Time Course of the Phenotype of Blood and Bone Marrow Monocytes and Macrophages in the Lung after Cigarette Smoke Exposure In Vivo

 ,
,  and
and 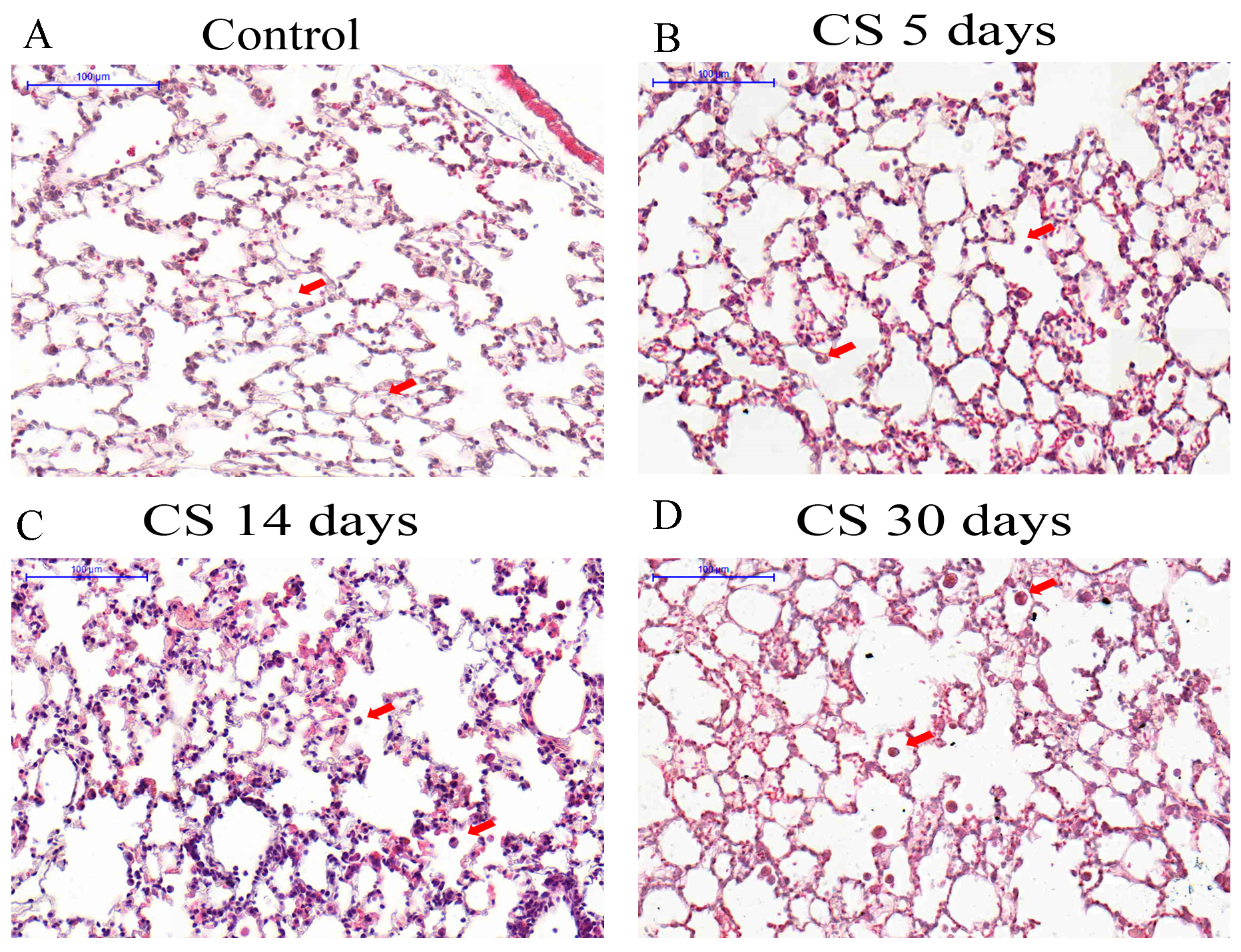{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cigarette Smoke (CS) Exposure Induces Time-Dependent Cellular Inflammation and Destruction in the Airways
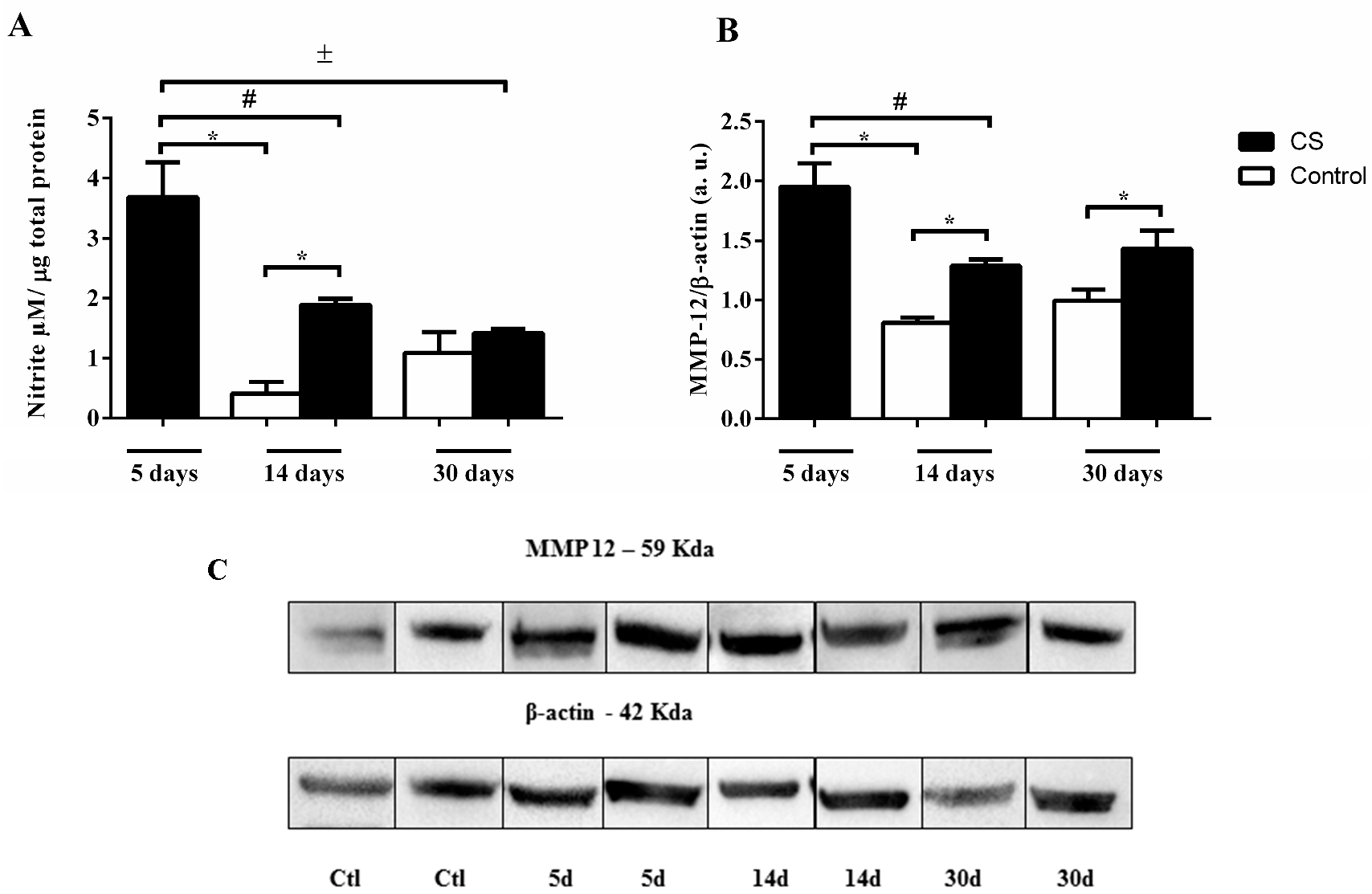
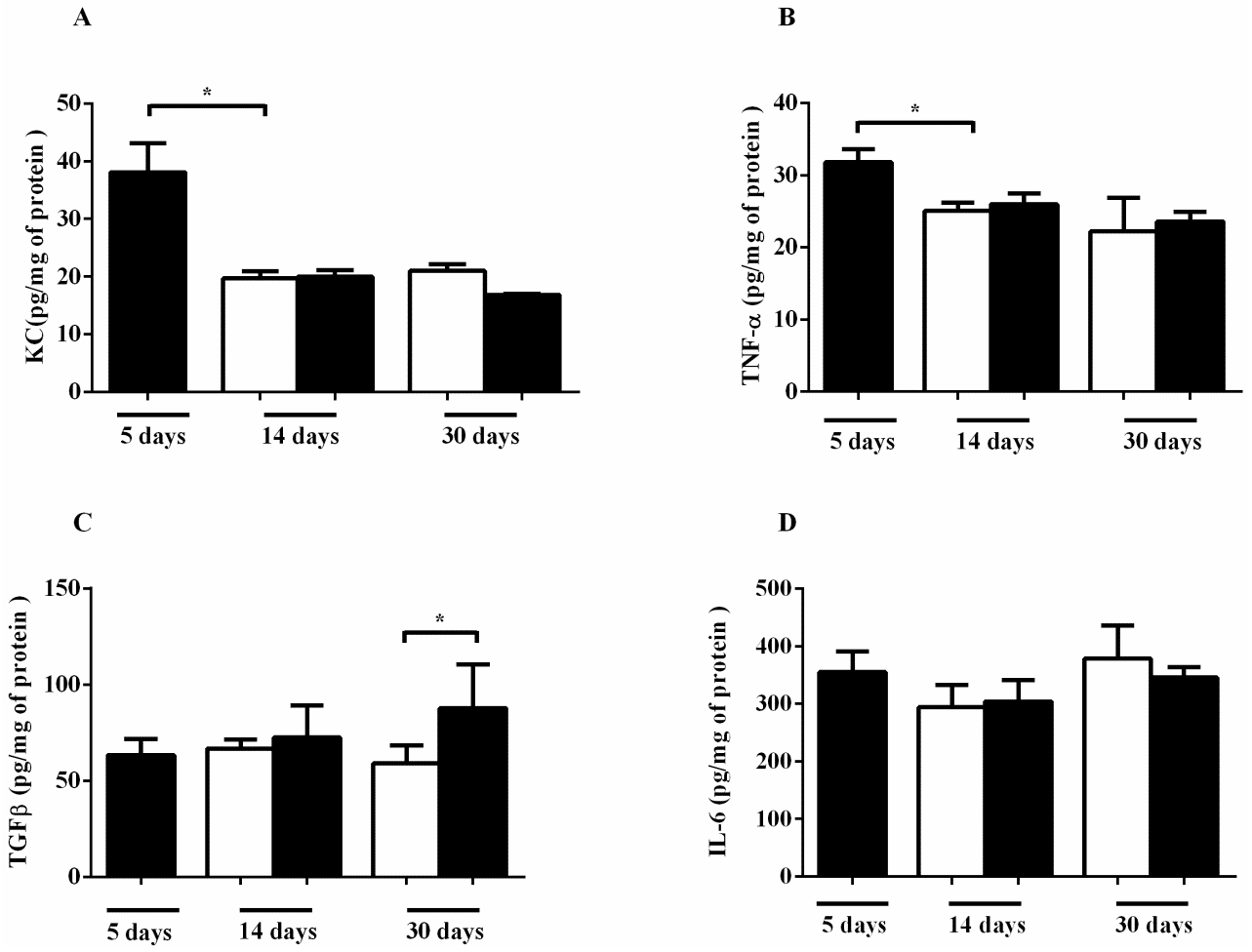
2.2. Effects of CS on Cytokine, NO Release and MMP-12 Expression in Lung Tissue
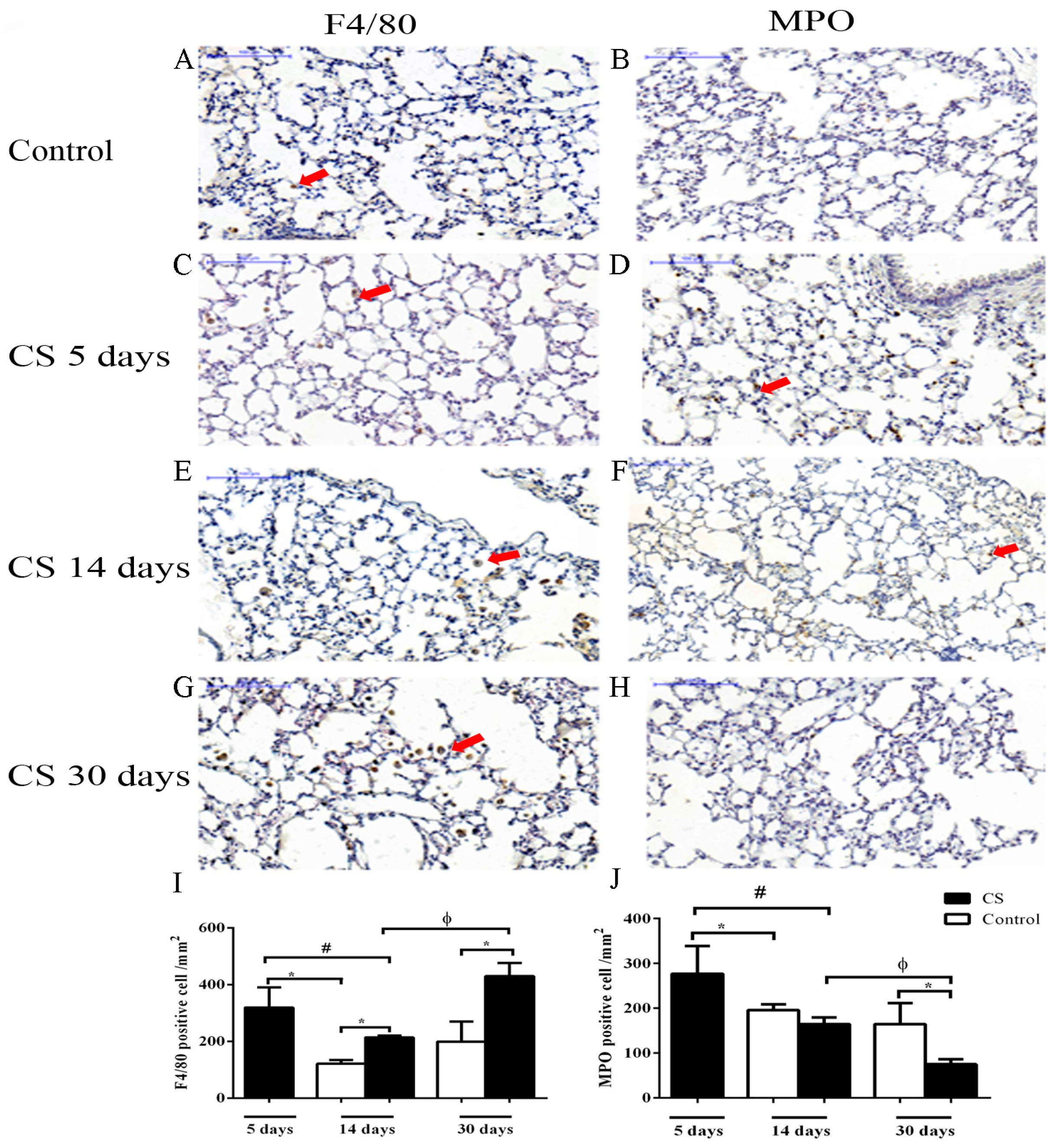
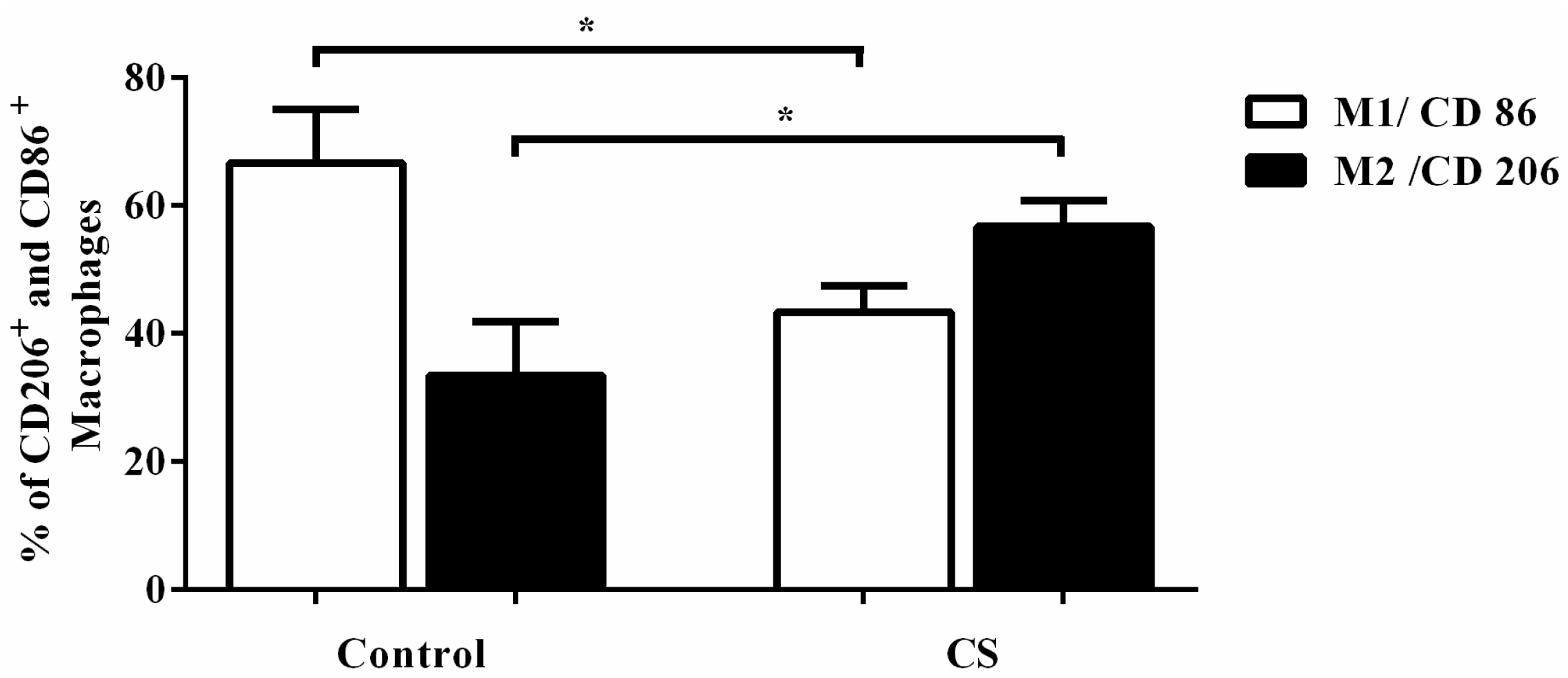
2.3. Effects of CS Macrophage Phenotypes in Bronchoalveolar Lavage (BAL) after Cigarette Smoke Exposure
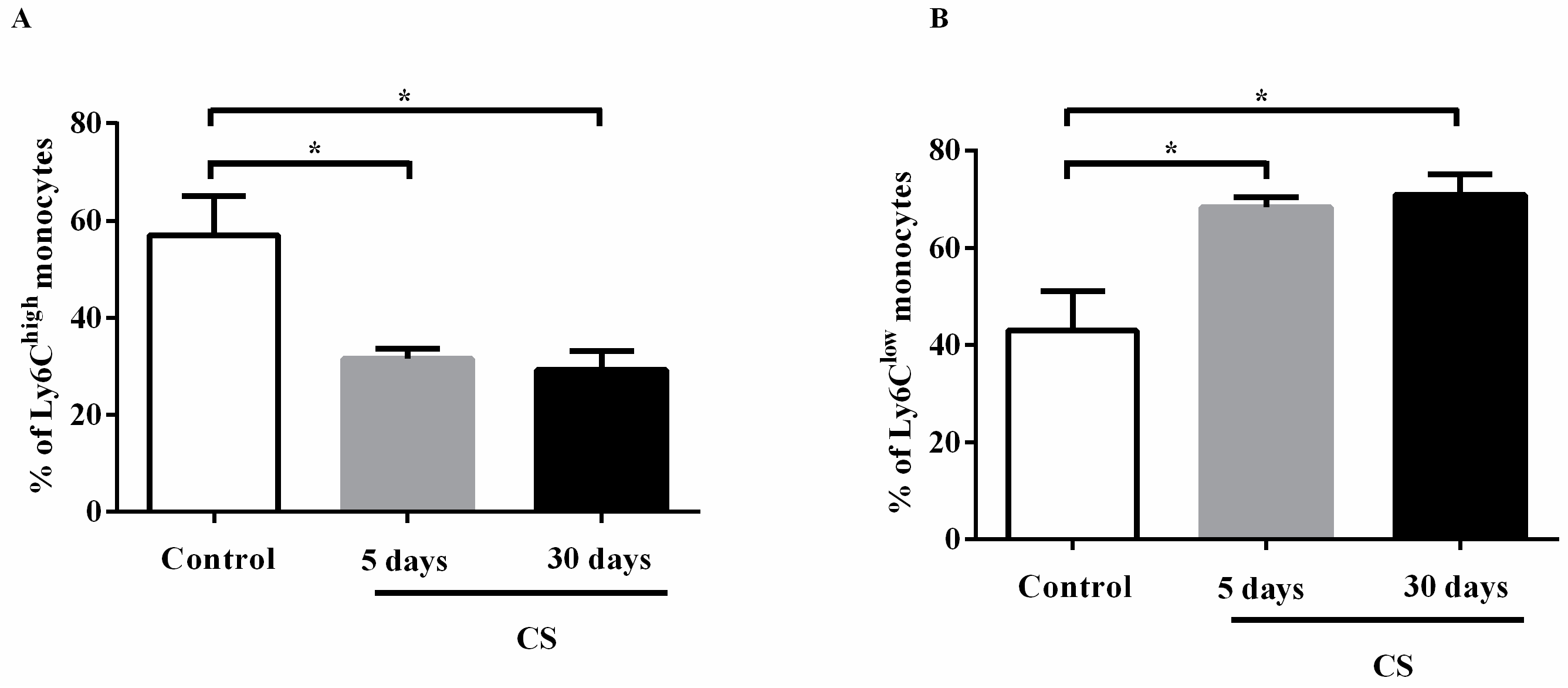
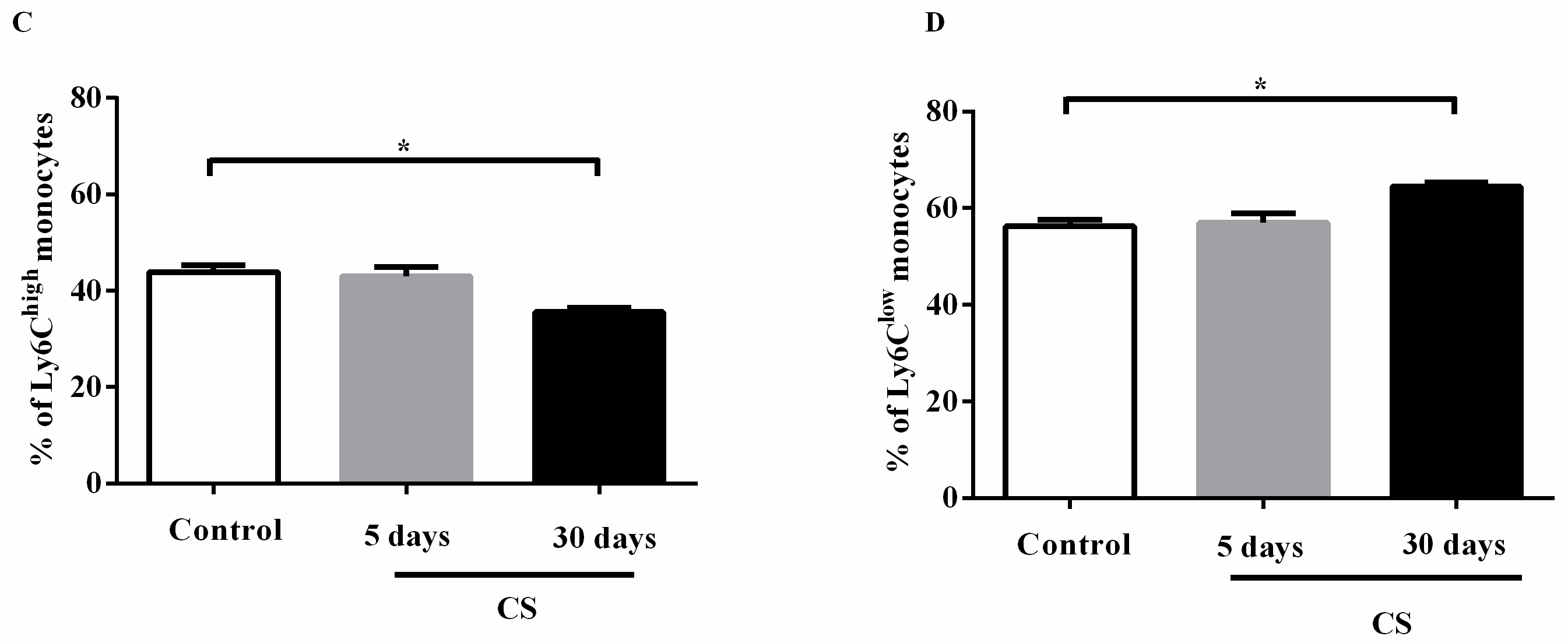
2.4. Effects of the CS Profile of Monocytes in the Circulation and in the Bone Marrow after Cigarette Smoke Exposure
3. Discussion
4. Materials and Methods
4.1. Cigarette Smoke Exposure
4.2. Analysis of Bronchoalveolar Lavage Fluid
4.3. Phenotype Analysis by Fluorescence-Activated Cell Sorting (FACS)
4.4. Tissue Processing
4.5. Histology and Immunohistochemistry
4.6. Analysis of Cytokines, Chemokines and Nitric Oxide
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AM | alveolar macrophages |
| BAL | bronchoalveolar lavage |
| BCA | bicinchoninic acid |
| COPD | chronic obstructive pulmonary disease |
| CS | cigarette smoke |
| IL | interleucin |
| KC | keratinocyte chemoattractant |
