Maternal Exposure to PM2.5 during Pregnancy Induces Impaired Development of Cerebral Cortex in Mice Offspring

 ,
,
Abstract
:
1. Introduction
2. Results
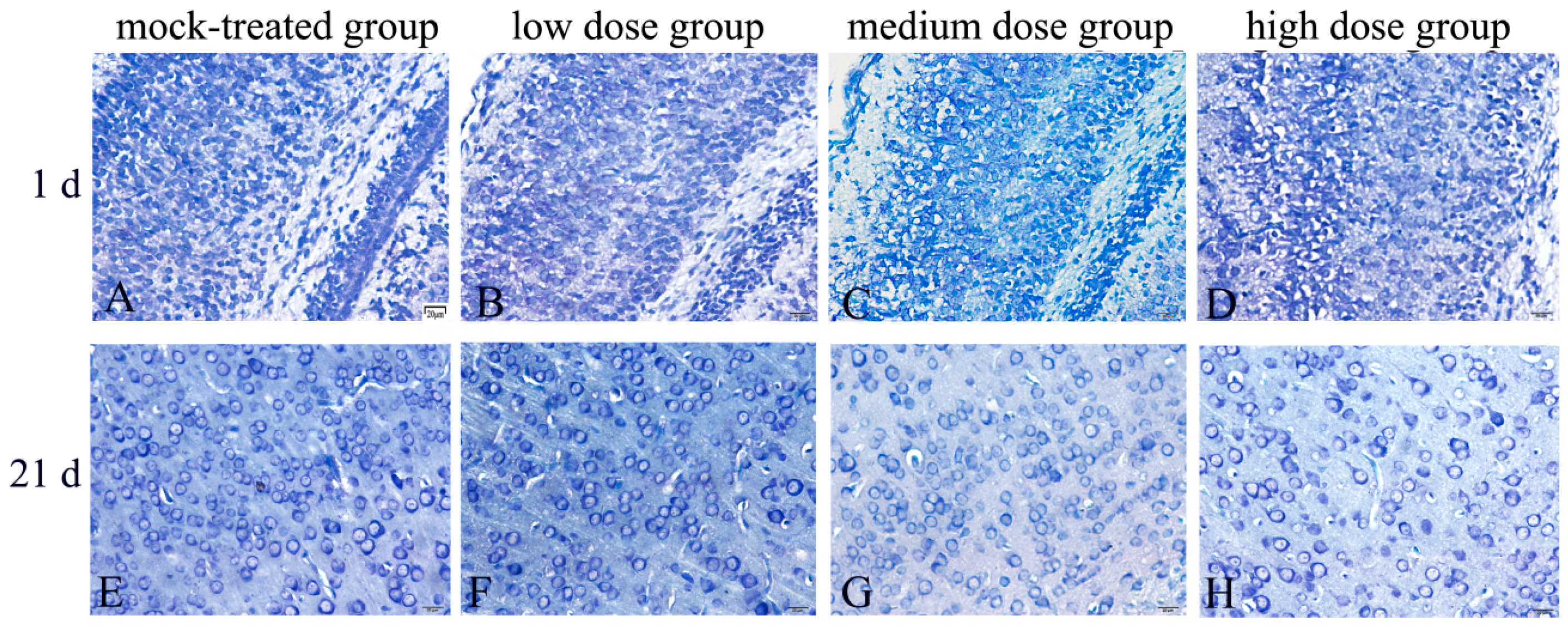
2.1. Exposure to PM2.5 during Pregnancy Caused Pathological Changes of Cerebral Cortex in Mice Offspring
2.2. Ultrastructural Changes of Cerebral Cortical Neurons in Newborn Mice Offspring
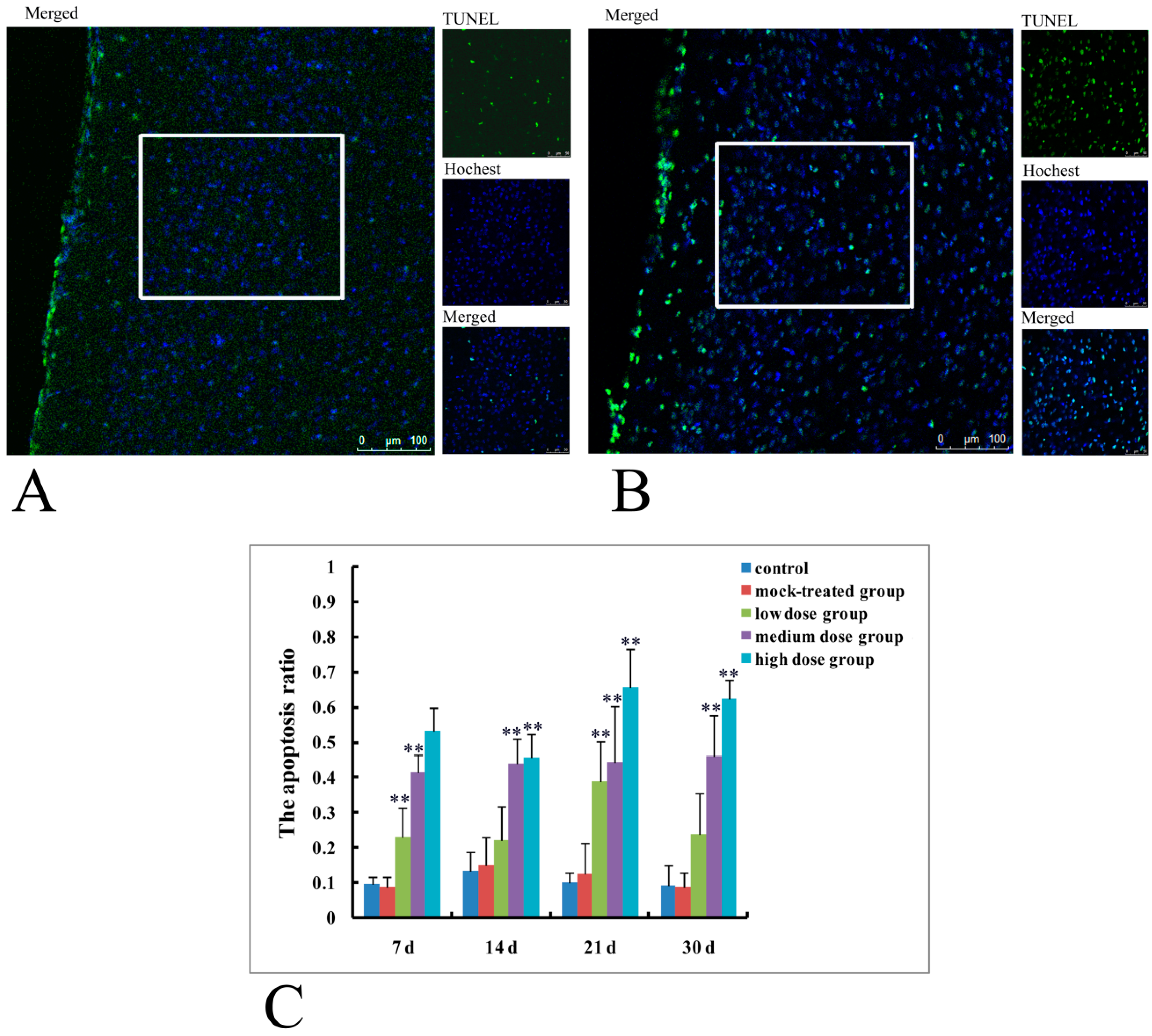
2.3. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL)
2.4. Exposure to PM2.5 during Pregnancy Caused Elevated Expressions of Apoptosis-Related Genes in Cerebral Cortex of Mice Offspring
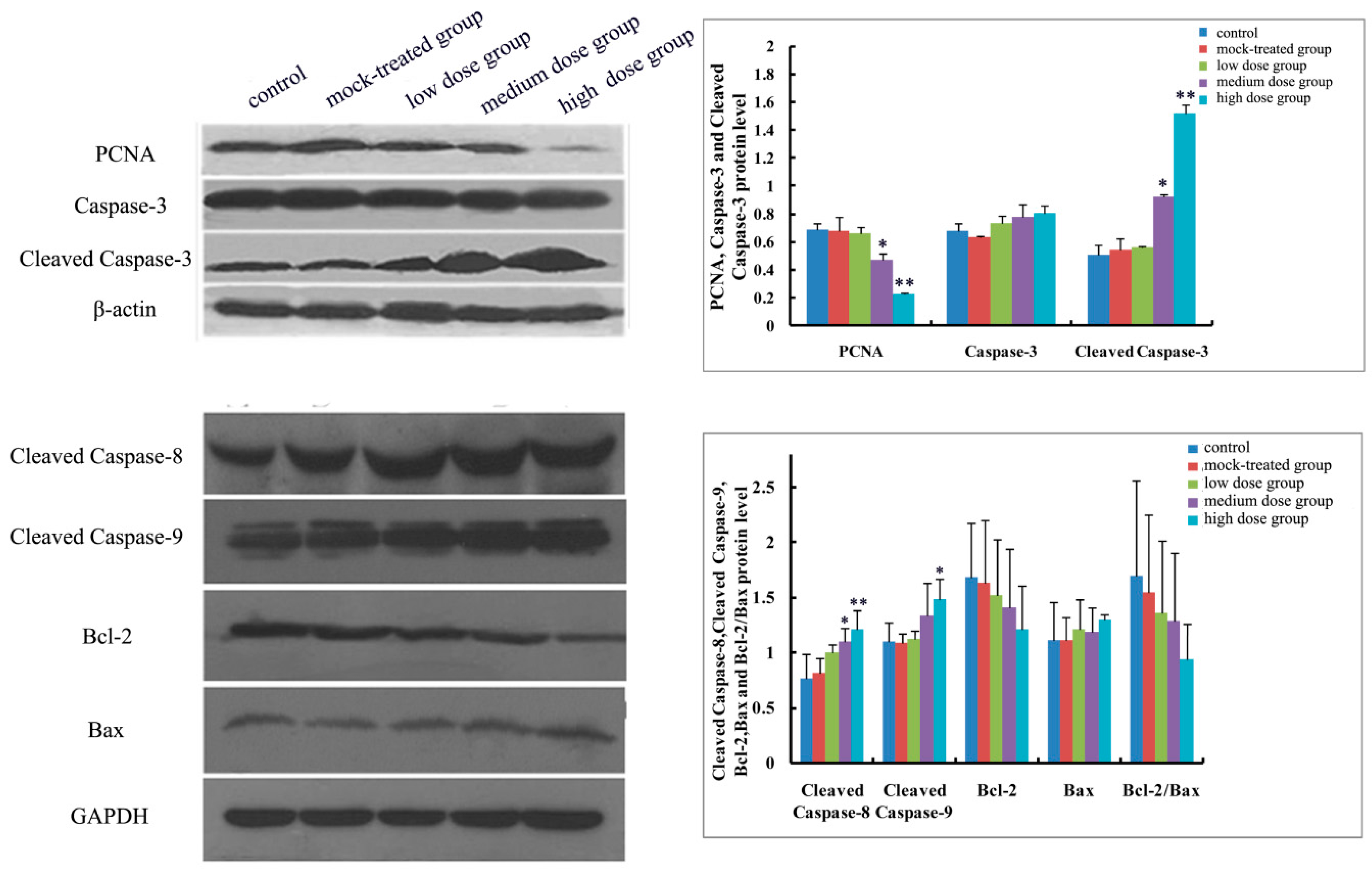
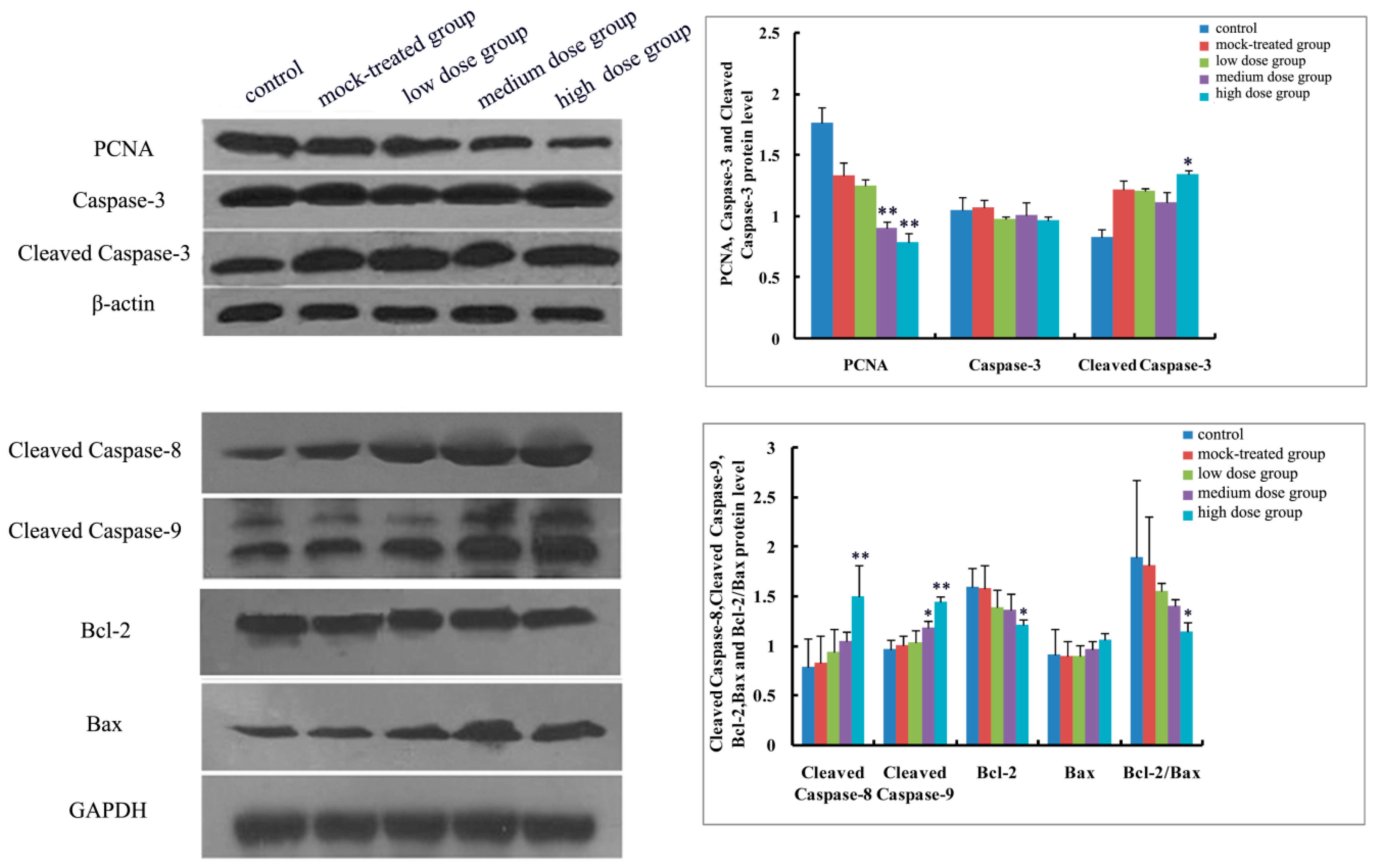
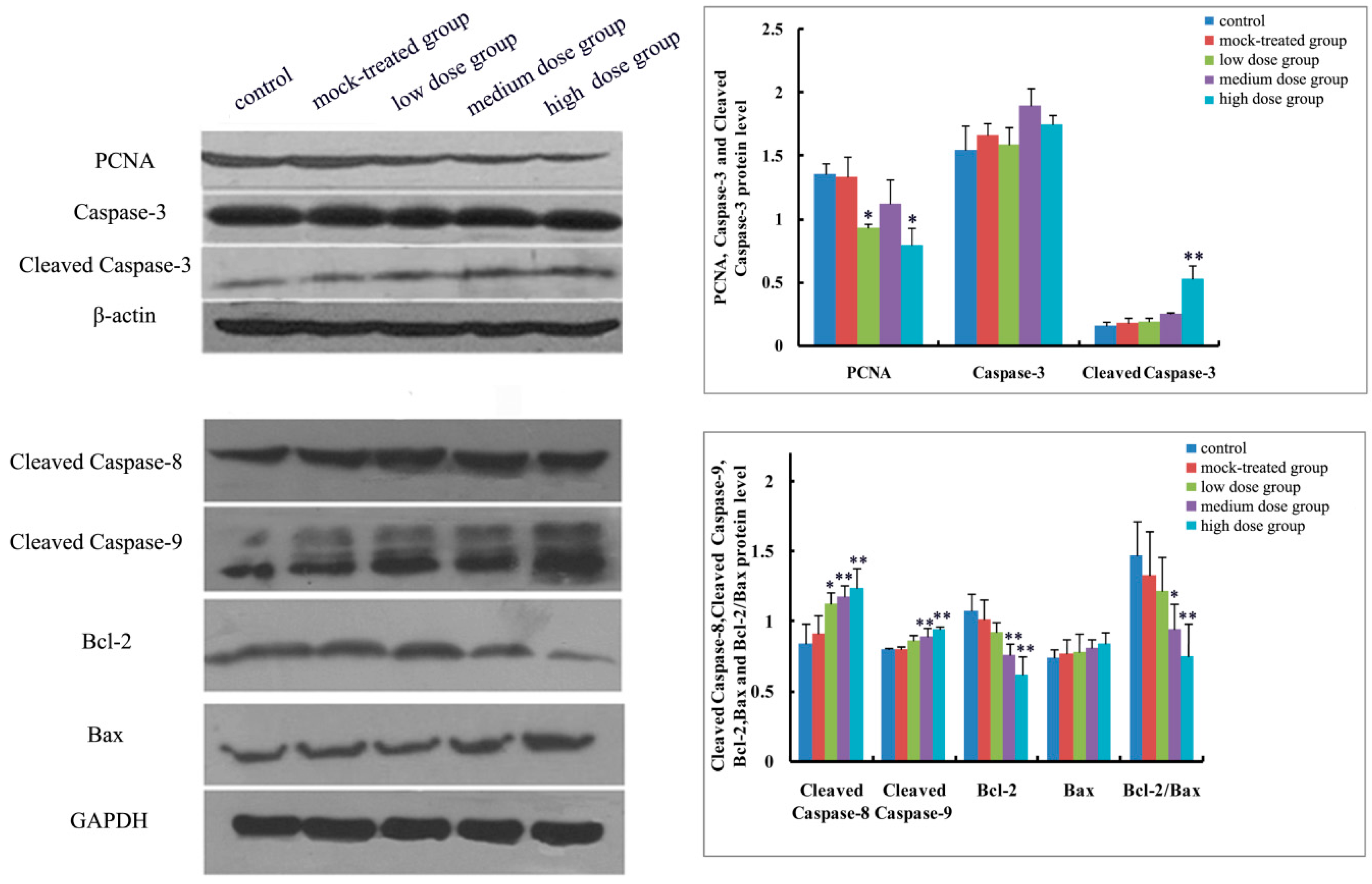
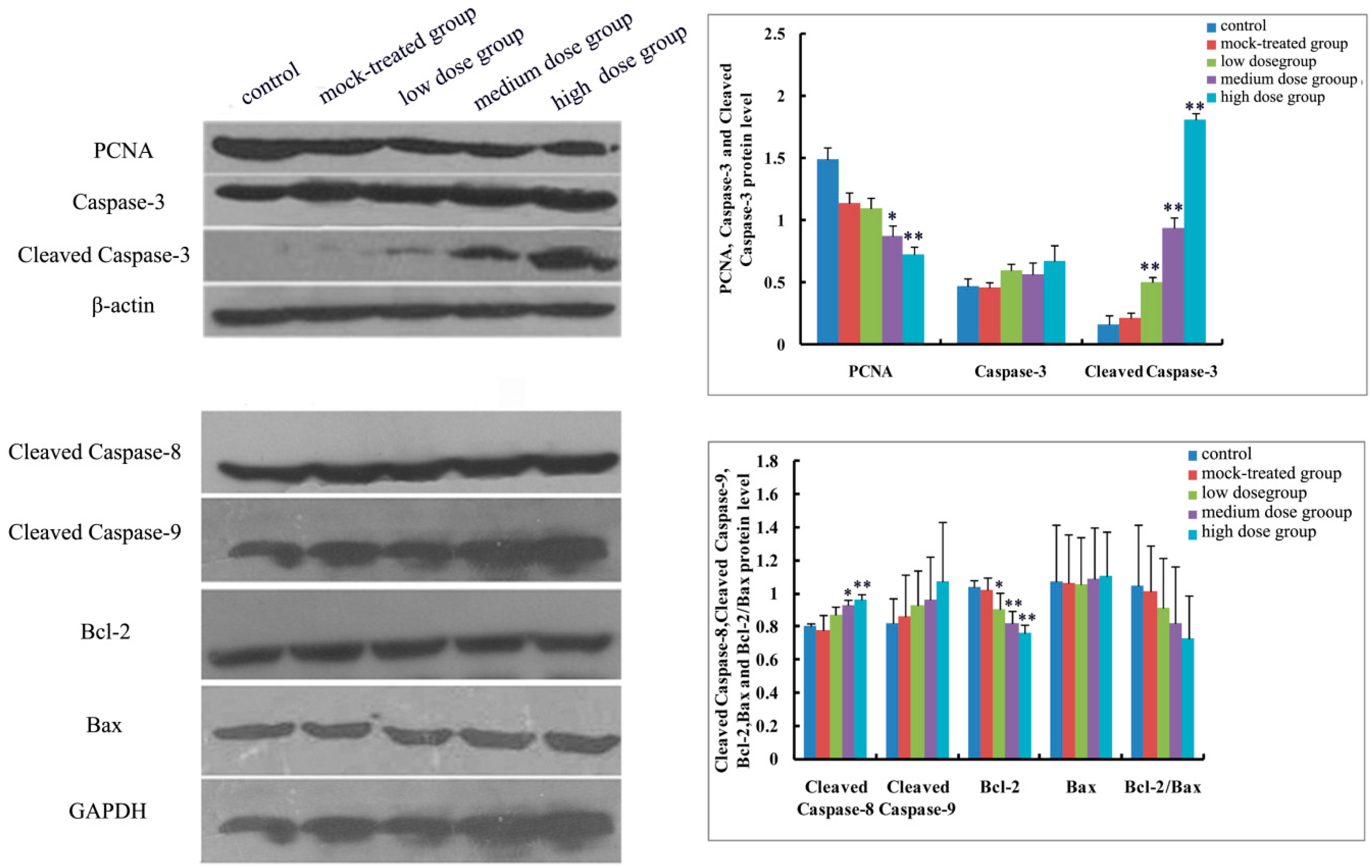
2.4.1. Effects of Exposure to PM2.5 on mRNA Levels of Caspase-3, -8, and -9 and Protein Expressions of Caspase-3, Cleaved Caspase-3, -8, and -9
2.4.2. Effects of Exposure to PM2.5 on mRNA Levels and Protein Expressions of Bcl-2 and Bax
2.4.3. Effects of Exposure to PM2.5 on Protein Expression of PCNA
2.5. Results of Behavioral Experiments in Mice Offspring
2.5.1. Results of Open Field Test
2.5.2. Results of Tail Suspension Experiment
3. Discussion
4. Materials and Methods
4.1. Sampling and Preparation of PM2.5
4.2. Animal Grouping and Model Preparation
4.3. Nissle Staining
4.4. TEM Observation
4.5. TUNEL
4.6. Real-Time Quantitative PCR
4.7. Western Blot
4.8. Behavioral Experiments
4.8.1. Open Field Test
4.8.2. Tail Suspension Experiment
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| EPA | American Environmental Protection Administration |
| TUNEL | Terminal Deoxynucleotidyl transferase dUTP Nick end Labeling |
| PBS | Phosphate Buffered Solution |
| TEM | Transmission Electron Microscope |
References
