Impaired Skin Barrier Due to Sebaceous Gland Atrophy in the Latent Stage of Radiation-Induced Skin Injury: Application of Non-Invasive Diagnostic Methods

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
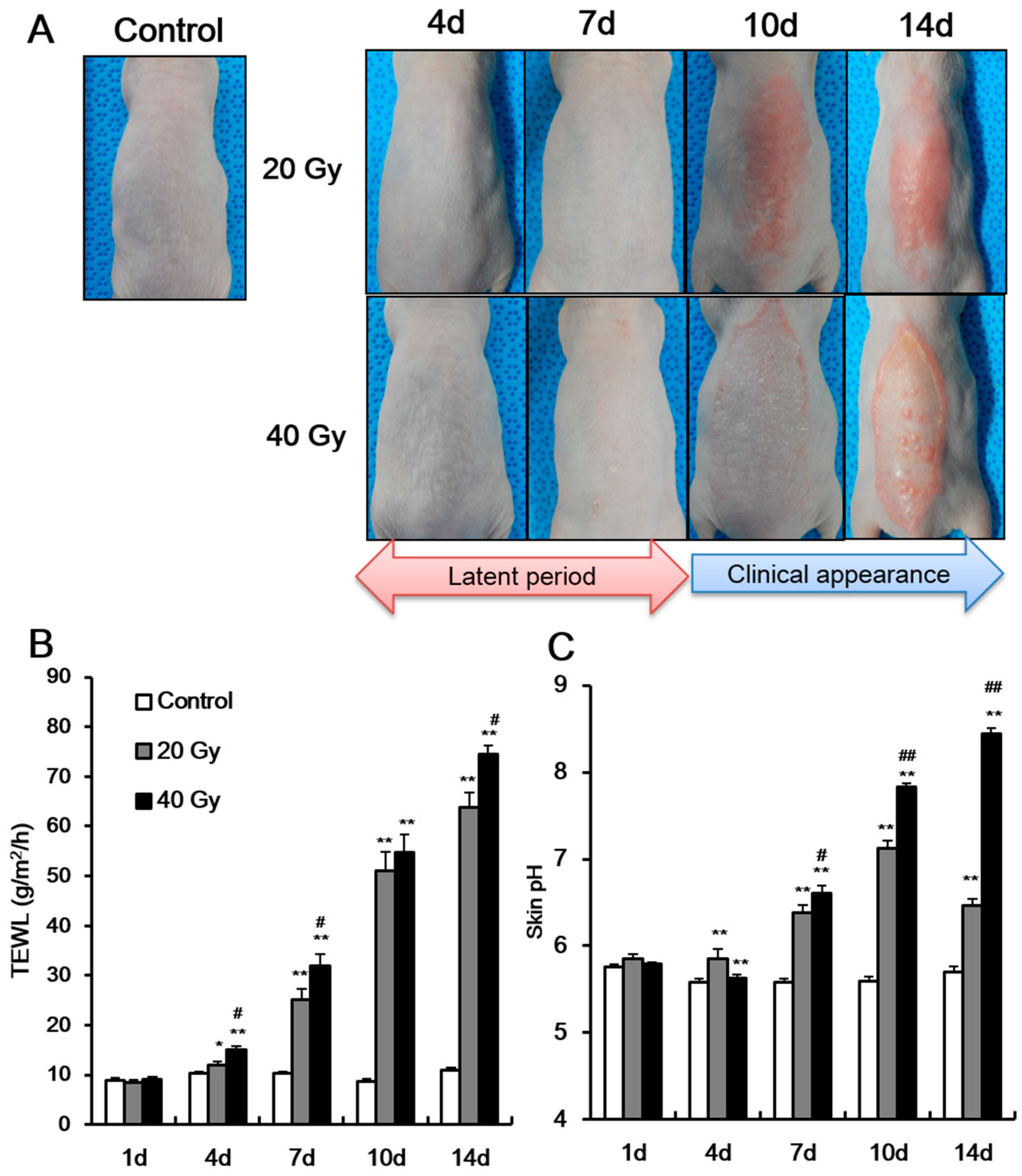
2.1. Radiation Exposure Results in the Development of Skin Barrier Dysfunction in the Latent Stage
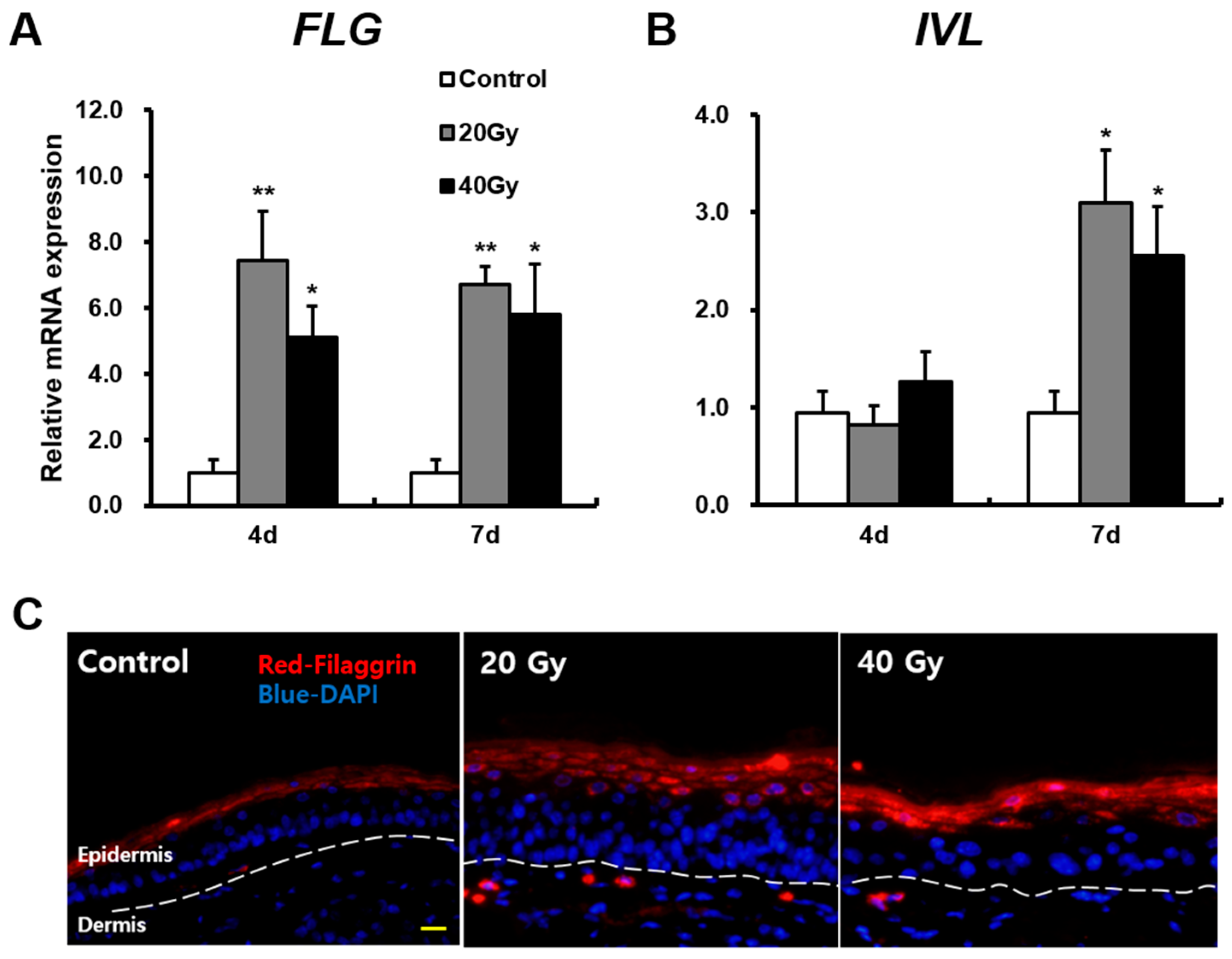
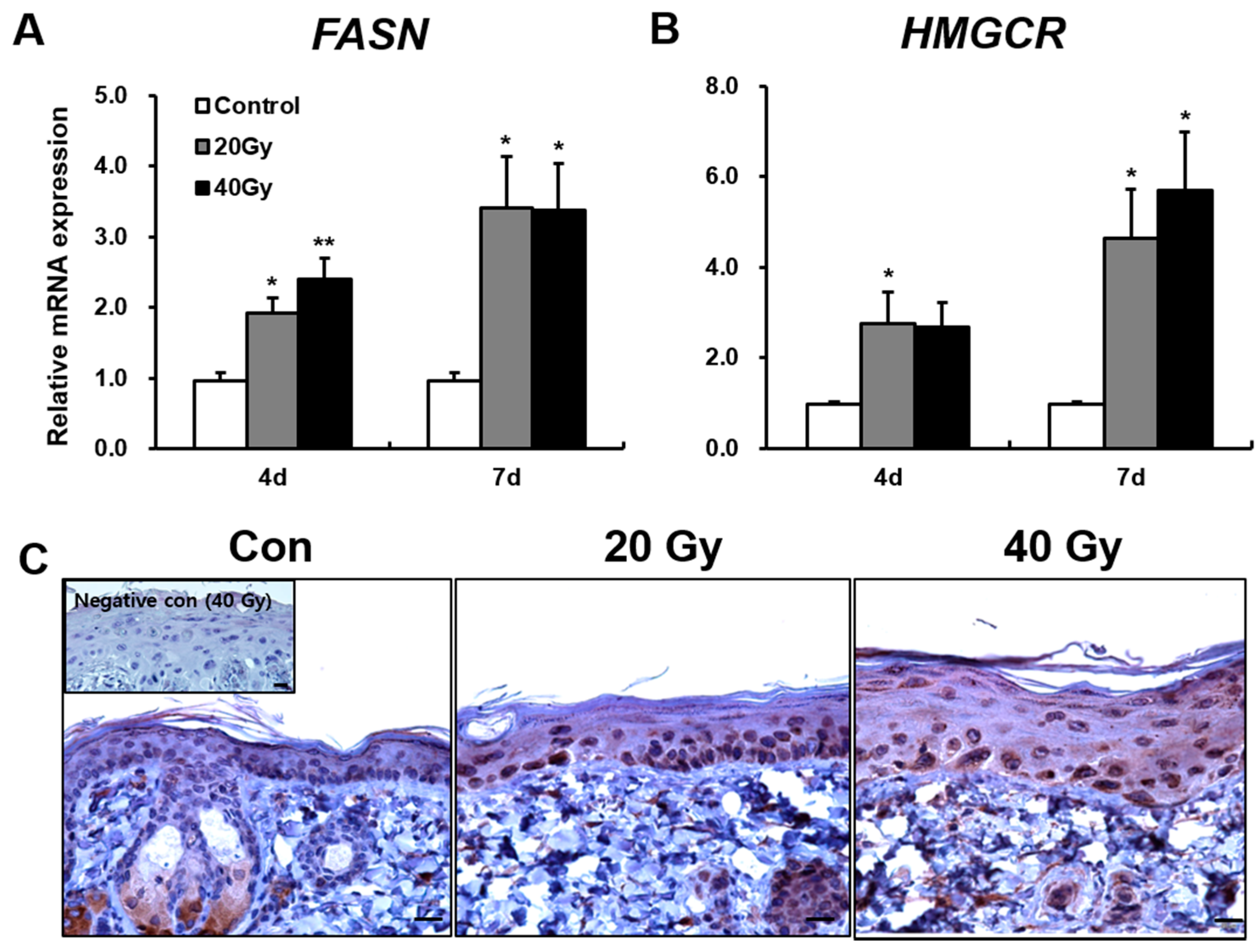
2.2. Skin Barrier Components Are Upregulated in Irradiated Skin in the Early Stage
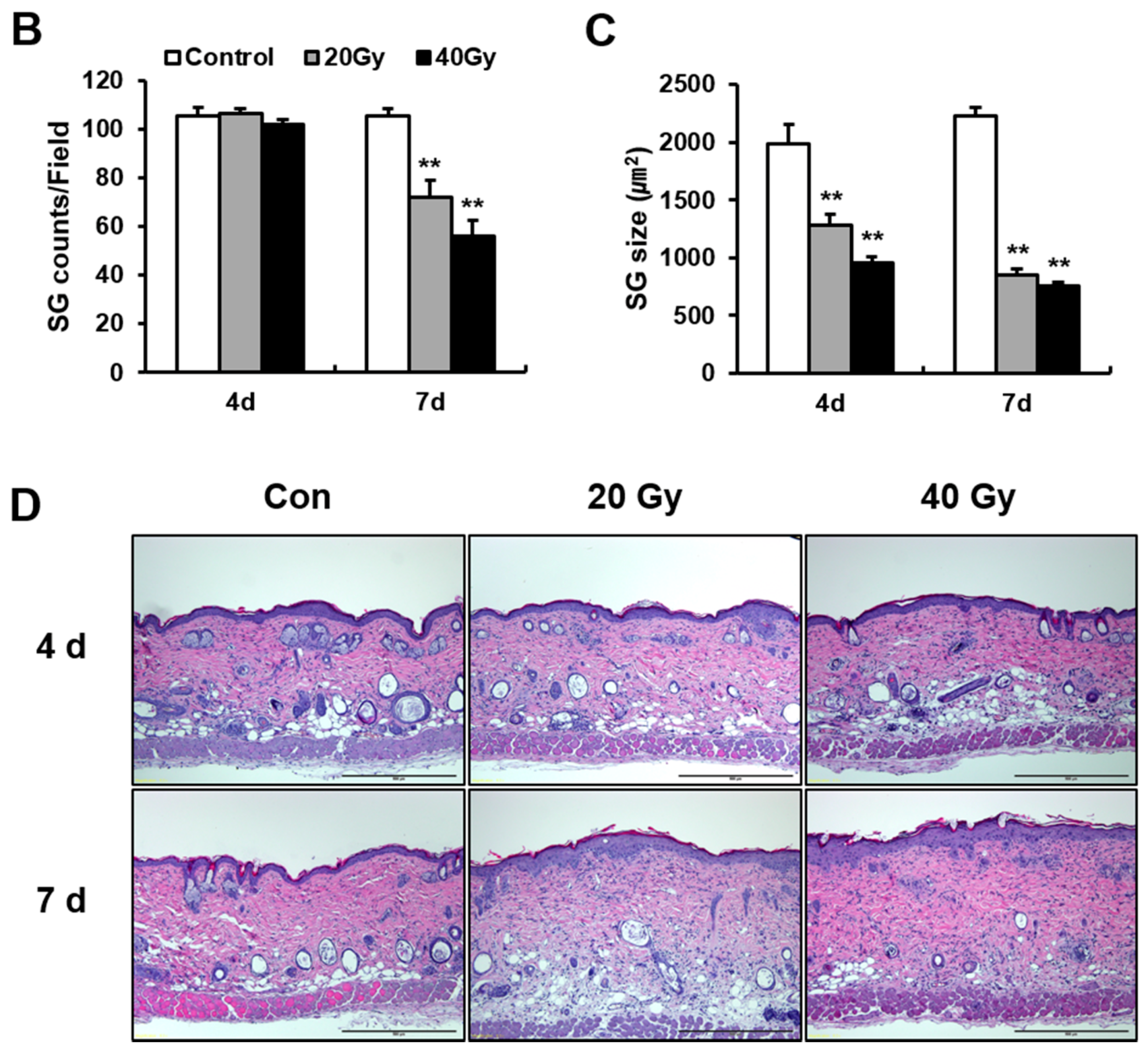
2.3. Early Change in Sebaceous Gland (SG) Impairs the Skin Barrier of Irradiated Skin in the Latent Stage
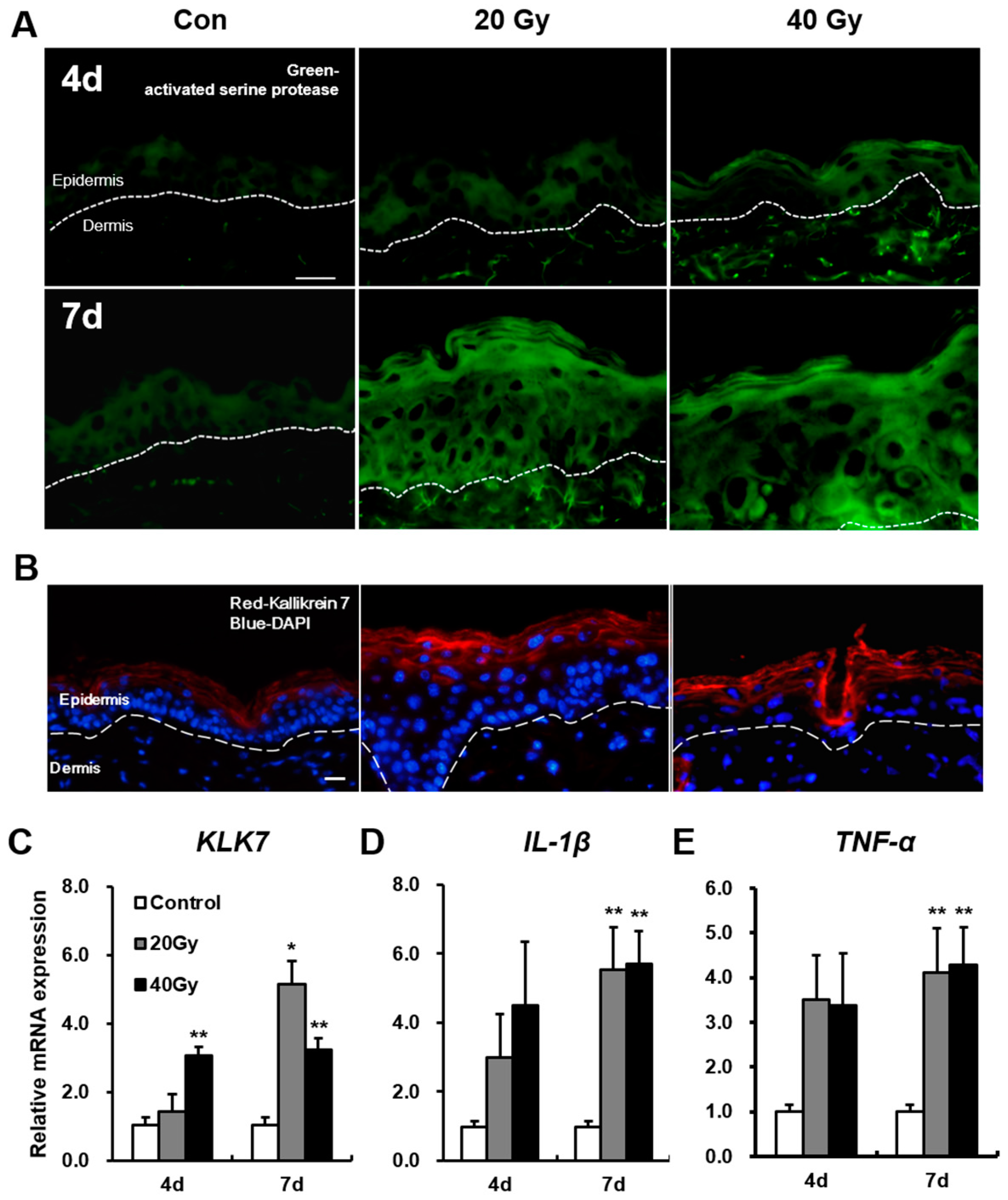
2.4. Serine Protease Hyperactivity Exacerbates Skin Inflammation in Irradiated Skin
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Irradiation of the Skin
4.3. Analysis of Transepidermal Water Loss (TEWL) and Skin Surface pH
4.4. RNA Extraction, Reverse Transcriptase Polymerase Chain Reaction (RT–PCR), and Real-Time PCR Quantification
4.5. Histological Analysis (Immunohistochemistry and Immunofluorescence)
4.6. Epidermal Separation and Sebaceous Gland Analysis
4.7. Serine Protease Activity Assessment by In Situ zymography
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| TEWL | Transepidermal water loss |
| FLG | Filaggrin |
| IVL | Involucrin |
| SG | Sebaceous gland |
| FASN | Fatty acid synthase |
| HMGCR | 3-Hydroxy-3-methyl-glutaryl-coenzyme A reductase |
| KLK | Kallikrein |
| TNF-α | Tumor necrosis factor-α |
| IL-1β | Interleukin-1β |
| RT–PCR | Reverse transcriptase polymerase chain reaction |
| PBS | Phosphate-buffered solution |
References