| MMP | matrix metalloproteinase |
| MPO | myeloperoxidase |
| NO | nitric oxide |
| ROS | reactive oxygen species |
| TGF-β | transforming growth factor-β |
| TNF-α | tumor necrosis factor-α |
References
- Barnes, P.J. Chronic obstructive pulmonary disease • 12: New treatments for COPD. Thorax 2003, 58, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Rovina, N.; Koutsoukou, A.; Koulouris, N.G. Inflammation and immune response in COPD: Where do we stand? Mediat. Inflamm. 2013. [Google Scholar] [CrossRef] [PubMed]
- Hogg, J.C.; Chu, F.; Utokaparch, S.; Woods, R.; Elliott, W.M.; Buzatu, L.; Cherniack, R.M.; Rogers, R.M.; Sciurba, F.C.; Coxson, H.O.; et al. The nature of small-airway obstruction in chronic obstructive pulmonary disease. N. Engl. J. Med. 2004, 350, 2645–2653. [Google Scholar] [CrossRef] [PubMed]
- Grashoff, W.F.; Sont, J.K.; Sterk, P.J.; Hiemstra, P.S.; de Boer, W.I.; Stolk, J.; Han, J.; van Krieken, J.M. Chronic obstructive pulmonary disease: Role of bronchiolar mast cells and macrophages. Am. J. Pathol. 1997, 151, 1785–1790. [Google Scholar] [PubMed]
- Vlahos, R.; Bozinovski, S. Role of alveolar macrophages in chronic obstructive pulmonary disease. Front. Immunol. 2014, 5, 435. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Keatings, V.M.; Collins, P.D.; Scott, D.M.; Barnes, P.J. Differences in interleukin-8 and tumor necrosis factor-α in induced sputum from patients with chronic obstructive pulmonary disease or asthma. Am. J. Respir. Crit. Care