Carbon Catabolite Repression in Filamentous Fungi

 , , ,
, , , 
Abstract
:1. Introduction
2. Sensing and Signaling Pathways of Carbon Catabolite Repression
2.1. Yeast
2.2. CCR of Filamentous Fungi and Differences with S. cerevisiae
3. Key Regulators of CCR in Filamentous Fungi
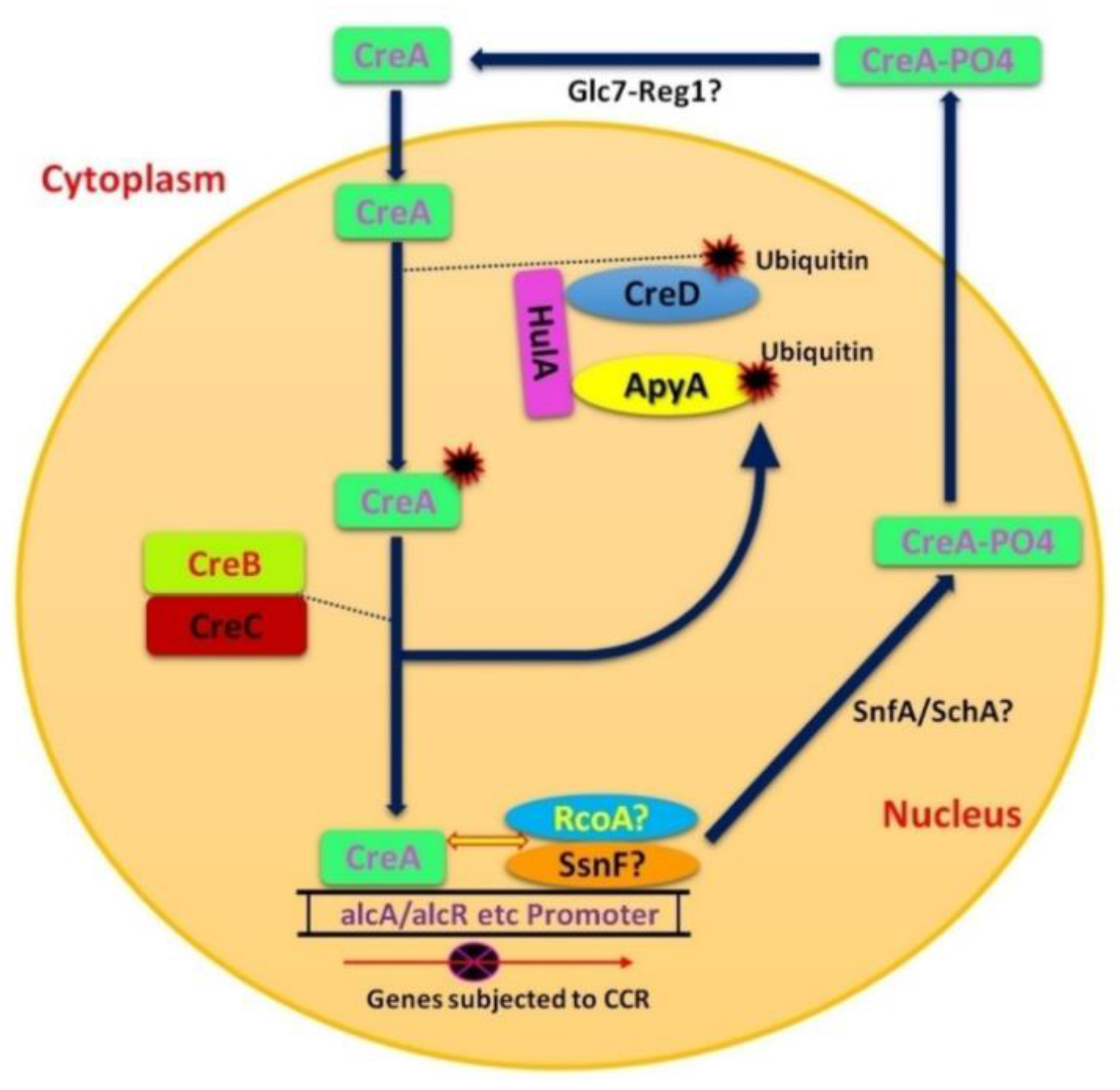
3.1. CreA/Cre1
3.2. CreB and CreC
3.3. CreD
3.4. Snf1
4. Chromatin Modification and CCR
5. Impact of CCR Components on Fungal Behavior
5.1. Regulation of Fungal Growth
5.2. Regulation/Utilization of Non-Glucose Sources
5.3. Virulence
6. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Katoh, H.; Ohtani, K.; Yamamoto, H.; Akimitsu, K. Overexpression of a gene encoding a catabolite repression element in Alternaria citri causes severe symptoms of black rot in citrus fruit. Phytopathology 2007, 97, 557–563. [Google Scholar] [CrossRef] [PubMed]
- New, A.M.; Cerulus, B.; Govers, S.K.; Perez-Samper, G.; Zhu, B.; Boogmans, S.; Xavier, J.B.; Verstrepen, K.J. Different levels of catabolite repression optimize growth in stable and variable environments. PLoS Biol. 2014, 12, e1001764. [Google Scholar] [CrossRef] [PubMed]
- Matar, K.A.O.; Chen, X.; Chen, D.; Anjago, W.M.; Norvienyeku, J.; Lin, Y.; Chen, M.; Wang, Z.; Ebbole, D.J.; Lu, G. WD40-repeat protein MoCreC is essential for carbon repression and is involved in conidiation, growth and pathogenicity of Magnaporthe oryzae. Curr. Genet. 2017, 63, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Portnoy, T.; Margeot, A.; Linke, R.; Atanasova, L.; Fekete, E.; Sándor, E.; Hartl, L.; Karaffa, L.; Druzhinina, I.S.; Seiboth, B. The CRE1 carbon catabolite repressor of the fungus Trichoderma reesei: A master regulator of carbon assimilation. BMC Genom. 2011, 12, 269. [Google Scholar] [CrossRef] [PubMed]
- Ries, L.N.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldman, G.H. Diverse regulation of the CreA carbon catabolite repressor in Aspergillus nidulans. Genet. 2016, 203, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Kayikci, Ö.; Nielsen, J. Glucose repression in Saccharomyces cerevisiae. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, J.; Nielsen, H.B.; Hofmann, G.; Nielsen, J. Transcription analysis using high-density micro-arrays of Aspergillus nidulans wild-type and creA mutant during growth on glucose or ethanol. Fungal Genet. Biol. 2006, 43, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Boase, N.A.; Kelly, J.M. A role for creD, a carbon catabolite repression gene from Aspergillus nidulans, in ubiquitination. Mol. Microbiol. 2004, 53, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Georgakopoulos, P.; Lockington, R.A.; Kelly, J.M. SAGA complex components and acetate repression in Aspergillus nidulans. G3 Genes Genom. Genet. 2012, 2, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Ichinose, S.; Tanaka, M.; Shintani, T.; Gomi, K. Improved α-amylase production by Aspergillus oryzae after a double deletion of genes involved in carbon catabolite repression. Appl. Microbiol. Biotechnol. 2014, 98, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.E.; Bernardo, S.M.; Cheetham, B.F. The interaction of induction, repression and starvation in the regulation of extracellular proteases in Aspergillus nidulans: Evidence for a role for CreA in the response to carbon starvation. Curr. Genet. 2008, 54, 47. [Google Scholar] [CrossRef] [PubMed]
- OBrian, G.R.; Fakhoury, A.M.; Payne, G.A. Identification of genes differentially expressed during aflatoxin biosynthesis in Aspergillus flavus and Aspergillus parasiticus. Fungal Genet. Biol. 2003, 39, 118–127. [Google Scholar] [CrossRef]
- Hunter, A.; Morris, T.; Jin, B.; Saint, C.P.; Kelly, J. Deletion of creB in Aspergillus oryzae increases secreted hydrolytic enzyme activity. Appl. Environ. Microbiol. 2013, 79, 5480–5487. [Google Scholar] [CrossRef] [PubMed]
- Amore, A.; Giacobbe, S.; Faraco, V. Regulation of cellulase and hemicellulase gene expression in fungi. Curr. Genom. 2013, 14, 230–249. [Google Scholar] [CrossRef] [PubMed]
- Tamayo, E.N.; Villanueva, A.; Hasper, A.A.; de Graaff, L.H.; Ramón, D.; Orejas, M. CreA mediates repression of the regulatory gene xlnR which controls the production of xylanolytic enzymes in Aspergillus nidulans. Fungal Genet. Biol. 2008, 45, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Kubicek, C.P.; Mikus, M.; Schuster, A.; Schmoll, M.; Seiboth, B. Metabolic engineering strategies for the improvement of cellulase production by Hypocrea jecorina. Biotechnol. Biofuels 2009, 2, 19. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Nielsen, J. Advances in metabolic pathway and strain engineering paving the way for sustainable production of chemical building blocks. Curr. Opin. Biotechnol. 2013, 24, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Liu, Y.; Guo, J.; Huang, L.; Zhang, X. Yeast synthetic biology for high-value metabolites. FEMS Yeast Res. 2015, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Borodina, I. Application of synthetic biology for production of chemicals in yeast Saccharomyces cerevisiae. FEMS Yeast Res 2015, 15. [Google Scholar] [CrossRef]
- Broach, J.R. Nutritional control of growth and development in yeast. Genet. 2012, 192, 73–105. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, G.M. Glucose signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 253–282. [Google Scholar] [CrossRef] [PubMed]
- Weinhandl, K.; Winkler, M.; Glieder, A.; Camattari, A. Carbon source dependent promoters in yeasts. Microb. Cell Factor. 2014, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Busti, S.; Coccetti, P.; Alberghina, L.; Vanoni, M. Glucose signaling-mediated coordination of cell growth and cell cycle in Saccharomyces cerevisiae. Sensors 2010, 10, 6195–6240. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Roy, A.; Jouandot, D.; Cho, K.H. The glucose signaling network in yeast. Biochim. Biophys. Acta-Gen. Subj. 2013, 1830, 5204–5210. [Google Scholar] [CrossRef] [PubMed]
- Peeters, K.; Thevelein, J.M. Glucose sensing and signal transduction in Saccharomyces cerevisiae. In Molecular Mechanisms in Yeast Carbon Metab; Springer: Berlin/Heidelberg, German, 2014; pp. 21–56. [Google Scholar]
- Tarinejad, A. Glucose Signalling in Saccharomyces cerevisiae. Agric. Commun. 2015, 3, 7–15. [Google Scholar]
- Tamanoi, F. Ras signaling in yeast. Genes Cancer 2011, 2, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Colombo, S.; Paiardi, C.; Pardons, K.; Winderickx, J.; Martegani, E. Evidence for adenylate cyclase as a scaffold protein for Ras2–Ira interaction in Saccharomyces cerevisie. Cell. Signal. 2014, 26, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Peeters, T.; Versele, M.; Thevelein, J.M. Directly from Gα to protein kinase A: The kelch repeat protein bypass of adenylate cyclase. Trends Biochem. Sci. 2007, 32, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Soulard, A.; Cremonesi, A.; Moes, S.; Schütz, F.; Jenö, P.; Hall, M.N. The rapamycin-sensitive phosphoproteome reveals that TOR controls protein kinase A toward some but not all substrates. Mol. Biol. Cell 2010, 21, 3475–3486. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.A.; de Gouvea, P.F.; Krohn, N.G.; Savoldi, M.; Goldman, G.H. Functional characterisation of the non-essential protein kinases and phosphatases regulating Aspergillus nidulans hydrolytic enzyme production. Biotechnol. Biofuels 2013, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozcan, S.; Johnston, M. Two different repressors collaborate to restrict expression of the yeast glucose transporter genes HXT2 and HXT4 to low levels of glucose. Mol. Cell. Biol. 1996, 16, 5536–5545. [Google Scholar] [CrossRef] [PubMed]