- Dörr, H.; Meineke, V. Acute radiation syndrome caused by accidental radiation exposure-therapeutic principles. BMC Med. 2011, 25, 126. [Google Scholar] [CrossRef] [PubMed]
- Peter, R.U. Cutaneous radiation syndrome in multi-organ failure. Br. J. Radiol. 2005, 27, 180–184. [Google Scholar] [CrossRef]
- Bray, F.N.; Simmons, B.J.; Wolfson, A.H.; Nouri, K. Acute and Chronic Cutaneous Reactions to Ionizing Radiation Therapy. Dermatol. Ther. 2016, 6, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.L. Ionizing radiation: The good, the bad, and the ugly. J. Investig. Dermatol. 2012, 132, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Hymes, S.R.; Strom, E.A.; Fife, C. Radiation dermatitis: Clinical presentation, patho-physiology, and treatment. J. Am. Acad. Dermatol. 2006, 54, 28–46. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Alavi, A.; Wong, R.; Akita, S. Radiodermatitis: A review of our current understanding. Am. J. Clin. Dermatol. 2016, 17, 277–292. [Google Scholar] [CrossRef] [PubMed]
- Meineken, V. The role of damage to the cutaneous system in radiation–induced multi-organ failure. BRJ Suppl. 2005, 27, 85–99. [Google Scholar]
- Ishii, T.; Futami, S.; Nishida, M.; Suzuki, T.; Sakamoto, T.; Suzuki, N.; Maekawa, K. Brief note and evaluation of acute-radiation syndrome and treatment of a Tokai-mura criticality accident patient. J. Radiat. Res. 2001, 42, S167–S182. [Google Scholar] [CrossRef] [PubMed]
- Andreo, P. Monte Carlo techniques in medical radiation physics. Phys. Med. Biol. 1991, 36, 861–920. [Google Scholar] [CrossRef] [PubMed]