Med. 1996, 153, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Churg, A.; Wang, R.D.; Tai, H.; Wang, X.; Xie, C.; Wright, J.L. Tumor necrosis factor-α drives 70% of cigarette smoke-induced emphysema in the mouse. Am. J. Respir. Crit. Care Med. 2004, 170, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Churg, A.; Dai, J.; Tai, H.; Xie, C.; Wright, J.L. Tumor necrosis factor-α is central to acute cigarette smoke-induced inflammation and connective tissue breakdown. Am. J. Respir. Crit. Care Med. 2002, 166, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Shaykhiev, R.; Krause, A.; Salit, J.; Strulovici-Barel, Y.; Harvey, B.G.; O’Connor, T.P.; Crystal, R.G. Smoking-dependent reprogramming of alveolar macrophage polarization: Implication for pathogenesis of chronic obstructive pulmonary disease. J. Immunol. 2009, 183, 2867–2883. [Google Scholar] [CrossRef] [PubMed]
- Kunz, L.I.; Lapperre, T.S.; Snoeck-Stroband, J.B.; Budulac, S.E.; Timens, W.; van Wijngaarden, S.; Schrumpf, J.A.; Rabe, K.F.; Postma, D.S.; Sterk, P.J.; et al. Smoking status and anti-inflammatory macrophages in bronchoalveolar lavage and induced sputum in COPD. Respir. Res. 2011, 12, 34. [Google Scholar] [CrossRef] [PubMed]
- Kaku, Y.; Imaoka, H.; Morimatsu, Y.; Komohara, Y.; Ohnishi, K.; Oda, H.; Takenaka, S.; Matsuoka, M.; Kawayama, T.; Takeya, M.; et al. Overexpression of CD163, CD204 and CD206 on alveolar macrophages in the lungs of patients with severe chronic obstructive pulmonary disease. PLoS ONE 2014, 9, e87400. [Google Scholar] [CrossRef] [PubMed]
