, A.; Vacher, S.; Pocard, M.; Bieche, I.; Perrot-Applanat, M. In utero and lactational exposure to vinclozolin and genistein induces genomic changes in the rat mammary gland. J. Endocrinol. 2013, 216, 245–263. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Davison, A.J.; Tsang, S.S. Vanadate-induced gene expression in mouse C127 cells: Roles of oxygen derived active species. Mol. Cell. Biochem. 1992, 115, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Barthold, J.S.; Kryger, J.V.; Derusha, A.M.; Duel, B.P.; Jednak, R.; Skafar, D.F. Effects of an environmental endocrine disruptor on fetal development, estrogen receptor α and epidermal growth factor receptor expression in the porcine male genital tract. J. Urol. 1999, 162, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.C.; Hudgins, S.; Lewis, A.; Schorr, K.; Sommer, R.; Peterson, R.E.; Flaws, J.A.; Furth, P.A. In utero and lactational treatment with 2,3,7,8-tetrachlorodibenzo-p-dioxin impairs mammary gland differentiation but does not block the response to exogenous estrogen in the postpubertal female rat. Toxicol. Sci. 2001, 62, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Schonfelder, G.; Flick, B.; Mayr, E.; Talsness, C.; Paul, M.; Chahoud, I. In utero exposure to low doses of bisphenol A lead to long-term deleterious effects in the vagina. Neoplasia 2002, 4, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Russo, J.; Ao, X.; Grill, C.; Russo, I.H. Pattern of distribution of cells positive for estrogen receptor α and progesterone receptor in relation to proliferating cells in the mammary gland. Breast Cancer Res. Treat. 1999, 53, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Sagsoz, H.; Ketani, M.A. The role of estrogen receptors, erbB receptors, vascular endothelial growth factor and its receptors, and vascular endothelial growth inhibitor in the development of the rat mammary gland. Growth Factors 2010, 28, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Saceda, M.; Lippman, M.E.; Chambon, P.; Lindsey, R.L.; Ponglikiturongkol, M.; Puente, M.; Martin, M.B. Regulation of the estrogen receptor in MCF-7 cells by estradiol. Mol. Endocrinol. 1988, 2, 1157–1162. [Google Scholar] [CrossRef] [PubMed]
- Saceda, M.; Lippman, M.E.; Lindsey, R.K.; Puente, M.M.M.B. Role of an estrogen receptor-dependent mechanism in the regulation of estrogen receptor mRNA in MCF-7 cells. Mol. Endocrinol. 1989, 3, 1782–1787. [Google Scholar] [CrossRef] [PubMed]
- Saceda, M.; Lindsey, R.K.; Solomon, H.; Angeloni, S.V.; Martin, M.B. Estradiol regulates estrogen receptor mRNA stability. J. Steroid Biochem. Mol. Biol. 1998, 66, 113–120. [Google Scholar] [CrossRef]
- Mohammed, H.; D’Santos, C.; Serandour, A.A.; Ali, H.R.; Brown, G.D.; Atkins, A.; Rueda, O.M.; Holmes, K.A.; Theodorou, V.; Robinson, J.L.; et al. Endogenous purification reveals GREB1 as a key estrogen receptor regulatory factor. Cell Rep. 2013, 3, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.G.; Thompson, D.A.; Weigel, R.J. PDZK1 and GREB1 are estrogen-regulated genes expressed in hormone-responsive breast cancer. Cancer Res. 2000, 60, 6367–6375. [Google Scholar] [PubMed]
- Kenney, N.J.; Smith, G.H.; Rosenberg, K.; Cutler, M.L.; Dickson, R.B. Induction of ductal morphogenesis and lobular hyperplasia by amphiregulin in the mouse mammary gland. Cell Growth Differ. 1996, 7, 1769–1781. [Google Scholar] [PubMed]
- Luetteke, N.C.; Qiu, T.H.; Fenton, S.E.; Troyer, K.L.; Riedel, R.F.; Chang, A.; Lee, D.C. Targeted inactivation of the EGF and amphiregulin genes reveals distinct roles for EGF receptor ligands in mouse mammary gland development. Development 1999, 126, 2739–2750. [Google Scholar] [PubMed]
