Metabolic Effects of Oral Phenelzine Treatment on High-Sucrose-Drinking Mice


Abstract
:1. Introduction
2. Results
2.1. Oral Chronic Phenelzine Treatment Has No Effect on Body Weight Gain and Adiposity of Sucrose-Drinking Mice
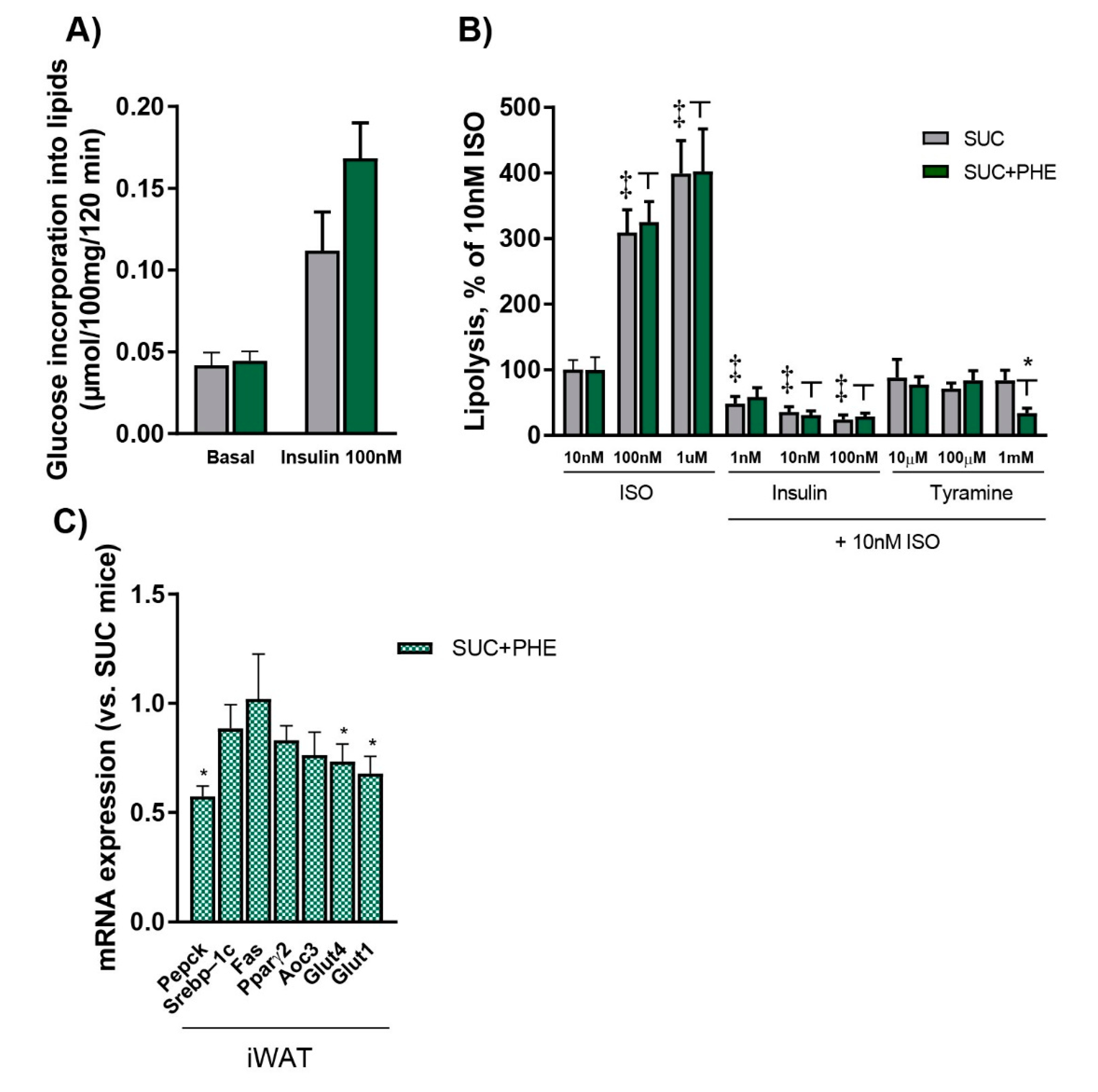
2.2. Chronic Phenelzine Treatment Does Not Alter Lipogenic and Lipolytic Activities, but Reduces Pepck Expression in the iWAT of Sucrose-Drinking Mice
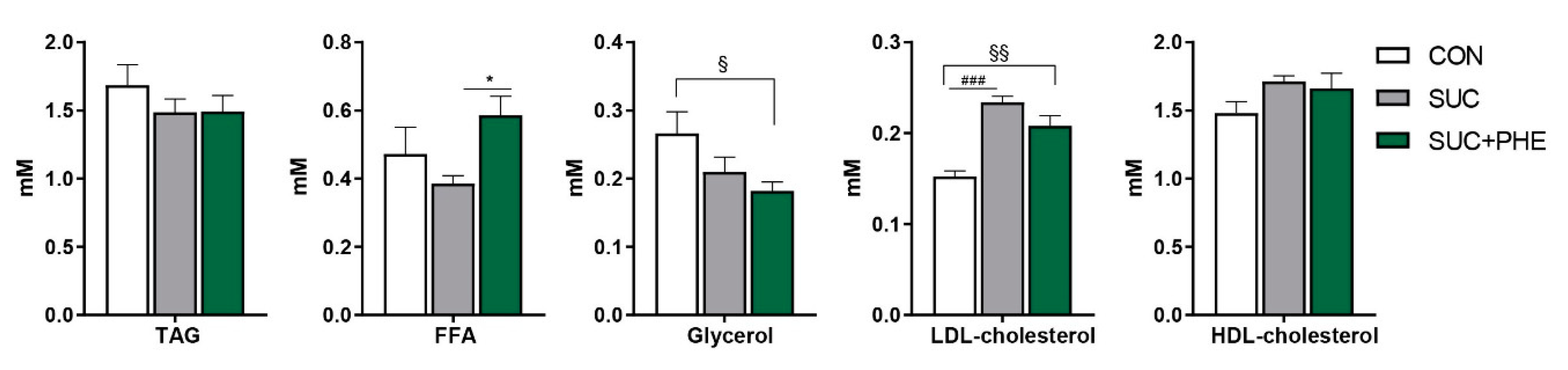
2.3. Impact of Phenelzine Treatment on Circulating Parameters Related to Lipid Metabolism in Sucrose-Drinking Mice
2.4. Phenelzine Treatment Restores Fasting Glycemia at the Expense of Increasing Insulinemia in Sucrose-Drinking Mice
2.5. Phenelzine Treatment Increases Rectal Temperature, but Appears to Be Unrelated to BAT Thermogenic and Muscle Catabolic Capacity
2.6. Effects of Phenelzine Treatment on Indicators of Oxidative Stress and Inflammation
3. Discussion
4. Materials and Methods
4.1. Animals, Diets, and Treatments
4.2. In Vivo Determinations and Sacrifice
4.3. Circulating Parameters and Tissue Composition Analysis
4.4. Lipogenesis and Lipolysis
4.5. Gene Expression
4.6. Oxidative Stress Biomarkers
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Song, M.S.; Matveychuk, D.; MacKenzie, E.M.; Duchcherer, M.; Mousseau, D.D.; Baker, G.B. An update on amine oxidase inhibitors: Multifaceted drugs. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 44, 118–124. [Google Scholar] [CrossRef] [PubMed]
- McKenna, K.F.; Baker, G.B.; Coutts, R.T.; Greenshaw, A.J. Chronic administration of the antidepressant-antipanic drug phenelzine and its N-acetylated analogue: Effects on monoamine oxidase, biogenic amines, and alpha 2-adrenoreceptor function. J. Pharm. Sci. 1992, 81, 832–835. [Google Scholar] [CrossRef] [PubMed]
- Matveychuk, D.; Nunes, E.; Ullah, N.; Velázquez-Martinez, C.A.; MacKenzie, E.M.; Baker, G.B. Comparison of phenelzine and geometric isomers of its active metabolite, β-phenylethylidenehydrazine, on rat brain levels of amino acids, biogenic amine neurotransmitters and methylamine. J. Neural Transm. (Vienna) 2013, 120, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Morin, N.; Lizcano, J.M.; Fontana, E.; Marti, L.; Smih, F.; Rouet, P.; Prévot, D.; Zorzano, A.; Unzeta, M.; Carpéné, C. Semicarbazide-sensitive amine oxidase substrates stimulate glucose transport and inhibit lipolysis in human adipocytes. J. Pharmacol. Exp. Ther. 2001, 297, 563–572. [Google Scholar] [PubMed]
- Mercader, J.; Iffiú-Soltész, Z.; Bour, S.; Carpéné, C. Oral Administration of Semicarbazide Limits Weight Gain together with Inhibition of Fat Deposition and of Primary Amine Oxidase Activity in Adipose Tissue. J. Obes. 2011, 2011, 475786. [Google Scholar] [CrossRef] [PubMed]