- Bazzan, E.; Turato, G.; Tine, M.; Radu, C.M.; Balestro, E.; Rigobello, C.; Biondini, D.; Schiavon, M.; Lunardi, F.; Baraldo, S.; et al. Dual polarization of human alveolar macrophages progressively increases with smoking and COPD severity. Respir. Res. 2017, 18, 40. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Fu, X.; Shi, H.; Chen, G.; Dong, P.; Zhang, W. Induction of murine macrophage m2 polarization by cigarette smoke extract via the JAK2/STAT3 pathway. PLoS ONE 2014, 9, e107063. [Google Scholar] [CrossRef] [PubMed]
- Maus, U.; Huwe, J.; Ermert, L.; Ermert, M.; Seeger, W.; Lohmeyer, J. Molecular pathways of monocyte emigration into the alveolar air space of intact mice. Am. J. Respir. Crit. Care Med. 2002, 165, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rial, S.; del Puerto-Nevado, L.; Terron-Exposito, R.; Giron-Martinez, A.; Gonzalez-Mangado, N.; Peces-Barba, G. Role of recently migrated monocytes in cigarette smoke-induced lung inflammation in different strain of mice. PLoS ONE 2013, 8, e72975. [Google Scholar] [CrossRef] [PubMed]
- Ziegler-Heitbrock, H.W.; Strobel, M.; Kieper, D.; Fingerle, G.; Schlunck, T.; Petersmann, I.; Ellwart, J.; Blumenstein, M.; Haas, J.G. Differential expression of cytokines in human blood monocyte subpopulations. Blood 1992, 79, 503–511. [Google Scholar] [PubMed]