- Ciarloni, L.; Mallepell, S.; Brisken, C. Amphiregulin is an essential mediator of estrogen receptor α function in mammary gland development. Proc. Natl. Acad. Sci. USA 2007, 104, 5455–5460. [Google Scholar] [CrossRef] [PubMed]
- Kariagina, A.; Xie, J.; Leipprandt, J.R.; Haslam, S.Z. Amphiregulin mediates estrogen, progesterone, and EGFR signaling in the normal rat mammary gland and in hormone-dependent rat mammary cancers. Horm. Cancer 2010, 1, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Kos, M.; Reid, G.; Denger, S.; Gannon, F. Minireview: Genomic organization of the human ERa gene promoter region. Mol. Endocrinol. 2001, 15, 2057–2063. [Google Scholar] [CrossRef] [PubMed]
- Weigel, R.J.; Crooks, D.L.; Iglehart, J.D.; deConinck, E.C. Quantitative analysis of the transcriptional start sites of estrogen receptor in breast cancer. Cell Growth Differ. 1995, 6, 707–711. [Google Scholar] [PubMed]
- Tanimoto, K.; Eguchi, H.; Yoshida, T.; Hajiro-Nakanishi, K.; Hayashi, S. Regulation of estrogen receptor α gene mediated by promoter B responsible for its enhanced expression in human breast cancer. Nucleic Acids Res. 1999, 27, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Imai, K.; Suga, K.; Kurihara, T.; Higashi, Y.; Nakachi, K. Two promoters in expression of estrogen receptor messenger RNA in human breast cancer. Carcinogenesis 1997, 18, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Monje, L.; Varayoud, J.; Luque, E.H.; Ramos, J.G. Neonatal exposure to bisphenol A modifies the abundance of estrogen receptor α transcripts with alternative 5′-untranslated regions in the female rat preoptic area. J. Endocrinol. 2007, 194, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Hirata, S.; Koh, T.; Yamada-Mouri, N.; Hoshi, K.; Kato, J. The untranslated first exon “exon 0S” of the rat estrogen receptor (ER) gene. FEBS Lett. 1996, 394, 371–373. [Google Scholar] [CrossRef]
- Hirata, S.; Koh, T.; Yamada-Mouri, N.; Kato, J. The novel untranslated first exon “exon 0N” of the rat estrogen receptor gene. Biochem. Biophys. Res. Commun. 1996, 225, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Osada, N.; Hirata, S.; Shoda, T.; Hoshi, K. The novel untranslated exon “exon 0T” encoded between the exon 0 and exon 1 of the rat estrogen receptor α (ERα) gene. Endocr. J. 2001, 48, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed]
- Richie, R.C.; Swanson, J.O. Breast cancer: A review of the literature. J. Insur. Med. 2003, 35, 85–101. [Google Scholar] [PubMed]
- Bray, F.; McCarron, P.; Parkin, D.M. The changing global patterns of female breast cancer incidence and mortality. Breast Cancer Res. 2004, 6, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampert, J.B.; Whittemore, A.S.; Paffenbarger, R.S., Jr. Combined effect of child-bearing, menstrual events, and body size on age-specific breast cancer risk. Am. J. Epidemiol. 1988, 128, 962–979. [Google Scholar] [CrossRef] [PubMed]
- Pike, M.C.; Henderson, B.E.; Casagrande, J.T.; Rosario, I.; Gray, G.E. Oral contraceptive use and early abortion as risk factors for breast cancer in young women. Br. J. Cancer 1981, 43, 72. [Google Scholar] [CrossRef] [PubMed]
- Althuis, M.D.; Dozier, J.M.; Anderson, W.F.; Devesa, S.S.; Brinton, L.A. Global trends in breast cancer incidence and mortality 1973–1997. Int. J. Epidemiol. 2005, 34, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.E.; Ross, R.K.; Judd, H.L.; Krailo, M.D.; Pike, M.C. Do regular ovulatory cycles increase breast cancer risk? Cancer 1985, 56, 1206. [Google Scholar] [CrossRef]