- Palomino, A.; Herrero, P.; Moreno, F. Tpk3 and Snf1 protein kinases regulate Rgt1 association with Saccharomyces cerevisiae HXK2 promoter. Nucleic Acids Res. 2006, 34, 1427–1438. [Google Scholar] [CrossRef] [PubMed]
- Ahuatzi, D.; Riera, A.; Peláez, R.; Herrero, P.; Moreno, F. Hxk2 regulates the phosphorylation state of Mig1 and therefore its nucleocytoplasmic distribution. J. Biol. Chem. 2007, 282, 4485–4493. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Tsuchiyama, S.; Yang, C.; Bulteau, A.L.; He, C.; Robison, B.; Tsuchiya, M.; Miller, D.; Briones, V.; Tar, K. Proteasomes, Sir2, and Hxk2 form an interconnected aging network that impinges on the AMPK/Snf1-regulated transcriptional repressor Mig1. PLoS Genet. 2015, 11, e1004968. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Johnston, M. Two glucose-sensing pathways converge on Rgt1 to regulate expression of glucose transporter genes in Saccharomyces cerevisiae. J. Biol. Chem. 2006, 281, 26144–26149. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kim, J.-H. Endocytosis and vacuolar degradation of the yeast cell surface glucose sensors Rgt2 and Snf3. J. Biol. Chem. 2016, 291, 14913. [Google Scholar] [CrossRef] [PubMed]
- Vega, M.; Riera, A.; Fernández-Cid, A.; Herrero, P.; Moreno, F. Hexokinase 2 is an intracellular glucose sensor of yeast cells that maintains the structure and activity of Mig1 protein repressor complex. J. Biol. Chem. 2016, 291, 7267–7285. [Google Scholar] [CrossRef] [PubMed]
- Rolland, F.; Winderickx, J.; Thevelein, J.M. Glucose-sensing and-signalling mechanisms in yeast. FEMS Yeast Res. 2002, 2, 183–201. [Google Scholar] [CrossRef] [PubMed]
- Welkenhuysen, N.; Borgqvist, J.; Backman, M.; Bendrioua, L.; Goksör, M.; Adiels, C.B.; Cvijovic, M.; Hohmann, S. Single-cell study links metabolism with nutrient signaling and reveals sources of variability. BMC Syst. Biol. 2017, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Otterstedt, K.; Larsson, C.; Bill, R.M.; Ståhlberg, A.; Boles, E.; Hohmann, S.; Gustafsson, L. Switching the mode of metabolism in the yeast Saccharomyces cerevisiae. EMBO Rep. 2004, 5, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Tabba, S.; Mangat, S.; McCartney, R.; Schmidt, M.C. PP1 phosphatase-binding motif in Reg1 protein of Saccharomyces cerevisiae is required for interaction with both the PP1 phosphatase Glc7 and the Snf1 protein kinase. Cell. Signal. 2010, 22, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Xu, X.; Carlson, M. Roles of two protein phosphatases, Reg1-Glc7 and Sit4, and glycogen synthesis in regulation of SNF1 protein kinase. Proc. Natl. Acad. Sci. USA 2011, 108, 6349–6354. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.V.; Heath, R.; Underwood, E.; Sanders, M.J.; Carmena, D.; McCartney, R.R.; Leiper, F.C.; Xiao, B.; Jing, C.; Walker, P.A. ADP regulates SNF1, the Saccharomyces cerevisiae homolog of AMP-activated protein kinase. Cell Metab. 2011, 14, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Vincent, O.; Kuchin, S.; Hong, S.-P.; Townley, R.; Vyas, V.K.; Carlson, M. Interaction of the Srb10 kinase with Sip4, a transcriptional activator of gluconeogenic genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 2001, 21, 5790–5796. [Google Scholar] [CrossRef] [PubMed]
- Ahuatzi, D.; Herrero, P.; De, L.C.T.; Moreno, F. The glucose-regulated nuclear localization of hexokinase 2 in Saccharomyces cerevisiae is Mig1-dependent. J. Biol. Chem. 2004, 279, 14440–14446. [Google Scholar] [CrossRef] [PubMed]
- Papamichos-Chronakis, M.; Petrakis, T.; Ktistaki, E.; Topalidou, I.; Tzamarias, D. Cti6, a PHD domain protein, bridges the Cyc8-Tup1 corepressor and the SAGA coactivator to overcome repression at GAL1. Mol. Cell 2002, 9, 1297–1305. [Google Scholar] [CrossRef]
- Papamichos-Chronakis, M.; Gligoris, T.; Tzamarias, D. The Snf1 kinase controls glucose repression in yeast by modulating interactions between the Mig1 repressor and the Cyc8-Tup1 co-repressor. EMBO Rep. 2004, 5, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Madi, L.; McBride, S.A.; Bailey, L.A.; Ebbole, D.J. rco-3, a gene involved in glucose transport and conidiation in Neurospora crassa. Genetics 1997, 146, 499–508. [Google Scholar] [PubMed]
- Flipphi, M.; van de Vondervoort, P.J.; Ruijter, G.J.; Visser, J.; Arst, H.N.; Felenbok, B. Onset of Carbon Catabolite Repression in Aspergillus nidulans. Parallel involvement of hexokinase and GLUCOKINASE in sugar signaling. J. Biol. Chem. 2003, 278, 11849–11857. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, S.M.; Gray, K.-A.; Todd, R.B.; Cheetham, B.F.; Katz, M.E. Characterization of regulatory non-catalytic hexokinases in Aspergillus nidulans. Mol. Genet. Genom. 2007, 277, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Doehlemann, G.; Berndt, P.; Hahn, M. Different signalling pathways involving a Gα protein, cAMP and a MAP kinase control germination of Botrytis cinerea conidia. Mol. Microbiol. 2006, 59, 821–835. [Google Scholar] [CrossRef] [PubMed]
- Lafon, A.; Seo, J.-A.; Han, K.-H.; Yu, J.-H.; d’Enfert, C. The heterotrimeric G-protein GanB (α)-SfaD (β)-GpgA (γ) is a carbon source sensor involved in early cAMP-dependent germination in Aspergillus nidulans. Genetics 2005, 171, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Zuber, S.; Hynes, M.J.; Andrianopoulos, A. The G-protein α-subunit GasC plays a major role in germination in the dimorphic fungus Penicillium marneffei. Genetics 2003, 164, 487–499. [Google Scholar] [PubMed]
- Xue, C.; Bahn, Y.-S.; Cox, G.M.; Heitman, J. G protein-coupled receptor Gpr4 senses amino acids and activates the cAMP-PKA pathway in Cryptococcus neoformans. Mol. Biol. Cell 2006, 17, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Alspaugh, J.A.; Perfect, J.R.; Heitman, J. Cryptococcus neoformans mating and virulence are regulated by the G-protein α subunit GPA1 and cAMP. Genes Dev. 1997, 11, 3206–3217. [Google Scholar] [CrossRef] [PubMed]
- Affeldt, K.J.; Brodhagen, M.; Keller, N.P. Aspergillus oxylipin signaling and quorum sensing pathways depend on G protein-coupled receptors. Toxins 2012, 4, 695–717. [Google Scholar] [CrossRef] [PubMed]
- Föge, M.; Heinekamp, T.; Brakhage, A. G protein-coupled receptors of the human-pathogenic fungus Aspergillus fumigatus. Mycoses 2012, 55, 141. [Google Scholar]
- Brown, N.A.; Dos Reis, T.F.; Ries, L.N.A.; Caldana, C.; Mah, J.H.; Yu, J.H.; Macdonald, J.M.; Goldman, G.H. G-protein coupled receptor-mediated nutrient sensing and developmental control in Aspergillus nidulans. Mol. Microbiol. 2015, 98, 420–439. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Borkovich, K.A. GPR-4 is a predicted G-protein-coupled receptor required for carbon source-dependent asexual growth and development in Neurospora crassa. Eukaryot.Cell 2006, 5, 1287–1300. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wright, S.J.; Krystofova, S.; Park, G.; Borkovich, K.A. Heterotrimeric G protein signaling in filamentous fungi. Annu. Rev. Microbiol. 2007, 61, 423–452. [Google Scholar] [CrossRef] [PubMed]