- Ziegler-Heitbrock, L. The CD14+ CD16+ blood monocytes: Their role in infection and inflammation. J. Leukoc. Biol. 2007, 81, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.; Bueno, C.; Alguero, M.C.; Sanchez, M.L.; de Santiago, M.; Escribano, L.; Diaz-Agustin, B.; Vaquero, J.M.; Laso, F.J.; San Miguel, J.F.; et al. Comparative analysis of the morphological, cytochemical, immunophenotypical, and functional characteristics of normal human peripheral blood lineage−/CD16+/HLA-DR+/CD14−/lo cells, CD14+ monocytes, and CD16− dendritic cells. Clin. Immunol. 2001, 100, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Geissmann, F.; Jung, S.; Littman, D.R. Blood monocytes consist of two principal subsets with distinct migratory properties. Immunity 2003, 19, 71–82. [Google Scholar] [CrossRef]
- Sunderkotter, C.; Nikolic, T.; Dillon, M.J.; Van Rooijen, N.; Stehling, M.; Drevets, D.A.; Leenen, P.J. Subpopulations of mouse blood monocytes differ in maturation stage and inflammatory response. J. Immunol. 2004, 172, 4410–4417. [Google Scholar] [CrossRef] [PubMed]
- Fadini, G.P.; Simoni, F.; Cappellari, R.; Vitturi, N.; Galasso, S.; Vigili de Kreutzenberg, S.; Previato, L.; Avogaro, A. Pro-inflammatory monocyte-macrophage polarization imbalance in human hypercholesterolemia and atherosclerosis. Atherosclerosis 2014, 237, 805–808. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.A.; Ferreira, T.S.; Nesi, R.T.; Lanzetti, M.; Pires, K.M.; Silva, A.M.; Borges, R.M.; Silva, A.J.; Valenca, S.S.; Porto, L.C. Antioxidant action of propolis on mouse lungs exposed to short-term cigarette smoke. Bioorg. Med. Chem. 2013, 21, 7570–7577. [Google Scholar] [CrossRef] [PubMed]