- Chao, M.W.; Yang, C.H.; Lin, P.T.; Yang, Y.H.; Chuang, Y.C.; Chung, M.C.; Tseng, C.Y. Exposure to PM2.5 causes genetic changes in fetal rat cerebral cortex and hippocampus. Environ. Toxicol. 2017, 32, 1412–1425. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Ma, Y.; Chen, R.; Zhou, Z.; Chen, B.; Kan, H. Size-fractionated particle number concentrations and daily mortality in a Chinese city. Environ. Health Perspect. 2013, 121, 1174–1178. [Google Scholar] [CrossRef] [PubMed]
- De Keijzer, C.; Agis, D.; Ambrós, A.; Arévalo, G.; Baldasano, J.M.; Bande, S.; Barrera-Gómez, J.; Benach, J.; Cirach, M. The association of air pollution and greenness with mortality and life expectancy in Spain: A small-area study. Environ. Int. 2017, 99, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Samoli, E.; Atkinson, R.W.; Analitis, A.; Fuller, G.W.; Beddows, D.; Green, D.C.; Mudway, I.S.; Harrison, R.M.; Anderson, H.R.; Kelly, F.J. Differential health effects of short-term exposure to source-specific particles in London, U.K. Environ. Int. 2016, 97, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Alimohammadi, H.; Fakhri, S.; Derakhshanfar, H.; Hosseini-Zijoud, S.M.; Safari, S.; Hatamabadi, H.R. The effects of air pollution on ischemic stroke admission rate. Chonnam Med. J. 2016, 52, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Scheers, H.; Jacobs, L.; Casas, L.; Nemery, B.; Nawrot, T.S. Long-term exposure to particulate matter air pollution is a risk factor for stroke: Meta-analytical evidence. Stroke 2015, 46, 3058–3066. [Google Scholar] [CrossRef] [PubMed]
- Sancini, G.; Farina, F.; Battaglia, C.; Cifola, I.; Mangano, E.; Mantecca, P.; Camatini, M.; Palestini, P. Health risk assessment for air pollutants: Alterations in lung and cardiac gene expression in mice exposed to Milano winter fine particulate matter (PM2.5). PLoS ONE 2014, 9, e109685. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhen, L.; Lv, P.; Jiang, R.; Song, W. Effects of ozone and fine particulate matter (PM2.5) on rat system inflammation and cardiac function. Wei Sheng Yan Jiu 2013, 42, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Rui, W.; Guan, L.; Zhang, F.; Zhang, W.; Ding, W. PM2.5-induced oxidative stress increases adhesion molecules expression in human endothelial cells through the ERK/AKT/NF-κB-dependent pathway. J. Appl. Toxicol. 