- Roux, A.; Bottollier-Depois, J.F.; Gaillard-Lecanu, E. Qualifying Numerical Tools for Reconstructing Physical Doses in the Case of Accidental Exposure to Ionising Radiation. Radiat. Prot. Dosim. 2000, 87, 243–249. [Google Scholar] [CrossRef]
- Irvine, A.D.; McLean, W.H.; Leung, D.Y. Filaggrin mutations associated with skin and allergic diseases. N. Engl. J. Med. 2011, 365, 1315–1327. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, M.; Dunn-Galvin, A.; Hourihane, J.O.; Murray, D.; Campbell, L.E.; McLean, W.H.; Irvine, A.D. Skin barrier dysfunction measured by transepidermal water loss at 2 days and 2 months predates and predicts atopic dermatitis at 1 year. J. Allergy Clin. Immunol. 2015, 35, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef] [PubMed]
- McAleer, M.A.; Irvine, A.D. The multifunctional role of filaggrin in allergic skin disease. J. Allergy Clin. Immunol. 2013, 131, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Hinde, E.; Haslam, I.S.; Schneider, M.R.; Langan, E.A.; Kloepper, J.E.; Schramm, C.; Zouboulis, C.C.; Paus, R. A practical guide for the study of human and murine sebaceous glands in situ. Exp. Dermatol. 2013, 22, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.R.; Paus, R. Sebocytes, multifaceted epithelial cells: Lipid production and holocrine secretion. Int. J. Biochem. Cell Biol. 2010, 42, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Tóth, B.I.; Oláh, A.; Szöllosi, A.G.; Czifra, G.; Bíró, T. “Sebocytes’ makeup”: Novel mechanisms and concepts in the physiology of the human sebaceous glands. Pflug. Arch. 2011, 461, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Ottey, K.A.; Wood, L.C.; Grunfeld, C.; Elias, P.M.; Feingold, K.R. Cutaneous permeability barrier disruption increases fatty acid synthetic enzyme activity in the epidermis of hairless mice. J. Investig. Dermatol. 