- Doz, E.; Noulin, N.; Boichot, E.; Guenon, I.; Fick, L.; Le Bert, M.; Lagente, V.; Ryffel, B.; Schnyder, B.; Quesniaux, V.F.; et al. Cigarette smoke-induced pulmonary inflammation is TLR4/MyD88 and IL-1R1/MyD88 signaling dependent. J. Immunol. 2008, 180, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- D’Hulst A, I.; Vermaelen, K.Y.; Brusselle, G.G.; Joos, G.F.; Pauwels, R.A. Time course of cigarette smoke-induced pulmonary inflammation in mice. Eur. Respir. J. 2005, 26, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Valenca, S.S.; da Hora, K.; Castro, P.; Moraes, V.G.; Carvalho, L.; Porto, L.C. Emphysema and metalloelastase expression in mouse lung induced by cigarette smoke. Toxicol. Pathol. 2004, 32, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Birrell, M.A.; Wong, S.; Catley, M.C.; Belvisi, M.G. Impact of tobacco-smoke on key signaling pathways in the innate immune response in lung macrophages. J. Cell. Physiol. 2008, 214, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Hogg, J.C.; Wright, J.L.; Wiggs, B.R.; Coxson, H.O.; Opazo Saez, A.; Pare, P.D. Lung structure and function in cigarette smokers. Thorax 1994, 49, 473–478. [Google Scholar] [CrossRef] [PubMed]