2016, 36, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Sunyer, J.; Esnaola, M.; Alvarez-Pedrerol, M.; Forns, J.; Rivas, I.; López-Vicente, M.; Suades-González, E.; Foraster, M.; Garcia-Esteban, R.; Basagaña, X. Association between traffic-related air pollution in schools and cognitive development in primary school children: A prospective cohort study. PLoS Med. 2015, 12, E1001792. [Google Scholar] [CrossRef] [PubMed]
- Genc, S.; Zadeoglulari, Z.; Fuss, S.H.; Genc, K. The adverse effects of air pollution on the nervous system. J. Toxicol. 2012, 2012, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Garcidueñas, L.; Reed, W.; Maronpot, R.R.; Henríquez-Roldán, C.; Delgado-Chavez, R.; Calderón-Garcidueñas, A.; Dragustinovis, I.; Franco-Lira, M.; Aragón-Flores, M.; Solt, A.C. Brain inflammation and Alzheimer’s-like pathology in individuals exposed to severe air pollution. Toxicol. Pathol. 2004, 32, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Jörg, S.; Grohme, D.A.; Erzler, M.; Binsfeld, M.; Haghikia, A.; Müller, D.N.; Linker, R.A.; Kleinewietfeld, M. Environmental factors in autoimmune diseases and their role in multiple sclerosis. Cell. Mol. Life Sci. 2016, 73, 4611–4622. [Google Scholar] [CrossRef] [PubMed]
- Onoda, A.; Takeda, K.; Umezawa, M. Dose-dependent induction of astrocyte activation and reactive astrogliosis in mouse brain following maternal exposure to carbon black nanoparticle. Particle Fibre Toxicol. 2017, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Lelieveld, J.; Evans, J.S.; Fnais, M.; Giannadaki, D.; Pozzer, A. The contribution of outdoorair pollution sources to premature mortality on a global scale. Nature 2015, 525, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Pujol, J.; Martínez-Vilavella, G.; Macià, D.; Fenoll, R.; Alvarez-Pedrerol, M.; Rivas, I.; Forns, J.; Blanco-Hinojo, L.; Capellades, J.; Querol, X. Traffic pollution exposure is associated with altered brain connectivity inschool children. Neuroimage 2016, 129, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.L.; Liu, X.; Weston, D.; Prince, L.; Oberdörster, G.; Finkelstein, J.N.; Johnston, C.J.; Cory-Slechta, D.A. Developmental exposure to concentrated ambient ultrafine particulate matter air pollution in mice results in persistent and sex-dependent behavioral neurotoxicity and glial activation. Toxicol. Sci. 2014, 140, 160–178. [Google Scholar] [CrossRef] [PubMed]
- Woodward, N.; Finch, C.E.; Morgan, T.E. Traffic-related air pollution and brain development. AIMS Environ. Sci. 2015, 2, 353–373. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.L.; Oberdorster, G.; Morris-Schaffer, K.; Wong, C.; Klocke, C.; Sobolewski, M.; Conrad, K.; Mayer-Proschel, M.; Cory-Slechta, D.A. Developmental neurotoxicity of inhaled ambient ultrafine particle air pollution: Parallels with neuropathological and behavioral features of autism and other neurodevelopmental disorders. Neurotoxicology 2017, 59, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Chang, Y.C.; Cole, T.B. Developmental neurotoxicity of traffic-related air pollution: Focus on autism. Curr. Environ. Health Rep. 2017, 4, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Tan, J.F.; Liu, D. Exposure to Atmospheric fine particulate matter damages multiple organs of pregnant mice and inhibits pregnancy and fetal development during pregnancy. Chin. J. Histochem. Cytochem. 2016, 25, 42–48. [Google Scholar]
- Zheng, X.R.; Zhao, H.; Tian, H.B. PM2.5 exposure during pregnancy causes abnormal development of major organs in offspring mice. Chin. J. Histochem. Cytochem. 2017, 26, 366–372. [Google Scholar]