1995, 104, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Proksch, E.; Elias, P.M.; Feingold, K.R. Regulation of 3-hydroxy-3-methylglutaryl-coenzyme A reductase activity in murine epidermis. Modulation of enzyme content and activation state by barrier requirements. J. Clin. Investig. 1990, 85, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Brattsand, M.; Egelrud, T. Purification, molecular cloning, and expression of a human stratum corneum trypsin-like serine protease with possible function in desquamation. J. Biol. Chem. 1999, 274, 30033–30040. [Google Scholar] [CrossRef] [PubMed]
- Hansson, L.; Strömqvist, M.; Bäckman, A.; Wallbrandt, P.; Carlstein, A.; Egelrud, T. Cloning, expression, and characterization of stratum corneum chymotryptic enzyme. A skin-specific human serine proteinase. J. Biol. Chem. 1994, 269, 19420–19426. [Google Scholar] [PubMed]
- Hachem, J.P.; Man, M.Q.; Crumrine, D.; Uchida, Y.; Brown, B.E.; Rogiers, V.; Roseeuw, D.; Feingold, K.R.; Elias, P.M. Sustained serine proteases activity by prolonged increase in pH leads to degradation of lipid processing enzymes and profound alterations of barrier function and stratum corneum integrity. J. Investig. Dermatol. 2005, 125, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Takai, T.; Fujimura, T.; Matsuoka, H.; Ogawa, T.; Murayama, K.; Ishii, A.; Ikeda, S.; Okumura, K.; Ogawa, H. Mite serine protease activates protease–activated receptor-2 and induces cytokine release in human keratinocytes. Allergy 2009, 64, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Denda, M.; Kitamura, K.; Elias, P.M.; Feingold, K.R. trans-4-(Aminomethyl)cyclohexane carboxylic acid (T-AMCHA), an anti-fibrinolytic agent, accelerates barrier recovery and prevents the epidermal hyperplasia induced by epidermal injury in hairless mice and humans. J. Investig. Dermatol. 