- De Souza, W.R.; Morais, E.R.; Krohn, N.G.; Savoldi, M.; Goldman, M.H.S.; Rodrigues, F.; Caldana, C.; Semelka, C.T.; Tikunov, A.P.; Macdonald, J.M. Identification of metabolic pathways influenced by the G-protein coupled receptors GprB and GprD in Aspergillus nidulans. PLoS ONE 2013, 8, e62088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krijgsheld, P.; Bleichrodt, R.; Van Veluw, G.; Wang, F.; Müller, W.; Dijksterhuis, J.; Wösten, H. Development in Aspergillus. Stud. Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Fuller, K.K.; Rhodes, J.C. Protein kinase A and fungal virulence: A sinister side to a conserved nutrient sensing pathway. Virulence 2012, 3, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Fuller, K.K.; Richie, D.L.; Feng, X.; Krishnan, K.; Stephens, T.J.; Wikenheiser-Brokamp, K.A.; Askew, D.S.; Rhodes, J.C. Divergent Protein Kinase A isoforms co-ordinately regulate conidial germination, carbohydrate metabolism and virulence in Aspergillus fumigatus. Mol. Microbiol. 2011, 79, 1045–1062. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.; Lockington, R.A.; Kelly, J.M. CreA-mediated repression in Aspergillus nidulans does not require transcriptional auto-regulation, regulated intracellular localisation or degradation of CreA. Fungal Genet. Biol. 2008, 45, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Fleck, C.B.; Brock, M. Aspergillus fumigatus catalytic glucokinase and hexokinase: Expression analysis and importance for germination, growth, and conidiation. Eukaryot. Cell 2010, 9, 1120–1135. [Google Scholar] [CrossRef] [PubMed]
- Madi, L.; Ebbole, D.J.; White, B.T.; Yanofsky, C. Mutants of Neurospora crassa that alter gene expression and conidia development. Proc. Natl. Acad. Sci. USA 1994, 91, 6226–6230. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, H.; Kristensen, J.B.; Felby, C. Enzymatic conversion of lignocellulose into fermentable sugars: Challenges and opportunities. Biofuels Bioprod. Biorefin. 2007, 1, 119–134. [Google Scholar] [CrossRef]
- Brown, N.A.; Ries, L.N.A.; Goldman, G.H. How nutritional status signalling coordinates metabolism and lignocellulolytic enzyme secretion. Fungal Genet. Biol. 2014, 72, 48–63. [Google Scholar] [CrossRef] [PubMed]
- Ubersax, J.A.; Ferrell, J.E., Jr. Mechanisms of specificity in protein phosphorylation. Nat. Rev. Mol. Cell Biol. 2007, 8, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Assis, L.J.; Ries, L.N.A.; Savoldi, M.; Reis, T.F.; Brown, N.A.; Goldman, G.H. Aspergillus nidulans protein kinase A plays an important role in cellulase production. Biotechnol. Biofuels 2015, 8, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.A.; Kamlangdee, N.; Kelly, J.M. The CreB deubiquitinating enzyme does not directly target the CreA repressor protein in Aspergillus nidulans. Curr. Genet. 2017, 63, 647–667. [Google Scholar] [CrossRef] [PubMed]
- García, I.; Mathieu, M.; Nikolaev, I.; Felenbok, B.; Scazzocchio, C. Roles of the Aspergillus nidulans homologues of Tup1 and Ssn6 in chromatin structure and cell viability. FEMS Microbiol. Lett. 2008, 289, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.; Lockington, R.A.; Strauss, J.; Dieringer, D.; Kubicek, C.P.; Kelly, J.; Keller, N. RcoA has pleiotropic effects on Aspergillus nidulans cellular development. Mol. Microbiol. 2001, 39, 1482–1493. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, C.T.; Ebbole, D.J.; Lee, B.-U.; Brown, R.E.; Bourland, C.; Madi, L.; Yanofsky, C. Characterization of rco-1 of Neurospora crassa, a pleiotropic gene affecting growth and development that encodes a homolog of Tup1 of Saccharomyces cerevisiae. Mol. Cell. Biol. 1996, 16, 6218–6228. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Ebbole, D.J. Tissue-Specific Repression of Starvation and Stress Responses of the Neurospora crassa con-10Gene Is Mediated by RCO1. Fungal Genet. Biol. 1998, 23, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Lockington, R.A.; Kelly, J.M. Carbon catabolite repression in Aspergillus nidulans involves deubiquitination. Mol. Microbiol. 2001, 40, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Lockington, R.A.; Kelly, J.M. The WD40-repeat protein CreC interacts with and stabilizes the deubiquitinating enzyme CreB in vivo in Aspergillus nidulans. Mol. Microbiol. 2002, 43, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Cupertino, F.B.; Virgilio, S.; Freitas, F.Z.; de Souza Candido, T.; Bertolini, M.C. Regulation of glycogen metabolism by the CRE-1, RCO-1 and RCM-1 proteins in Neurospora crassa. The role of CRE-1 as the central transcriptional regulator. Fungal Genet. Biol. 2015, 77, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Kelly, J.M. Proteins interacting with CreA and CreB in the carbon catabolite repression network in Aspergillus nidulans. Curr. Genet. 2017, 63, 669–683. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, C.; Benocci, T.; Battaglia, E.; Benoit, I.; de Vries, R.P. Chapter one-sugar catabolism in Aspergillus and other fungi related to the utilization of plant biomass. Adv. Appl. Microbiol. 2015, 90, 1–28. [Google Scholar] [PubMed]
- Ebbole, D.J. Carbon catabolite repression of gene expression and conidiation in Neurospora crassa. Fungal Genet. Biol. 1998, 25, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Tudzynski, B.; Liu, S.; Kelly, J.M. Carbon catabolite repression in plant pathogenic fungi: Isolation and characterization of the Gibberella fujikuroi and Botrytis cinerea creA genes. FEMS Microbiol. Lett. 2000, 184, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Tonukari, N.J.; Scott-Craig, J.S.; Walton, J.D. Isolation of the carbon catabolite repressor (CREA) gene from the plant-pathogenic fungus Cochliobolus carbonum. DNA Seq. 2003, 14, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Benocci, T.; Aguilar-Pontes, M.V.; Zhou, M.; Seiboth, B.; Vries, R.P. Regulators of plant biomass degradation in ascomycetous fungi. Biotechnol. Biofuels 2017, 10, 152. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Hikita, M. Anomalous Field-Induced Resistive Transition in La2-xSrxCuO4. Jpn. J. Appl. Phys. 1989, 28, L1368. [Google Scholar] [CrossRef]
- David, H.; Krogh, A.M.; Roca, C.; Åkesson, M.; Nielsen, J. CreA influences the metabolic fluxes of Aspergillus nidulans during growth on glucose and xylose. Microbiology 2005, 151, 2209–2221. [Google Scholar] [CrossRef] [PubMed]
- Klaubauf, S.; Narang, H.M.; Post, H.; Zhou, M.; Brunner, K.; Mach-Aigner, A.R.; Mach, R.L.; Heck, A.J.; Altelaar, A.M.; de Vries, R.P. Similar is not the same: Differences in the function of the (hemi-) cellulolytic regulator XlnR (Xlr1/Xyr1) in filamentous fungi. Fungal Genet. Biol. 2014, 72, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, G.J.; Visser, J. Carbon repression in Aspergilli. FEMS Microbiol. Lett. 1997, 151, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Macios, M.; Caddick, M.X.; Weglenski, P.; Scazzocchio, C.; Dzikowska, A. The GATA factors AREA and AREB together with the co-repressor NMRA, negatively regulate arginine catabolism in Aspergillus nidulans in response to nitrogen and carbon source. Fungal Genet. Biol. 2012, 49, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Hynes, M.J.; Todd, R.B.; Davis, M.A. Deletion and overexpression of the Aspergillus nidulans GATA factor AreB reveals unexpected pleiotropy. Microbiology 2009, 155, 3868–3880. [Google Scholar] [CrossRef] [PubMed]
- Delmas, S.; Pullan, S.T.; Gaddipati, S.; Kokolski, M.; Malla, S.; Blythe, M.J.; Ibbett, R.; Campbell, M.; Liddell, S. Uncovering the genome-wide transcriptional responses of the filamentous fungus Aspergillus niger to lignocellulose using RNA sequencing. PLoS Genet. 2012, 8, e1002875. [Google Scholar] [CrossRef] [PubMed]