- Brauer, M.; Lencar, C.; Tamburic, L.; Koehoorn, M.; Demers, P.; Karr, C. A cohort study of traffic-related air pollution impacts on birth outcomes. Environ. Health Perspect. 2008, 116, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, X.; Hong, X.; Liu, C.; Huang, H.; Wang, Q.; Chen, S.; Chen, H.; Yang, K.; Sun, Q. Maternal Exposure to ambient PM2.5 exaggerates fetal cardiovascular maldevelopment induced by homocysteine in rats. Environ. Toxicol. 2017, 32, 877–889. [Google Scholar] [CrossRef] [PubMed]
- Sagai, M.; Tin Win-Shwe, T. Oxidative stress derived from airborne fine and ultrafine particles and the effects on brain-nervous system: Part 2. Nihon Eiseigaku Zasshi 2015, 70, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.H.; Gold, D.R.; Rifas-Shiman, S.L.; Melly, S.J.; Zanobetti, A.; Coull, B.A.; Schwartz, J.D.; Gryparis, A.; Kloog, I.; Koutrakis, P. Prenatal and childhood traffic-related air pollution exposure and childhood executive function and behavior. Neurotoxicol. Teratol. 2016, 57, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Danysh, H.E.; Zhang, K.; Mitchell, L.E.; Scheurer, M.E.; Lupo, P.J. Maternal residential proximity to major roadways at delivery and childhood central nervous system tumors. Environ. Res. 2016, 146, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Raz, R.; Roberts, A.L.; Lyall, K.; Hart, J.E.; Just, A.C.; Laden, F.; Weisskopf, M.G. Autism spectrum disorder and particulate matter air pollution before, during, and after pregnancy: A nested case-control analysis within the Nurses’ Health Study II Cohort. Environ. Health Perspect. 2015, 123, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Ku, T.; Ji, X.; Zhang, Y.; Li, G.; Sang, N. PM2.5, SO2 and NO2 co-exposure impairs neurobehavior and induces mitochondrial injuries in the mouse brain. Chemosphere 2016, 163, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Block, M.L.; Calderón-Garcidueñas, L. Air pollution: Mechanisms of neuroinflammationand CNS disease. Trends Neurosci. 