1997, 109, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ding, I.; Chen, K.; Olschowka, J.; Xu, J.; Hu, D.; Morrow, G.R.; Okunieff, P. Interleukin 1β (IL1β) signaling is a critical component of radiation–induced skin fibrosis. Radiat. Res. 2006, 165, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Di Maggio, F.M.; Minafra, L.; Forte, G.I.; Cammarata, F.P.; Lio, D.; Messa, C.; Gilardi, M.C.; Bravatà, V. Portrait of inflammatory response to ionizing radiation treatment. J. Inflamm. 2015, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Xie, G.; Zhu, L.; Chen, X.; Li, X.; Lu, H.; Xu, B.; Ramot, Y.; Paus, R.; Yue, Z. p53 is adirect transcriptional repressor of keratin 17: Lessons from a rat model of radiation dermatitis. J. Investig. Dermatol. 2016, 136, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Moniaga, C.S.; Egawa, G.; Kawasaki, H.; Hara-Chikuma, M.; Honda, T.; Tanizaki, H.; Nakajima, S.; Otsuka, A.; Matsuoka, H.; Kubo, A.; et al. Flaky tail mouse denotes human atopic dermatitis in the steady state and by topical application with Dermatophagoides pteronyssinus extract. Am. J. Pathol. 2010, 176, 2385–2393. [Google Scholar] [CrossRef] [PubMed]
- Törmä, H.; Lindberg, M.; Berne, B. Skin barrier disruption by sodium lauryl sulfate-exposure alters the expressions of involucrin, transglutaminase 1, profilaggrin, and kallikreins during the repair phase in human skin in vivo. J. Investig. Dermatol. 