- Carpéné, C.; Iffiú-Soltesz, Z.; Bour, S.; Prévot, D.; Valet, P. Reduction of fat deposition by combined inhibition of monoamine oxidases and semicarbazide-sensitive amine oxidases in obese Zucker rats. Pharmacol. Res. 2007, 56, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Carpéné, C.; Abello, V.; Iffiú-Soltész, Z.; Mercier, N.; Fève, B.; Valet, P. Limitation of adipose tissue enlargement in rats chronically treated with semicarbazide-sensitive amine oxidase and monoamine oxidase inhibitors. Pharmacol. Res. 2008, 57, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Carpéné, C.; Gomez-Zorita, S.; Gupta, R.; Grès, S.; Rancoule, C.; Cadoudal, T.; Mercader, J.; Gomez, A.; Bertrand, C.; Iffiu-Soltész, Z. Combination of low dose of the anti-adipogenic agents resveratrol and phenelzine in drinking water is not sufficient to prevent obesity in very-high-fat diet-fed mice. Eur. J. Nutr. 2014, 53, 1625–1635. [Google Scholar] [CrossRef] [PubMed]
- Carpéné, C.; Grès, S.; Rascalou, S. The amine oxidase inhibitor phenelzine limits lipogenesis in adipocytes without inhibiting insulin action on glucose uptake. J. Neural Transm. (Vienna) 2013, 120, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Chiche, F.; Le Guillou, M.; Chétrite, G.; Lasnier, F.; Dugail, I.; Carpéné, C.; Moldes, M.; Fève, B. Antidepressant phenelzine alters differentiation of cultured human and mouse preadipocytes. Mol. Pharmacol. 2009, 75, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Byun, Y.; Park, J.; Hong, S.H.; Han, M.H.; Park, S.; Jung, H.I.; Noh, M. The opposite effect of isotype-selective monoamine oxidase inhibitors on adipogenesis in human bone marrow mesenchymal stem cells. Bioorg. Med. Chem. Lett. 2013, 23, 3273–3276. [Google Scholar] [CrossRef] [PubMed]
- Carpéné, C.; Mercader, J.; Le Gonidec, S.; Schaak, S.; Mialet-Perez, J.; Zakaroff-Girard, A.; Galitzky, J. Body fat reduction without cardiovascular changes in mice after oral treatment with the MAO inhibitor phenelzine. Br. J. Pharmacol. 2018, 175, 2428–2440. [Google Scholar] [CrossRef] [PubMed]
- Gillman, P.K. Advances pertaining to the pharmacology and interactions of irreversible nonselective monoamine oxidase inhibitors. J. Clin. Psychopharmacol. 2011, 31, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Finberg, J.P.; Gillman, K. Selective inhibitors of monoamine oxidase type B and the “cheese effect”. Int. Rev. Neurobiol. 2011, 100, 169–190. [Google Scholar] [PubMed]