2009, 32, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Kanao, R.; Masutani, C. Regulation of DNA damage tolerance in mammalian cells by posttranslational modifications of PCNA. Mutat. Res. 2017, 803–805, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bai, C.; Guan, H.; Chen, R.; Wang, X.; Wang, B.; Jin, H.; Piao, F. Subchronic exposure to arsenic induces apoptosis in the hippocampus of the mouse brains through the Bcl-2/Bax pathway. J. Occup. Health 2015, 57, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Guru, S.K.; Rath, S.K.; Chinthakindi, P.K.; Singh, B.; Koul, S.; Bhushan, S.; Sangwan, P.L. A novel triazole derivative of betulinic acid induces extrinsic and intrinsic apoptosis in human leukemia HL-60 cells. Eur. J. Med. Chem. 2016, 108, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, J.; Gong, Z. Resveratrol induces apoptosis in K562 cells via the regulation of mitochondrial signaling pathways. Int. J. Clin. Exp. Med. 2015, 8, 16926–16933. [Google Scholar] [PubMed]
- Dong, J.; Zhao, J.; Lin, Y.; Liang, H.; He, X.; Zheng, X.; Sui, M.; Zhuang, Z.; Yan, T. Exercise improves recognition memory and synaptic plasticity in the prefrontal cortex for rats modelling vascular dementia. Neurol. Res. 2017, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Cai, P.; Zhou, J.; Liu, M.; Jiang, X. Effects of asiaticoside on human umbilical vein endothelial cell apoptosis induced by Aβ1-42. Int. J. Clin. Exp. Med. 2015, 8, 15828–15833. [Google Scholar] [PubMed]
- Khwanraj, K.; Phruksaniyom, C.; Madlah, S.; Dharmasaroja, P. Differential Expression of Tyrosine Hydroxylase Protein and Apoptosis-Related Genes in Differentiated and Undifferentiated SH-SY5Y Neuroblastoma Cells Treated with MPP+. Neurol. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Kim, J.E.; Park, W.H. Cytoprotective effect of rhamnetin on miconazole-induced H9c2 cell damage. Nutr. Res. Pract. 