- Hoover, R.; Gray, L.A.S.; Cole, P.; MacMahon, B. Menopausal estrogens and breast cancer. N. Engl. J. Med. 1976, 295, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Hankinson, S.E.; Willett, W.C.; Manson, J.E.; Colditz, G.A.; Hunter, D.J.; Spiegelman, D.; Barbieri, R.L.; Speizer, F.E. Plasma sex steroid hormone levels and risk of breast cancer in postmenopausal women. J. Natl. Cancer Inst. 1998, 90, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Toniolo, P.G.; Levitz, M.; Zeleniuch-Jacquotte, A.; Banerjee, S.; Koenig, K.L.; Shore, R.E.; Strax, P.; Pasternack, B.S. A prospective study of endogenous estrogens and breast cancer in postmenopausal women. J. Natl. Cancer Inst. 1995, 87, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, I.; Vollmar, B.; Runnebaum, B.; Kubli, F. Weight percentile at birth. I. Clinical data of pregnancy and relevance for early childhood development. Eur. J. Obstet. Gynecol. Reprod. Biol. 1987, 26, 303–311. [Google Scholar] [CrossRef]
- Olsen, J.; Storm, H. Pregnancy experience in women who later developed oestrogen-related cancers (Denmark). Cancer Causes Control 1998, 9, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Depue, R.H.; Bernstein, L.; Ross, R.K.; Judd, H.L.; Henderson, B.E. Hyperemesis gravidarum in relation to estradiol levels, pregnancy outcome, and other maternal factors: A seroepidemiologic study. Am. J. Obstet. Gynecol. 1987, 156, 1137–1141. [Google Scholar] [CrossRef]
- Hsieh, C.; Pavia, M.; Lambe, M.; Lan, S.J.; Colditz, G.A.; Ekbom, A.; Adami, H.O.; Trichopoulos, D.; Willett, W.C. Dual effect of parity on breast cancer risk. Eur. J. Cancer 1994, 30A, 969–973. [Google Scholar] [CrossRef]
- Ekbom, A.; Trichopoulos, D.; Adami, H.O.; Hsieh, C.C.; Lan, S.J. Evidence of prenatal influences on breast cancer risk. Lancet 1992, 340, 1015–1018. [Google Scholar] [CrossRef]
- Michels, K.B.; Trichopoulos, D.; Robins, J.M.; Rosner, B.A.; Manson, J.E.; Hunter, D.J.; Colditz, G.A.; Hankinson, S.E.; Speizer, F.E.; Willett, W.C. Birthweight as a risk factor for breast cancer. Lancet 1996, 348, 1542–1546. [Google Scholar] [CrossRef]
- Sanderson, M.; Williams, M.A.; Malone, K.E.; Stanford, J.L.; Emanuel, I.; White, E.; Daling, J.R. Perinatal factors and risk of breast cancer. Epidemiology 1996, 7, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L.; Clarke, R.; Onojafe, I.; Raygada, M.; Cho, E.; Lippman, M.E. A maternal diet high in n-6 polyunsaturated fats alters mammary gland development, puberty onset, and breast cancer risk among female offspring. Proc. Natl. Acad. Sci. USA 1997, 94, 9372–9377. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.E. Tumors of female offspring of mice exposed prenatally to diethylstilbestrol. J. Natl. Cancer Inst. 1984, 73, 133–140. [Google Scholar] [PubMed]
- Russo, I.H.; Russo, J. Mammary gland neoplasia in long-term rodent studies. Environ. Health Perspect. 1996, 104, 938–967. [Google Scholar] [CrossRef] [PubMed]
- Tiede, B.; Kang, Y. From milk to malignancy: The role of mammary stem cells in development, pregnancy and breast cancer. Cell Res. 2011, 21, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.W.; Silberstein, G.B. Postnatal development of the rodent mammary gland. In The Mammary Gland; Neville, M.C., Daniel, C.W., Eds.; Plenum Press: New York, NY, USA, 1987; pp. 3–36. [Google Scholar]