- McIntyre, R.S.; Soczynska, J.K.; Konarski, J.Z.; Kennedy, S.H. The effect of antidepressants on glucose homeostasis and insulin sensitivity: Synthesis and mechanisms. Expert Opin. Drug Saf. 2006, 5, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Haeckel, R.; Terlutter, H.; Schumann, G.; Oellerich, M. Hydrazonopropionic acids, a new class of hypoglycemic substances, 3. Inhibition of jejunal glucose uptake in the rat and guinea pig. Horm. Metab. Res. 1984, 16, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Haeckel, R.; Oellerich, M. The influence of hydrazine, phenelzine and nialamide on gluconeogenesis and cell respiration in the perfused guinea-pig liver. Eur. J. Clin. Investig. 1977, 7, 393–400. [Google Scholar] [CrossRef]
- Feldman, J.M.; Chapman, B. Monoamine oxidase inhibitors: Nature of their interaction with rabbit pancreatic islets to alter insluin secretion. Diabetologia 1975, 11, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Pannecoeck, R.; Serruys, D.; Benmeridja, L.; Delanghe, J.R.; van Geel, N.; Speeckaert, R.; Speeckaert, M.M. Vascular adhesion protein-1: Role in human pathology and application as a biomarker. Crit. Rev. Clin. Lab. Sci. 2015, 52, 284–300. [Google Scholar] [CrossRef] [PubMed]
- Iffiú-Soltész, Z.; Mercader, J.; Daviaud, D.; Boucher, J.; Carpéné, C. Increased primary amine oxidase expression and activity in white adipose tissue of obese and diabetic db-/- mice. J. Neural Transm. (Vienna) 2011, 118, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Hamann, K.; Nehrt, G.; Ouyang, H.; Duerstock, B.; Shi, R. Hydralazine inhibits compression and acrolein-mediated injuries in ex vivo spinal cord. J. Neurochem. 2008, 104, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.N.; Gilmer, L.K.; Miller, D.M.; Cebak, J.E.; Wang, J.A.; Hall, E.D. Phenelzine mitochondrial functional preservation and neuroprotection after traumatic brain injury related to scavenging of the lipid peroxidation-derived aldehyde 4-hydroxy-2-nonenal. J. Cereb. Blood Flow Metab. 2013, 33, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.G.; Al-Shboul, O.; Alfaqih, M.A.; Al-Qudah, M.A.; Al-Dwairi, A.N. Phenelzine reduces the oxidative damage induced by peroxynitrite in plasma lipids and proteins. Arch. Physiol. Biochem. 2017, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Galvani, S.; Coatrieux, C.; Elbaz, M.; Grazide, M.H.; Thiers, J.C.; Parini, A.; Uchida, K.; Kamar, N.; Rostaing, L.; Baltas, M.; et al. Carbonyl scavenger and antiatherogenic effects of hydrazine derivatives. Free Radic. Biol. Med. 2008, 45, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Marques-Lopes, I.; Forga, L.; Martínez, J.A. Thermogenesis induced by a high-carbohydrate meal in fasted lean and overweight young men: Insulin, body fat, and sympathetic nervous system involvement. Nutrition 2003, 19, 25–29. [Google Scholar] [CrossRef]