- Znameroski, E.A.; Coradetti, S.T.; Roche, C.M.; Tsai, J.C.; Iavarone, A.T.; Cate, J.H.; Cate, J.H.; Glass, N.L. Induction of lignocellulose-degrading enzymes in Neurospora crassa by cellodextrins. Proc. Natl. Acad. Sci. USA 2012, 109, 6012–6017. [Google Scholar] [CrossRef] [PubMed]
- Cziferszky, A.; Mach, R.L.; Kubicek, C.P. Phosphorylation positively regulates DNA binding of the carbon catabolite repressor Cre1 of Hypocrea jecorina (Trichoderma reesei). J. Biol. Chem. 2002, 277, 14688–14694. [Google Scholar] [CrossRef] [PubMed]
- Cziferszky, A.; Seiboth, B.; Kubicek, C.P. The Snf1 kinase of the filamentous fungus Hypocrea jecorina phosphorylates regulation-relevant serine residues in the yeast carbon catabolite repressor Mig1 but not in the filamentous fungal counterpart Cre1. Fungal Genet. Biol. 2003, 40, 166–175. [Google Scholar] [CrossRef]
- Mathieu, M.; Fillinger, S.; Felenbok, B. In Vivo studies of upstream regulatory cis-acting elements of the alcR gene encoding the transactivator of the ethanol regulon in Aspergillus nidulans. Mol. Microbiol. 2000, 36, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Nikolaev, I.; Scazzocchio, C.; Felenbok, B. Patterns of nucleosomal organization in the alc regulon of Aspergillus nidulans: Roles of the AlcR transcriptional activator and the CreA global repressor. Mol. Microbiol. 2005, 56, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Mach-Aigner, A.R.; Pucher, M.E.; Steiger, M.G.; Bauer, G.E.; Preis, S.J.; Mach, R.L. Transcriptional regulation of xyr1, encoding the main regulator of the xylanolytic and cellulolytic enzyme system in Hypocrea jecorina. Appl. Environ. Microbiol. 2008, 74, 6554–6562. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.; van de Vondervoort, P.; Hendriks, L.; Van de Belt, M.; Visser, J. Regulation of the a-glucuronidase-encoding gene (aguA) from Aspergillus niger. Mol. Genet. Genom. 2002, 268, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Tian, C.; Diamond, S.; Glass, N.L. Deciphering transcriptional regulatory mechanisms associated with hemicellulose degradation in Neurospora crassa. Eukaryot. Cell 2012, 11, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Glass, N.L. Identification of the CRE-1 cellulolytic regulon in Neurospora crassa. PLoS ONE 2011, 6, e25654. [Google Scholar] [CrossRef] [PubMed]
- Nitta, M.; Furukawa, T.; Shida, Y.; Mori, K.; Kuhara, S.; Morikawa, Y.; Ogasawara, W. A new Zn (II) 2 Cys 6-type transcription factor BglR regulates β-glucosidase expression in Trichoderma reesei. Fungal Genet. Biol. 2012, 49, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Sun, J.; Glass, N.L. VIB1, a link between glucose signaling and carbon catabolite repression, is essential for plant cell wall degradation by Neurospora crassa. PLoS Genet. 2014, 10, e1004500. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Inoue, H.; Ishikawa, K. Decreased cellulase and xylanase production in the fungus Talaromyces cellulolyticus by disruption of tacA and tctA genes, encoding putative zinc finger transcriptional factors. Appl. Biochem. Biotechnol. 2015, 175, 3218–3229. [Google Scholar] [CrossRef] [PubMed]
- Coradetti, S.T.; Craig, J.P.; Xiong, Y.; Shock, T.; Tian, C.; Glass, N.L. Conserved and essential transcription factors for cellulase gene expression in ascomycete fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 7397–7402. [Google Scholar] [CrossRef] [PubMed]
- Craig, J.P.; Coradetti, S.T.; Starr, T.L.; Glass, N.L. Direct target network of the Neurospora crassa plant cell wall deconstruction regulators CLR-1, CLR-2, and XLR-1. MBio 2015, 6, e01452-15. [Google Scholar] [CrossRef] [PubMed]
- Raulo, R.; Kokolski, M.; Archer, D.B. The roles of the zinc finger transcription factors XlnR, ClrA and ClrB in the breakdown of lignocellulose by Aspergillus niger. AMB Exp. 2016, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Aro, N.; Saloheimo, A.; Ilmén, M.; Penttilä, M. ACEII, a novel transcriptional activator involved in regulation of cellulase and xylanase genes of Trichoderma reesei. J. Biol. Chem. 2001, 276, 24309–24314. [Google Scholar] [CrossRef] [PubMed]
- Häkkinen, M.; Valkonen, M.J.; Westerholm-Parvinen, A.; Aro, N.; Arvas, M.; Vitikainen, M.; Penttilä, M.; Saloheimo, M.; Pakula, T.M. Screening of candidate regulators for cellulase and hemicellulase production in Trichoderma reesei and identification of a factor essential for cellulase production. Biotechnol. Biofuels 2014, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, Y.; Endo, Y.; Li, N.; Yoshizawa, M.; Aoyama, M.; Watanabe, A.; Kanamaru, K.; Kato, M.; Kobayashi, T. Regulation of cellulolytic genes by McmA, the SRF-MADS box protein in Aspergillus nidulans. Biochem. Biophys. Res. Commun. 2013, 431, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Saloheimo, A.; Aro, N.; Ilmén, M.; Penttilä, M. Isolation of the ace1 gene encoding a Cys2-His2 transcription factor involved in regulation of activity of the cellulase promoter cbh1of Trichoderma reesei. J. Biol. Chem. 2000, 275, 5817–5825. [Google Scholar] [CrossRef] [PubMed]
- Chilton, I.J.; Delaney, C.; Barham-Morris, J.; Fincham, D.A.; Hooley, P.; Whitehead, M.P. The Aspergillus nidulans stress response transcription factor StzA is ascomycete-specific and shows species-specific polymorphisms in the C-terminal region. Mycol. Res. 2008, 112, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, M.; Tian, C.; Sun, J.; Tisch, D.; Glass, N.L. Unravelling the molecular basis for light modulated cellulase gene expression-the role of photoreceptors in Neurospora crassa. BMC Genom. 2012, 13, 127. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.; Loros, J.J.; Dunlap, J.C. Chapter two-dissecting the mechanisms of the clock in Neurospora. Methods Enzymol. 2015, 551, 29–52. [Google Scholar] [PubMed]
- Dasgupta, A.; Fuller, K.K.; Dunlap, J.C.; Loros, J.J. Seeing the world differently: Variability in the photosensory mechanisms of two model fungi. Environ. Microbiol. 2016, 18, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lin, L.; Li, H.; Tian, C.; Ma, Y. Transcriptional comparison of the filamentous fungus Neurospora crassa growing on three major monosaccharides D-glucose, D-xylose and L-arabinose. Biotechnol. Biofuels 2014, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, E.; Visser, L.; Nijssen, A.; van Veluw, G.; Wösten, H.; de Vries, R. Analysis of regulation of pentose utilisation in Aspergillus niger reveals evolutionary adaptations in Eurotiales. Stud. Mycol. 2011, 69, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, E.; Zhou, M.; de Vries, R.P. The transcriptional activators AraR and XlnR from Aspergillus niger regulate expression of pentose catabolic and pentose phosphate pathway genes. Res. Microbiol. 2014, 165, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Klaubauf, S.; Zhou, M.; Lebrun, M.H.; Vries, R.P.; Battaglia, E. A novel l-arabinose-responsive regulator discovered in the rice-blast fungus Pyricularia oryzae (Magnaporthe oryzae). FEBS Lett. 2016, 590, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Gomi, K.; Akeno, T.; Minetoki, T.; Ozeki, K.; Kumagai, C.; Okazaki, N.; Iimura, Y. Molecular cloning and characterization of a transcriptional activator gene, amyR, involved in the amylolytic gene expression in Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2000, 64, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Benen, J.A.; Wösten, H.A.; Visser, J.; de Vries, R.P. A broader role for AmyR in Aspergillus niger: Regulation of the utilisation of D-glucose or D-galactose containing oligo-and polysaccharides. Appl. Microbiol. Biotechnol. 2012, 93, 285–293. [Google Scholar]