2015, 9, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tu, M.; Cheng, C.; Tian, J.; Zhang, F.; Deng, Z.; Li, X.; Li, Z.; Liu, Y.; Lei, G. Wogonoside induces apoptosis in Bel-7402, a hepatocellular carcinoma cell line, by regulating Bax/Bcl-2. Oncol. Lett. 2015, 10, 1831–1835. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.L.; Jiang, L.; Qu, F.; Wang, Z.Y.; Zhao, L.M. Inhibitory effect of Embelin on human acute T cell lymphoma Jurkat cells through activation of the apoptotic pathway. Oncol. Lett. 2015, 10, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Mu, L. The role of heme oxygenase-1 (HO-1) in the regulation of inflammatory reaction, neuronal cell proliferation and apoptosis in rats after intracerebral hemorrhage (ICH). Neuropsychiatr. Dis. Treat. 2016, 13, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.T.; Youle, R.J. Nonionic detergents induce dimerization among members of the Bcl-2 family. J. Biol. Chem. 1997, 272, 13829–13834. [Google Scholar] [CrossRef]
- Zimcikova, E.; Simko, J.; Karesova, I.; Kremlacek, J.; Malakova, J. Behavioral effects of antiepileptic drugs in rats: Are the effects on mood and behavior detectable in open-field test? Seizure 2017, 52, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Bae, C.H.; Jun, Y.L.; Jeon, H.; Koo, S.; Kim, S. Acupuncture alters pro-inflammatory cytokines in the plasma of maternally separated rat pups. Chin. J. Integr. Med. 2017, 23, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, J.; Lin, Q.; Mao, K.; Tian, F.; Jing, C.; Wang, C.; Ding, L.; Pang, C. Proanthocyanidin prevents lipopolysaccharide-induced depressive-like behavior in mice via neuroinflammatory pathway. Brain Res. Bull. 2017, 135, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Miner, L.H.; Schroeter, S.; Blakely, R.D.; Sesack, S.R. Ultrastructural localization of the serotonin transporter in superficial and deep layers of the rat prelimbic prefrontal cortex and its spatial relationship to dopamine terminals. J. Comp. Neurol. 2000, 427, 220–234. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Diameter of Neurons (μm) | Number of Neurons |