- McCrea, K.A.; Ensor, J.E.; Nall, K.; Bleecker, E.R.; Hasday, J.D. Altered cytokine regulation in the lungs of cigarette smokers. Am. J. Respir. Crit. Care Med. 1994, 150, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Le Quement, C.; Guenon, I.; Gillon, J.Y.; Valenca, S.; Cayron-Elizondo, V.; Lagente, V.; Boichot, E. The selective MMP-12 inhibitor, AS111793 reduces airway inflammation in mice exposed to cigarette smoke. Br. J. Pharmacol. 2008, 154, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Nenan, S.; Boichot, E.; Lagente, V.; Bertrand, C.P. Macrophage elastase (MMP-12): A pro-inflammatory mediator? Mem. Inst. Oswaldo Cruz 2005, 100, 167–172. [Google Scholar] [CrossRef]
- Demedts, I.K.; Morel-Montero, A.; Lebecque, S.; Pacheco, Y.; Cataldo, D.; Joos, G.F.; Pauwels, R.A.; Brusselle, G.G. Elevated MMP-12 protein levels in induced sputum from patients with COPD. Thorax 2006, 61, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Morissette, M.C.; Lamontagne, M.; Berube, J.C.; Gaschler, G.; Williams, A.; Yauk, C.; Couture, C.; Laviolette, M.; Hogg, J.C.; Timens, W.; et al. Impact of cigarette smoke on the human and mouse lungs: A gene-expression comparison study. PLoS ONE 2014, 9, e92498. [Google Scholar] [CrossRef] [PubMed]
- Romana-Souza, B.; Nascimento, A.P.; Brum, P.C.; Monte-Alto-Costa, A. Deletion of the α2a/α2c-adrenoceptors accelerates cutaneous wound healing in mice. Inter. J. Exp. Pathol. 2014, 95, 330–341. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira da Silva, C.; Monte-Alto-Costa, A.; Renovato-Martins, M.; Viana Nascimento, F.J.; Dos Santos Valença, S.; Lagente, V.; Pôrto, L.C.; Victoni, T. Time Course of the Phenotype of Blood and Bone Marrow Monocytes and Macrophages in the Lung after Cigarette Smoke Exposure In Vivo. Int. J. Mol. Sci. 2017, 18, 1940. https://doi.org/10.3390/ijms18091940
Oliveira da Silva C, Monte-Alto-Costa A, Renovato-Martins M, Viana Nascimento FJ, Dos Santos Valença S, Lagente V, Pôrto LC, Victoni T. Time Course of the Phenotype of Blood and Bone Marrow Monocytes and Macrophages in the Lung after Cigarette Smoke Exposure In Vivo. International Journal of Molecular Sciences. 2017; 18(9):1940. https://doi.org/10.3390/ijms18091940
Chicago/Turabian StyleOliveira da Silva, Camila, Andréa Monte-Alto-Costa, Mariana Renovato-Martins, Filipe Jorge Viana Nascimento, Samuel Dos Santos Valença, Vincent Lagente, Luís Cristóvão Pôrto, and Tatiana Victoni. 2017. "Time Course of the Phenotype of Blood and Bone Marrow Monocytes and Macrophages in the Lung after Cigarette Smoke Exposure In Vivo" International Journal of Molecular Sciences 18, no. 9: 1940. https://doi.org/10.3390/ijms18091940
APA StyleOliveira da Silva, C., Monte-Alto-Costa, A., Renovato-Martins, M., Viana Nascimento, F. J., Dos Santos Valença, S., Lagente, V., Pôrto, L. C., & Victoni, T. (2017). Time Course of the Phenotype of Blood and Bone Marrow Monocytes and Macrophages in the Lung after Cigarette Smoke Exposure In Vivo. International Journal of Molecular Sciences, 18(9), 1940. https://doi.org/10.3390/ijms18091940