- Hasegawa, S.; Takizawa, M.; Suyama, H.; Shintani, T.; Gomi, K. Characterization and expression analysis of a maltose-utilizing (MAL) cluster in Aspergillus oryzae. Fungal Genet. Biol. 2010, 47, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Tanaka, M.; Konno, Y.; Ichikawa, T.; Ichinose, S.; Hasegawa-Shiro, S.; Shintani, T.; Gomi, K. Distinct mechanism of activation of two transcription factors, AmyR and MalR, involved in amylolytic enzyme production in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2015, 99, 1805–1815. [Google Scholar] [CrossRef] [PubMed]
- Kunitake, E.; Tani, S.; Sumitani, J.-I.; Kawaguchi, T. A novel transcriptional regulator, ClbR, controls the cellobiose-and cellulose-responsive induction of cellulase and xylanase genes regulated by two distinct signaling pathways in Aspergillus aculeatus. Appl. Microbiol. Biotechnol. 2013, 97, 2017–2028. [Google Scholar] [CrossRef] [PubMed]
- Kunitake, E.; Kawamura, A.; Tani, S.; Takenaka, S.; Ogasawara, W.; Sumitani, J.-I.; Kawaguchi, T. Effects of clbR overexpression on enzyme production in Aspergillus aculeatus vary depending on the cellulosic biomass-degrading enzyme species. Biosci. Biotechnol. Biochem. 2015, 79, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, G.; Liu, W. Multiple metabolic signals influence GAL gene activation by modulating the interaction of Gal80p with the transcriptional activator Gal4p. Mol. Microbiol. 2010, 78, 414–428. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, J.E.; Gruben, B.S.; Battaglia, E.; Wiebenga, A.; Majoor, E.; de Vries, R.P. Genetic Interaction of Aspergillus nidulans galR, xlnR and araR in regulating D-Galactose and L-Arabinose release and catabolism gene expression. PLoS ONE 2015, 10, e0143200. [Google Scholar] [CrossRef] [PubMed]
- Stricker, A.R.; Steiger, M.G.; Mach, R.L. Xyr1 receives the lactose induction signal and regulates lactose metabolism in Hypocrea jecorina. FEBS Lett. 2007, 581, 3915–3920. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.-L.; Roubos, J.A.; van den Hondel, C.A.; Ram, A.F. Identification of InuR, a new Zn (II) 2Cys6 transcriptional activator involved in the regulation of inulinolytic genes in Aspergillus niger. Mol. Genet. Genom. 2008, 279, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lubbers, R.J.; Simon, A.; Stassen, J.H.; Vargas Ribera, P.R.; Viaud, M.; van Kan, J.A. A novel Zn2Cys6 transcription factor BcGaaR regulates D-galacturonic acid utilization in Botrytis cinerea. Mol. Microbiol. 2016, 100, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Derntl, C.; Rassinger, A.; Srebotnik, E.; Mach, R.L.; Mach-Aigner, A.R. Xpp1 regulates the expression of xylanases, but not of cellulases in Trichoderma reesei. Biotechnol. Biofuels 2015, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Derntl, C.; Kluger, B.; Bueschl, C.; Schuhmacher, R.; Mach, R.L.; Mach-Aigner, A.R. Transcription factor Xpp1 is a switch between primary and secondary fungal metabolism. Proc. Natl. Acad. Sci. USA 2017, 114, E560–E569. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, R. A survey of 178 NF-Y binding CCAAT boxes. Nucleic Acids Res. 1998, 26, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Zeilinger, S.; Mach, R.L.; Kubicek, C.P. Two adjacent protein binding motifs in the cbh2 (cellobiohydrolase II-encoding) promoter of the fungus Hypocrea jecorina (Trichoderma reesei) cooperate in the induction by cellulose. J. Biol. Chem. 1998, 273, 34463–34471. [Google Scholar] [CrossRef] [PubMed]
- Narendja, F.M.; Davis, M.A.; Hynes, M.J. AnCF, the CCAAT Binding Complex of Aspergillus nidulans, Is Essential for the Formation of a DNase I-Hypersensitive Site in the 5′ Region of theamdS Gene. Mol. Cell. Biol. 1999, 19, 6523–6531. [Google Scholar] [CrossRef] [PubMed]
- Steidl, S.; Papagiannopoulos, P.; Litzka, O.; Andrianopoulos, A.; Davis, M.A.; Brakhage, A.A.; Hynes, M.J. AnCF, the CCAAT Binding Complex of Aspergillus nidulans, Contains Products of the hapB, hapC, and hapE Genes and Is Required for Activation by the Pathway-Specific Regulatory GeneamdR. Mol. Cell. Biol. 1999, 19, 99–106. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.P.; Jansen, J.; Aguilar, G.; Pařenicová, L.; Joosten, V.; Wülfert, F.; Benen, J.A.; Visser, J. Expression profiling of pectinolytic genes from Aspergillus niger. FEBS Lett. 2002, 530, 41–47. [Google Scholar] [CrossRef]
- Gruben, B.S.; Zhou, M.; Wiebenga, A.; Ballering, J.; Overkamp, K.M.; Punt, P.J.; de Vries, R.P. Aspergillus niger RhaR, a regulator involved in L-rhamnose release and catabolism. Appl. Microbiol. Biotechnol. 2014, 98, 5531–5540. [Google Scholar] [CrossRef] [PubMed]
- Christensen, U.; Gruben, B.S.; Madrid, S.; Mulder, H.; Nikolaev, I.; de Vries, R.P. Unique regulatory mechanism for D-galactose utilization in Aspergillus nidulans. Appl. Environ. Microbiol. 2011, 77, 7084–7087. [Google Scholar] [CrossRef] [PubMed]
- Culleton, H.; McKie, V.; de Vries, R.P. Physiological and molecular aspects of degradation of plant polysaccharides by fungi: What have we learned from Aspergillus? Biotechnol. J. 2013, 8, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Todd, R.; Lockington, R.; Kelly, J. The Aspergillus nidulans creC gene involved in carbon catabolite repression encodes a WD40 repeat protein. Mol. Gen. Genet. 2000, 263, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Neer, E.J.; Schmidt, C.J.; Nambudripad, R.; Smith, T.F. The ancient regulatory-protein family of WD-repeat proteins. Nature 1994, 371, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Denton, J.A.; Kelly, J.M. Disruption of Trichoderma reesei cre2, encoding an ubiquitin C-terminal hydrolase, results in increased cellulase activity. BMC Biotechnol. 2011, 11, 103. [Google Scholar] [CrossRef] [PubMed]
- Hynes, M.; Kelly, J.M. Pleiotropic mutants of Aspergillus nidulans altered in carbon metabolism. Mol. Gen. Genet. 1977, 150, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.I.; Einbond, A.; Kwak, S.-J.; Linn, H.; Koepf, E.; Peterson, S.; Kelly, J.W.; Sudol, M. Characterization of the WW domain of human yes-associated protein and its polyproline-containing ligands. J. Biol. Chem. 1997, 272, 17070–17077. [Google Scholar] [CrossRef] [PubMed]
- Kiekhaefer, C.M.; Boyer, M.E.; Johnson, K.D.; Bresnick, E.H. A WW domain-binding motif within the activation domain of the hematopoietic transcription factor NF-E2 is essential for establishment of a tissue-specific histone modification pattern. J. Biol. Chem. 2004, 279, 7456–7461. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Kus, B.; Fladd, C.; Wasmuth, J.; Tonikian, R.; Sidhu, S.; Krogan, N.J.; Parkinson, J.; Rotin, D. Ubiquitination screen using protein microarrays for comprehensive identification of Rsp5 substrates in yeast. Mol. Syst. Biol. 2007, 3, 116. [Google Scholar] [CrossRef] [PubMed]
- Carlson, M.; Osmond, B.C.; Botstein, D. Mutants of yeast defective in sucrose utilization. Genetics 1981, 98, 25–40. [Google Scholar] [PubMed]