|---|---|---|
| Mock-treated group | 18.25 ± 1.09 | 59 ± 4.58 |
| Low-dosage group | 17.66 ± 0.88 | 58 ± 5.29 |
| Medium-dosage group | 14.58 ± 1.02 ** | 45.33 ± 6.43 * |
| High-dosage group | 14.34 ± 1.14 ** | 38 ± 7.21 ** |
| Target | Primer Sequence (5′–3′) |
|---|---|
| Caspase-3 | F: CTGGACTGCGGTATTGAGAC |
| R: CCGGGTGCGGTAGAGTAAGC | |
| Caspase-8 | F: TGCTTGGACTACATCCCACAC |
| R: TGCAGTCTAGGAAGTTGACCA | |
| Caspase-9 | F: TCCTGGTACATCGAGACCTTG |
| R: AAGTCCCTTTCGCAGAAACAG | |
| Bcl-2 | F: GTCGCTACCGTCGTGACTTC |
| R: CAGACATGCACCTACCCAGC | |
| Bax | F: TGAAGACAGGGGCCTTTTTG |
| R: AATTCGCCGGAGACACTCG | |
| β-actin | F: GGCTGTATTCCCCTCCATCG |
| R: CCAGTTGGTAACAATGCCATGT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Zheng, X.; Wang, X.; Zhao, H.; Wang, T.; Zhang, H.; Li, W.; Shen, H.; Yu, L. Maternal Exposure to PM2.5 during Pregnancy Induces Impaired Development of Cerebral Cortex in Mice Offspring. Int. J. Mol. Sci. 2018, 19, 257. https://doi.org/10.3390/ijms19010257
Zhang T, Zheng X, Wang X, Zhao H, Wang T, Zhang H, Li W, Shen H, Yu L. Maternal Exposure to PM2.5 during Pregnancy Induces Impaired Development of Cerebral Cortex in Mice Offspring. International Journal of Molecular Sciences. 2018; 19(1):257. https://doi.org/10.3390/ijms19010257
Chicago/Turabian StyleZhang, Tianliang, Xinrui Zheng, Xia Wang, Hui Zhao, Tingting Wang, Hongxia Zhang, Wanwei Li, Hua Shen, and Li Yu. 2018. "Maternal Exposure to PM2.5 during Pregnancy Induces Impaired Development of Cerebral Cortex in Mice Offspring" International Journal of Molecular Sciences 19, no. 1: 257. https://doi.org/10.3390/ijms19010257
APA StyleZhang, T., Zheng, X., Wang, X., Zhao, H., Wang, T., Zhang, H., Li, W., Shen, H., & Yu, L. (2018). Maternal Exposure to PM2.5 during Pregnancy Induces Impaired Development of Cerebral Cortex in Mice Offspring. International Journal of Molecular Sciences, 19(1), 257. https://doi.org/10.3390/ijms19010257