- Asselin-Labat, M.L.; Vaillant, F.; Sheridan, J.M.; Pal, B.; Wu, D.; Simpson, E.R.; Yasuda, H.; Smyth, G.K.; Martin, T.J.; Lindeman, G.J.; et al. Control of mammary stem cell function by steroid hormone signalling. Nature 2010, 465, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Tiede, B.J.; Owens, L.A.; Li, F.; DeCoste, C.; Kang, Y. A novel mouse model for non-invasive single marker tracking of mammary stem cells in vivo reveals stem cell dynamics throughout pregnancy. PLoS ONE 2009, 4, e8035. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.U.; Boulanger, C.A.; Henry, M.D.; Sgagias, M.; Hennighausen, L.; Smith, G.H. An adjunct mammary epithelial cell population in parous females: Its role in functional adaptation and tissue renewal. Development 2002, 129, 1377–1386. [Google Scholar] [PubMed]
- Matulka, L.A.; Triplett, A.A.; Wagner, K.U. Parity-induced mammary epithelial cells are multipotent and express cell surface markers associated with stem cells. Dev. Biol. 2007, 303, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Sogon, T.; Masamura, S.; Hayashi, S.; Santen, R.J.; Nakachi, K.; Eguchi, H. Demethylation of promoter C region of estrogen receptor α gene is correlated with its enhanced expression in estrogen-ablation resistant MCF-7 cells. J. Steroid Biochem. Mol. Biol. 2007, 105, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Block, C.; Freyermuth, S.; Beyersmann, D.; Malviya, A.N. Role of cadmium in activating nuclear protein kinase C and the enzyme binding to nuclear protein. J. Biol. Chem. 1992, 267, 19824–19828. [Google Scholar] [PubMed]
- Stuart, G.W.; Searle, P.F.; Palmiter, R.D. Identification of multiple metal regulatory elements in mouse metallothionein-I promoter by assaying synthetic sequences. Nature 1985, 317, 828–831. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, G.; Regan, J.; Smalley, M.J. Mammary stem cells and breast cancer. Cell. Mol. Life Sci. 2007, 64, 3248–3260. [Google Scholar] [CrossRef] [PubMed]
- Sawaki, M.; Noda, S.; Muroi, T.; Mitoma, H.; Takakura, S.; Sakamoto, S.; Yamasaki, K. Evaluation of an in utero through lactational exposure protocol for detection of estrogenic effects of ethinyl estradiol on the offspring of rats: Preliminary trial. Reprod. Toxicol. 2003, 17, 335–343. [Google Scholar] [CrossRef]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parodi, D.A.; Greenfield, M.; Evans, C.; Chichura, A.; Alpaugh, A.; Williams, J.; Cyrus, K.C.; Martin, M.B. Alteration of Mammary Gland Development and Gene Expression by In Utero Exposure to Cadmium. Int. J. Mol. Sci. 2017, 18, 1939. https://doi.org/10.3390/ijms18091939
Parodi DA, Greenfield M, Evans C, Chichura A, Alpaugh A, Williams J, Cyrus KC, Martin MB. Alteration of Mammary Gland Development and Gene Expression by In Utero Exposure to Cadmium. International Journal of Molecular Sciences. 2017; 18(9):1939. https://doi.org/10.3390/ijms18091939
Chicago/Turabian StyleParodi, Daniela A., Morgan Greenfield, Claire Evans, Anna Chichura, Alexandra Alpaugh, James Williams, Kedra C. Cyrus, and Mary Beth Martin. 2017. "Alteration of Mammary Gland Development and Gene Expression by In Utero Exposure to Cadmium" International Journal of Molecular Sciences 18, no. 9: 1939. https://doi.org/10.3390/ijms18091939
APA StyleParodi, D. A., Greenfield, M., Evans, C., Chichura, A., Alpaugh, A., Williams, J., Cyrus, K. C., & Martin, M. B. (2017). Alteration of Mammary Gland Development and Gene Expression by In Utero Exposure to Cadmium. International Journal of Molecular Sciences, 18(9), 1939. https://doi.org/10.3390/ijms18091939