- Hardie, D.G. AMP-activated/SNF1 protein kinases: Conserved guardians of cellular energy. Nat. Rev. Mol. Cell Biol. 2007, 8, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G.; Carling, D.; Carlson, M. The AMP-activated/SNF1 protein kinase subfamily: Metabolic sensors of the eukaryotic cell? Annu. Rev. 1998, 67, 821–855. [Google Scholar] [CrossRef] [PubMed]
- García-Salcedo, R.; Lubitz, T.; Beltran, G.; Elbing, K.; Tian, Y.; Frey, S.; Wolkenhauer, O.; Krantz, M.; Klipp, E.; Hohmann, S. Glucose de-repression by yeast AMP-activated protein kinase SNF1 is controlled via at least two independent steps. FEBS J. 2014, 281, 1901–1917. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Park, J.-H.; Ahn, J.-H.; Lee, Y.-H. MoSNF1 regulates sporulation and pathogenicity in the rice blast fungus Magnaporthe oryzae. Fungal Genet. Biol. 2008, 45, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Brosch, G.; Loidl, P.; Graessle, S. Histone modifications and chromatin dynamics: A focus on filamentous fungi. FEMS Microbiol. Rev. 2008, 32, 409–439. [Google Scholar] [CrossRef] [PubMed]
- Soyer, J.L.; El Ghalid, M.; Glaser, N.; Ollivier, B.; Linglin, J.; Grandaubert, J.; Balesdent, M.H.; Connolly, L.R.; Freitag, M.; Rouxel, T. Epigenetic control of effector gene expression in the plant pathogenic fungus Leptosphaeria maculans. PLoS Genet. 2014, 10, e1004227. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Jin, G.; Zhou, X. Modeling the relationship of epigenetic modifications to transcription factor binding. Nucleic Acids Res. 2015, 43, 3873–3885. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Karimi-Aghcheh, R.; Bok, J.W.; Phatale, P.A.; Smith, K.M.; Baker, S.E.; Lichius, A.; Omann, M.; Zeilinger, S.; Seiboth, B.; Rhee, C. Functional analyses of Trichoderma reesei LAE1 reveal conserved and contrasting roles of this regulator. G3 Gen. Genomes Genet. 2013, 3, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, A.R.; Németh, Z.; Atanasova, L.; Fekete, E.; Paholcsek, M. The VELVET A Orthologue VEL1 of Trichoderma reesei regulates fungal development and is essential for cellulase gene expression. PLoS ONE 2014, 9, e112799. [Google Scholar]
- Fekete, E.; Karaffa, L.; Aghcheh, R.K.; Németh, Z.; Fekete, É.; Orosz, A.; Paholcsek, M.; Stágel, A.; Kubicek, C.P. The transcriptome of lae1 mutants of Trichoderma reesei cultivated at constant growth rates reveals new targets of LAE1 function. BMC Genom. 2014, 15, 447. [Google Scholar] [CrossRef] [PubMed]
- Zeilinger, S.; Schmoll, M.; Pail, M.; Mach, R.L.; Kubicek, C.P. Nucleosome transactions on the Hypocrea jecorina (Trichoderma reesei) cellulase promoter cbh2 associated with cellulase induction. Mol. Genet. Genom. 2003, 270, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Ronne, H. Glucose repression in fungi. Trends Genet. 1995, 11, 12–17. [Google Scholar] [CrossRef]
- García, I.; Gonzalez, R.; Gómez, D.; Scazzocchio, C. Chromatin rearrangements in the prnD-prnB bidirectional promoter: Dependence on transcription factors. Eukaryot. Cell 2004, 3, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Ries, L.; Belshaw, N.; Ilmén, M.; Penttilä, M.; Alapuranen, M.; Archer, D. The role of CRE1 in nucleosome positioning within the cbh1 promoter and coding regions of Trichoderma reesei. Appl. Microbiol. Biotechnol. 2014, 98, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Mello-de-Sousa, T.M.; Rassinger, A.; Pucher, M.E.; dos Santos Castro, L.; Persinoti, G.F.; Silva-Rocha, R.; Poças-Fonseca, M.J.; Mach, R.L.; Silva, R.N.; Mach-Aigner, A.R. The impact of chromatin remodelling on cellulase expression in Trichoderma reesei. BMC Genom. 2015, 16, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mello-de-Sousa, T.M.; Rassinger, A.; Derntl, C.; Poças-Fonseca, M.J.; Mach, R.L.; R Mach-Aigner, A. The relation between promoter chromatin status, Xyr1 and cellulase expression in Trichoderma reesei. Curr. Genom. 2016, 17, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Xin, Q.; Gong, Y.; Lv, X.; Chen, G.; Liu, W. Trichoderma reesei histone acetyltransferase Gcn5 regulates fungal growth, conidiation, and cellulase gene expression. Curr. Microbiol. 2013, 67, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Aro, N.; Pakula, T.; Penttilä, M. Transcriptional regulation of plant cell wall degradation by filamentous fungi. FEMS Microbiol. Rev. 2005, 29, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Ilyés, H.; Fekete, E.; Karaffa, L.; Fekete, É.; Sándor, E.; Szentirmai, A.; Kubicek, C.P. CreA-mediated carbon catabolite repression of β-galactosidase formation in Aspergillus nidulans is growth rate dependent. FEMS Microbiol. Lett. 2004, 235, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Mc Mahon, H.; Kelly, C.; Fogarty, W. Effect of growth rate on α-amylase production by Streptomyces sp. IMD 2679. Appl. Microbiol. Biotechnol. 1997, 48, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Van Hoek, P.; Van Dijken, J.P.; Pronk, J.T. Effect of specific growth rate on fermentative capacity of baker’s yeast. Appl. Environ. Microbiol. 1998, 64, 4226–4233. [Google Scholar] [PubMed]
- Warner, J.B.; Lolkema, J.S. CcpA-dependent carbon catabolite repression in bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.; Horvath, H.K.; Abdallah, B.M.; Kindermann, J.; Mach, R.L.; Kubicek, C.P. The function of CreA, the carbon catabolite repressor of Aspergillus nidulans, is regulated at the transcriptional and post-transcriptional level. Mol. Microbiol. 1999, 32, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Kuchin, S.; Vyas, V.K.; Carlson, M. Snf1 protein kinase and the repressors Nrg1 and Nrg2 regulate FLO11, haploid invasive growth, and diploid pseudohyphal differentiation. Mol. Cell. Biol. 2002, 22, 3994–4000. [Google Scholar] [CrossRef] [PubMed]
- Somerville, C.; Bauer, S.; Brininstool, G.; Facette, M.; Hamann, T.; Milne, J.; Osborne, E.; Paredez, A.; Persson, S.; Raab, T. Toward a systems approach to understanding plant cell walls. Science 2004, 306, 2206–2211. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.; Somerville, C. Cellulosic biofuels. Annu. Rev. Plant Biol. 2009, 60, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, E. Regulation and Diversity of Plant Polysaccharide Utilisation in Fungi. Ph.D. Thesis, University Utrecht, Utrecht, The Netherlands, 2011. [Google Scholar]
- Kowalczyk, J.E.; Benoit, I.; de Vries, R.P. Regulation of plant biomass utilization in Aspergillus. Adv. Appl. Microbiol. 2014, 88, 31–56. [Google Scholar] [PubMed]
- Mäkelä, M.R.; Donofrio, N.; de Vries, R.P. Plant biomass degradation by fungi. Fungal Genet. Biol. 2014, 72, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Tonukari, N.J.; Scott-Craig, J.S.; Waltonb, J.D. The Cochliobolus carbonum SNF1 gene is required for cell wall–degrading enzyme expression and virulence on maize. Plant Cell 2000, 12, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Ospina-Giraldo, M.D.; Mullins, E.; Kang, S. Loss of function of the Fusarium oxysporum SNF1 gene reduces virulence on cabbage and Arabidopsis. Curr. Genet. 2003, 44, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Hedbacker, K.; Carlson, M. SNF1/AMPK pathways in yeast. Front. Biosci. A J. Virtual Libr. 2008, 13, 2408. [Google Scholar] [CrossRef]
- Orejas, M.; MacCabe, A.P.; Pérez González, J.A.; Kumar, S.; Ramón, D. Carbon catabolite repression of the Aspergillus nidulans xlnA gene. Mol. Microbiol. 1999, 31, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Jonkers, W.; Rep, M. Mutation of CRE1 in Fusarium oxysporum reverts the pathogenicity defects of the FRP1 deletion mutant. Mol. Microbiol. 2009, 74, 1100–1113. [Google Scholar] [CrossRef] [PubMed]