2008, 128, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Matsuda, A.; Jung, K.; Karasawa, K.; Matsuda, K.; Oida, K.; Ishizaka, S.; Ahn, G.; Amagai, Y.; Moon, C.; et al. Skin pH Is the Master Switch of Kallikrein 5-Mediated Skin Barrier Destruction in a Murine Atopic Dermatitis Model. J. Investig. Dermatol. 2016, 136, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R.; Man, M.Q.; Menon, G.K.; Cho, S.S.; Brown, B.E.; Elias, P.M. Cholesterol synthesis is required for cutaneous barrier function in mice. J. Clin. Investig. 1990, 86, 1738–1745. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.H.; Shim, S.; Wang, T.; Yoon, Y.; Jang, W.S.; Myung, J.K.; Park, S.; Kim, K.H. In vivo characterization of early–stage radiation skin injury in a mouse model by two-photon microscopy. Sci. Rep. 2016, 6, 19216. [Google Scholar] [CrossRef] [PubMed]
- Kippenberger, S.; Havlíček, J.; Bernd, A.; Thaçi, D.; Kaufmann, R.; Meissner, M. ‘Nosing Around’ the human skin: What information is concealed in skin odour? Exp. Dermatol. 2012, 21, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Takigawa, H.; Nakagawa, H.; Kuzukawa, M.; Mori, H.; Imokawa, G. Deficient production of hexadecenoic acid in the skin is associated in part with the vulnerability of atopic dermatitis patients to colonization by Staphylococcus aureus. Dermatology 2005, 211, 240–248. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.; Valacchi, G. Surface lipids as multifunctional mediators of skin responses to environmental stimuli. Mediat. Inflamm. 2010, 10, 321494. [Google Scholar] [CrossRef] [PubMed]