- Mach-Aigner, A.R.; Omony, J.; Jovanovic, B.; van Boxtel, A.J.; de Graaff, L.H. D-Xylose concentration-dependent hydrolase expression profiles and the function of CreA and XlnR in Aspergillus niger. Appl. Environ. Microbiol. 2012, 78, 3145–3155. [Google Scholar] [CrossRef] [PubMed]
- Felenbok, B.; Flipphi, M.; Nikolaev, I. Ethanol catabolism in Aspergillus nidulans: A model system for studying gene regulation. Prog.Nucleic Acid Res. Mol. Biol. 2001, 69, 149–204. [Google Scholar] [PubMed]
- Fillinger, S.; Felenbok, B. A newly identified gene cluster in Aspergillus nidulans comprises five novel genes localized in the alc region that are controlled both by the specific transactivator AlcR and the general carbon-catabolite repressor CreA. Mol. Microbiol. 1996, 20, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Flipphi, M.; Mathieu, M.; Cirpus, I.; Panozzo, C.; Felenbok, B. Regulation of the aldehyde dehydrogenase gene (aldA) and its role in the control of the coinducer level necessary for induction of the ethanol utilization pathway in Aspergillus nidulans. J. Biol. Chem. 2001, 276, 6950–6958. [Google Scholar] [CrossRef] [PubMed]
- Flipphi, M.; Kocialkowska, J.; Felenbok, B. Characteristics of physiological inducers of the ethanol utilization (alc) pathway in Aspergillus nidulans. Biochem. J. 2002, 364, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Felenbok, B. The Aspergillus nidulans CREA protein mediates glucose repression of the ethanol regulon at various levels through competition with the ALCR-specific transactivator. EMBO J. 1994, 13, 4022. [Google Scholar] [PubMed]
- Panozzo, C.; Cornillot, E.; Felenbok, B. The CreA Repressor Is the Sole DNA-binding Protein Responsible for Carbon Catabolite Repression of the alcA Gene in Aspergillus nidulans via Its Binding to a Couple of Specific Sites. J. Biol. Chem. 1998, 273, 6367–6372. [Google Scholar] [CrossRef] [PubMed]
- Takashima, S.; Iikura, H.; Nakamura, A.; Masaki, H.; Uozumi, T. Analysis of Cre1 binding sites in the Trichoderma reesei cbh1 upstream region. FEMS Microbiol. Lett. 1996, 145, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Portnoy, T.; Margeot, A.; Seidl-Seiboth, V.; Le Crom, S.; Chaabane, F.B.; Linke, R.; Seiboth, B.; Kubicek, C.P. Differential regulation of the cellulase transcription factors XYR1, ACE2, and ACE1 in Trichoderma reesei strains producing high and low levels of cellulase. Eukaryot. Cell 2011, 10, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Antoniêto, A.C.C.; dos Santos Castro, L.; Silva-Rocha, R.; Persinoti, G.F.; Silva, R.N. Defining the genome-wide role of CRE1 during carbon catabolite repression in Trichoderma reesei using RNA-Seq analysis. Fungal Genet. Biol. 2014, 73, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Lorito, M.; Mach, R.L.; Sposato, P.; Strauss, J.; Peterbauer, C.K.; Kubicek, C.P. Mycoparasitic interaction relieves binding of the Cre1 carbon catabolite repressor protein to promoter sequences of the ech42 (endochitinase-encoding) gene in Trichoderma harzianum. Proc. Natl. Acad. Sci. USA 1996, 93, 14868–14872. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Pava-Ripoll, M.; Wang, S.; Leger, R.S. Protein kinase A regulates production of virulence determinants by the entomopathogenic fungus, Metarhizium anisopliae. Fungal Genet. Biol. 2009, 46, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Leng, B.; Xiao, Y.; Jin, K.; Ma, J.; Fan, Y.; Feng, J.; Yang, X.; Zhang, Y.; Pei, Y. Cloning of Beauveria bassiana chitinase gene Bbchit1 and its application to improve fungal strain virulence. Appl. Environ. Microbiol. 2005, 71, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Screen, S.; Bailey, A.; Charnley, K.; Cooper, R.; Clarkson, J. Carbon regulation of the cuticle-degrading enzyme PR1 from Metarhizium anisopliae may involve a trans-acting DNA-binding protein CRR1, a functional equivalent of the Aspergillus nidulans CREA protein. Curr. Genet. 1997, 31, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Baksi, S.; Koris, A.; Vatai, G. Journey of enzymes in entomopathogenic fungi. Pac. Sci. Rev. A Nat. Sci. Eng. 2016, 18, 85–99. [Google Scholar] [CrossRef]
- Jin, K.; Luo, Z.; Jiang, X.; Zhang, Y.; Zhou, Y.; Pei, Y. Carbon catabolite repressor gene BbCre1 influences carbon source uptake but does not have a big impact on virulence in Beauveria bassiana. J. Invertebr. Pathol. 2011, 106, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.A.; Gibson, R.P.; Quispe, C.F.; Littlechild, J.A.; Talbot, N.J. An NADPH-dependent genetic switch regulates plant infection by the rice blast fungus. Proc. Natl. Acad. Sci. USA 2010, 107, 21902–21907. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.; Wright, J.D.; Hartline, D.; Quispe, C.F.; Madayiputhiya, N.; Wilson, R.A. Principles of carbon catabolite repression in the rice blast fungus: Tps1, Nmr1-3, and a MATE–Family Pump regulate glucose metabolism during Infection. PLoS Genet. 2012, 8, e1002673. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Kim, K.H.; La Rota, M.; Scott, D.; Santopietro, G.; Callihan, M.; Mitchell, T.K.; Lawrence, C.B. Identification of novel virulence factors associated with signal transduction pathways in Alternaria brassicicola. Mol. Microbiol. 2009, 72, 1316–1333. [Google Scholar] [CrossRef] [PubMed]
- Tzima, A.K.; Paplomatas, E.J.; Rauyaree, P.; Ospina-Giraldo, M.D.; Kang, S. VdSNF1, the sucrose nonfermenting protein kinase gene of Verticillium dahliae, is required for virulence and expression of genes involved in cell-wall degradation. Mol. Plant-Microbe Interact. 2011, 24, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Nakari-Setälä, T.; Paloheimo, M.; Kallio, J.; Vehmaanperä, J.; Penttilä, M.; Saloheimo, M. Genetic modification of carbon catabolite repression in Trichoderma reesei for improved protein production. Appl. Environ. Microbiol. 2009, 75, 4853–4860. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Inoue, H.; Ishikawa, K. Enhancing cellulase and hemicellulase production by genetic modification of the carbon catabolite repressor gene, creA, in Acremonium cellulolyticus. AMB Exp. 2013, 3, 73. [Google Scholar] [CrossRef] [PubMed]
- Bi, F.; Barad, S.; Ment, D.; Luria, N.; Dubey, A.; Casado, V.; Glam, N.; Mínguez, J.D.; Espeso, E.A.; Fluhr, R. Carbon regulation of environmental pH by secreted small molecules that modulate pathogenicity in phytopathogenic fungi. Mol. Plant Pathol. 2016, 17, 1178–1195. [Google Scholar] [CrossRef] [PubMed]
- Beattie, S.R.; Mark, K.M.; Thammahong, A.; Ries, L.N.A.; Dhingra, S.; Caffrey-Carr, A.K.; Cheng, C.; Black, C.C.; Bowyer, P.; Bromley, M.J. Filamentous fungal carbon catabolite repression supports metabolic plasticity and stress responses essential for disease progression. PLoS Pathog. 2017, 13, e1006340. [Google Scholar] [CrossRef] [PubMed]
- Vinuselvi, P.; Kim, M.-K.; Lee, S.-K.; Ghim, C.-M. Rewiring carbon catabolite repression for microbial cell factory. BMB Rep. 2012, 45, 59–70. [Google Scholar] [CrossRef] [PubMed]

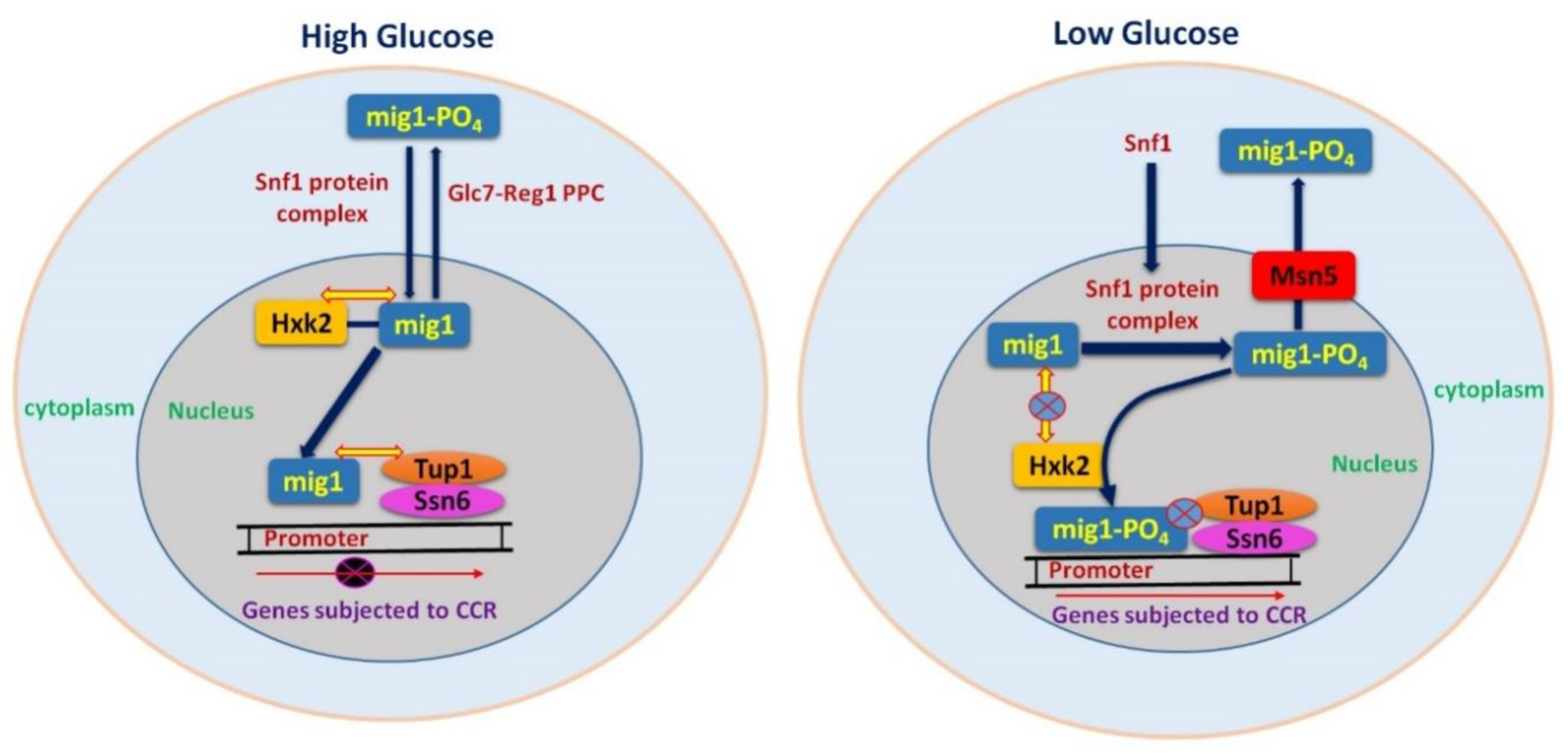
 represents protein interaction in the nucleus, mig1 and hxk2, Ssn6-Tup1 co-repressor complex and
represents protein interaction in the nucleus, mig1 and hxk2, Ssn6-Tup1 co-repressor complex and  mig1. Shows that there is no protein interaction, mig1 and hxk2, Ssn6-Tup1 co-repressor complex and mig1 and between Ssn6-Tup1 co-repressor complex and promoter gene.
mig1. Shows that there is no protein interaction, mig1 and hxk2, Ssn6-Tup1 co-repressor complex and mig1 and between Ssn6-Tup1 co-repressor complex and promoter gene.  represents the inhibition of transcription [34,45,46,47,48]. During high glucose conditions, inactive Snf1 cannot cause phosphorylation of Mig1 and cellular movement of Mig1 is dependent on Glc7–Reg1 complex; whereas during low glucose levels Snf1 will be active and cause direct repression of Snf1, which will be unable to repress the CCR subjected genes.
represents protein interaction in the nucleus, mig1 and hxk2, Ssn6-Tup1 co-repressor complex and mig1. Shows that there is no protein interaction, mig1 and hxk2, Ssn6-Tup1 co-repressor complex and mig1 and between Ssn6-Tup1 co-repressor complex and promoter gene. represents the inhibition of transcription [34,45,46,47,48]. During high glucose conditions, inactive Snf1 cannot cause phosphorylation of Mig1 and cellular movement of Mig1 is dependent on Glc7–Reg1 complex; whereas during low glucose levels Snf1 will be active and cause direct repression of Snf1, which will be unable to repress the CCR subjected genes.
represents the inhibition of transcription [34,45,46,47,48]. During high glucose conditions, inactive Snf1 cannot cause phosphorylation of Mig1 and cellular movement of Mig1 is dependent on Glc7–Reg1 complex; whereas during low glucose levels Snf1 will be active and cause direct repression of Snf1, which will be unable to repress the CCR subjected genes.
represents protein interaction in the nucleus, mig1 and hxk2, Ssn6-Tup1 co-repressor complex and mig1. Shows that there is no protein interaction, mig1 and hxk2, Ssn6-Tup1 co-repressor complex and mig1 and between Ssn6-Tup1 co-repressor complex and promoter gene. represents the inhibition of transcription [34,45,46,47,48]. During high glucose conditions, inactive Snf1 cannot cause phosphorylation of Mig1 and cellular movement of Mig1 is dependent on Glc7–Reg1 complex; whereas during low glucose levels Snf1 will be active and cause direct repression of Snf1, which will be unable to repress the CCR subjected genes.


{kind=link}
{kind=link}
{kind=link}
| Nature/Kind of Glucose Sensor | Sensor/Sensors in S. cerevisiae | Sensor/Sensors in Filamentous Fungi | Function in Filamentous Fungi |
|---|---|---|---|
| Transporter | Snf3, Rgt2 | Rco3 (N. crassa) | Rco3 in N. crassa might perform the same role as compared to yeast proteins but structure of Rco3 is different. It may have wider role than yeast proteins in CCR which only regulate hexose transporters [49]. |
| Hexokinase | Glk2, Hxk1, Hxk2 | HxkA, GlkA (A. nidulans) | No evidence showed direct role of a single hexokinase like yeast to start CCR but phosphorylation is required [50]. |
| HxkC, HxkD (A. nidulans) | Show response in carbon starved conditions while not involved in CCR [51]. | ||
| G-protein coupled receptor | Gpr1, Gpa2 (Gα3) | Gpr4, GCNA1-3 (Gα1-3) (N. crassa) | In response to glucose the G protein coupled receptor GPR4 play role in cAMP signaling. While GPR4 in yeast interacts with Gα1 instead of Gα3, and Gα1 through Gα3 play role in nutrient sensing. |
| BCG3 (Gα3) (Botrytis cinerea) | During germinating conidia has function in cAMP signaling/carbon sensing [52]. | ||
| GanB (Gα3) (A. nidulans) | During germinating conidia has function in cAMP signaling/carbon sensing [53]. | ||
| GasC (Gα3) (Penicillium marneffei) | Has no role in nutrient sensing but essential for germinating conidia [54]. | ||
| Gpr4 (C. neoformans) | Homologous to yeast Gpr1 but has no role glucose signaling [55]. | ||
| Gpa1 (Gα3) (Cryptococcus neoformans) | Involved in cAMP signaling in response to glucose and melanin production [56]. |
| Transcription Factor | Major Role | Fungi |
|---|---|---|
| CreA/CRE1 | CCR | Aspergillus spp., N. crassa, Trichoderma spp., etc. [70] |
| BglR/COL-26 | Sugar sensing and regulation of glucosidase expression | N. crassa, Trichoderma reesei [103,104,105]. |
| VIB1 | C-derepression, may also play role in cellulases induction | N. crassa [104] |
| CLR-1/ClrA | Cellulose utilization | N. crassa, Aspergillus spp. [106,107,108]. |
| CLR-2/ClrB/ManR | Cellulose utilization | N. crassa, Aspergillus spp., Talaromyces cellulolyticus, Peniciulium oxalicum [106,107,108] |
| ACE2, ACE3 | Cellulose utilization | T. reesei [109,110] |
| McmA | Cellulase regulation | A. nidulans [111] |
| ACE1 | Cellulase repression | T. reesei [112,113] |
| XlnR/XLR1/XYR1 | Hemi-cellulose utilization | Aspergillus spp., Fusarium spp., Trichoderma spp., M. oryzae, Protubera canescens, N. crassa, T. cellulolyticus [89] |
| WC-1/BLR1, WC-2/BLR2 | Hemi-cellulose utilization | Trichoderma spp., N. crassa [114,115,116] |
| HCR-1 | Hemi-cellulase repressor | N. crassa [117] |
| AraR | l-Arabinose utilization | Aspergillus spp. [118,119] |
| ARA1 | l-Arabinose utilization | M. oryzae [120] |
| AmyR | Starch utilization | Aspergillus spp. [121,122] |
| MalR | Maltose utilization | Aspergillus spp. [123,124] |
| ClbR | Cellobiose utilization | Aspergillus aculeatus [125,126] |
| GalR | d-Galactose utilization | A. nidulans [127,128] |
| GalX | d-Galactose utilization | Aspergillus spp. [129] |
| GaaR | Galacturonic acid utilization | B. cinerea, Aspergillus niger [130,131]. |
| GaaX | Galacturonic acid repressor | A. niger [131] |
| XPP1 | Xylanases repressor | T. reesei [132,133] |
| HAP complex | Carbohydrate-Active enZymes regulation (CAZy regulation) | Aspergillus spp., Trichoderma spp., N. crassa [134,135,136,137] |
| RhaR | l-Rhamnose utilization | Aspergillus spp. [138,139] |
| InuR | Inulin utilization | A. niger [140,141] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adnan, M.; Zheng, W.; Islam, W.; Arif, M.; Abubakar, Y.S.; Wang, Z.; Lu, G. Carbon Catabolite Repression in Filamentous Fungi. Int. J. Mol. Sci. 2018, 19, 48. https://doi.org/10.3390/ijms19010048
Adnan M, Zheng W, Islam W, Arif M, Abubakar YS, Wang Z, Lu G. Carbon Catabolite Repression in Filamentous Fungi. International Journal of Molecular Sciences. 2018; 19(1):48. https://doi.org/10.3390/ijms19010048
Chicago/Turabian StyleAdnan, Muhammad, Wenhui Zheng, Waqar Islam, Muhammad Arif, Yakubu Saddeeq Abubakar, Zonghua Wang, and Guodong Lu. 2018. "Carbon Catabolite Repression in Filamentous Fungi" International Journal of Molecular Sciences 19, no. 1: 48. https://doi.org/10.3390/ijms19010048
APA StyleAdnan, M., Zheng, W., Islam, W., Arif, M., Abubakar, Y. S., Wang, Z., & Lu, G. (2018). Carbon Catabolite Repression in Filamentous Fungi. International Journal of Molecular Sciences, 19(1), 48. https://doi.org/10.3390/ijms19010048