- Fluhr, J.W.; Mao-Qiang, M.; Brown, B.E.; Wertz, P.W.; Crumrine, D.; Sundberg, J.P.; Feingold, K.R.; Elias, P.M. Glycerol regulates stratum corneum hydration in sebaceous gland deficient (asebia) mice. J. Investig. Dermatol. 2003, 120, 728–737. [Google Scholar] [CrossRef] [PubMed]
- Brockschnieder, D.; Pechmann, Y.; Sonnenberg-Riethmacher, E.; Riethmacher, D. An improved mouse line for Cre-induced cell ablation due to diphtheria toxin A, expressed from the Rosa26 locus. Genesis 2006, 44, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Firooz, A.; Gorouhi, F.; Davari, P.; Atarod, M.; Hekmat, S.; Rashighi-Firoozabadi, M.; Solhpour, A. Comparison of hydration, sebum and pH values in clinically normal skin of patients with atopic dermatitis and healthy controls. Clin. Exp. Dermatol. 2007, 32, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Briot, A.; Deraison, C.; Lacroix, M.; Bonnart, C.; Robin, A.; Besson, C.; Dubus, P.; Hovnanian, A. Kallikrein 5 induces atopic dermatitis–like lesions through PAR2-mediated thymic stromal lymphopoietin expression in Netherton syndrome. J. Exp. Med. 2009, 206, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Lee, D.W.; Choi, W.H.; Jeon, Y.R.; Kim, S.H.; Cho, H.; Lee, E.J.; Hong, Z.Y.; Lee, W.J.; Cho, J. Development of a porcine skin injury model and characterization of the dose-dependent response to high-dose radiation. J. Radiat. Res. 2013, 54, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Janko, M.; Ontiveros, F.; Fitzgerald, T.J.; Deng, A.; DeCicco, M.; Rock, K.L. IL-1 generated subsequent to radiation-induced tissue injury contributes to the pathogenesis of radiodermatitis. Radiat. Res. 2012, 178, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Braun, K.M.; Niemann, C.; Jensen, U.B.; Sundberg, J.P.; Silva-Vargas, V.; Watt, F.M. Manipulation of stem cell proliferation and lineage commitment: Visualisation of label-retaining cells in wholemounts of mouse epidermis. Development 2003, 130, 5241–5255. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, H.; Myung, H.; Lee, J.; Myung, J.K.; Jang, W.-S.; Lee, S.-J.; Bae, C.-H.; Kim, H.; Park, S.; Shim, S. Impaired Skin Barrier Due to Sebaceous Gland Atrophy in the Latent Stage of Radiation-Induced Skin Injury: Application of Non-Invasive Diagnostic Methods. Int. J. Mol. Sci. 2018, 19, 185. https://doi.org/10.3390/ijms19010185
Jang H, Myung H, Lee J, Myung JK, Jang W-S, Lee S-J, Bae C-H, Kim H, Park S, Shim S. Impaired Skin Barrier Due to Sebaceous Gland Atrophy in the Latent Stage of Radiation-Induced Skin Injury: Application of Non-Invasive Diagnostic Methods. International Journal of Molecular Sciences. 2018; 19(1):185. https://doi.org/10.3390/ijms19010185
Chicago/Turabian StyleJang, Hyosun, Hyunwook Myung, Janet Lee, Jae Kyung Myung, Won-Suk Jang, Sun-Joo Lee, Chang-Hwan Bae, Hyewon Kim, Sunhoo Park, and Sehwan Shim. 2018. "Impaired Skin Barrier Due to Sebaceous Gland Atrophy in the Latent Stage of Radiation-Induced Skin Injury: Application of Non-Invasive Diagnostic Methods" International Journal of Molecular Sciences 19, no. 1: 185. https://doi.org/10.3390/ijms19010185
APA StyleJang, H., Myung, H., Lee, J., Myung, J. K., Jang, W. -S., Lee, S. -J., Bae, C. -H., Kim, H., Park, S., & Shim, S. (2018). Impaired Skin Barrier Due to Sebaceous Gland Atrophy in the Latent Stage of Radiation-Induced Skin Injury: Application of Non-Invasive Diagnostic Methods. International Journal of Molecular Sciences, 19(1), 185. https://doi.org/10.3390/ijms19010185