- de França, S.A.; dos Santos, M.P.; Przygodda, F.; Garófalo, M.A.; Kettelhut, I.C.; Magalhães, D.A.; Bezerra, K.S.; Colodel, E.M.; Flouris, A.D.; Andrade, C.M.; et al. A Low-Protein, High-Carbohydrate Diet Stimulates Thermogenesis in the Brown Adipose Tissue of Rats via ATF-2. Lipids 2016, 51, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Carmo, H.; Remião, F.; Carvalho, F.; Fernandes, E.; de Boer, D.; dos Reys, L.A.; de Lourdes Bastos, M. 4-Methylthioamphetamine-induced hyperthermia in mice: Influence of serotonergic and catecholaminergic pathways. Toxicol. Appl. Pharmacol. 2003, 190, 262–271. [Google Scholar] [CrossRef]
- Takeda, M.; Imaizumi, M.; Sawano, S.; Manabe, Y.; Fushiki, T. Long-term optional ingestion of corn oil induces excessive caloric intake and obesity in mice. Nutrition 2001, 17, 117–120. [Google Scholar] [CrossRef]
- Jürgens, H.; Haass, W.; Castañeda, T.R.; Schürmann, A.; Koebnick, C.; Dombrowski, F.; Otto, B.; Nawrocki, A.R.; Scherer, P.E.; Spranger, J.; et al. Consuming fructose-sweetened beverages increases body adiposity in mice. Obes. Res. 2005, 13, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.C.; Wang, S.Y.; Chen, G.C.; Lin, Y.S.; Chao, P.M. The functional assessment of Alpinia pricei on metabolic syndrome induced by sucrose-containing drinking water in mice. Phytother. Res. 2009, 23, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.D.; Lin, P.Y. Zinc-induced hyperleptinemia relates to the amelioration of sucrose-induced obesity with zinc repletion. Obes. Res. 2000, 8, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.S.; Lopes, K.L.; Freitas, R.e.A.; de Oliveira-Sales, E.B.; Bergasmaschi, C.T.; Campos, R.R.; Casarini, D.E.; Carmona, A.K.; Araújo, M.a.S.; Heimann, J.C.; et al. High sucrose intake in rats is associated with increased ACE2 and angiotensin-(1-7) levels in the adipose tissue. Regul. Pept. 2010, 162, 61–67. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, R.S.; Soczynska, J.K.; Konarski, J.Z.; Kennedy, S.H. The effect of antidepressants on lipid homeostasis: A cardiac safety concern? Expert Opin. Drug Saf. 2006, 5, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Mazzio, E.; Soliman, K.F. The role of glycolysis and gluconeogenesis in the cytoprotection of neuroblastoma cells against 1-methyl 4-phenylpyridinium ion toxicity. Neurotoxicology 2003, 24, 137–147. [Google Scholar] [CrossRef]
- Aleyassine, H.; Gardiner, R.J. Dual action of antidepressant drugs (MAO inhibitors) on insulin release. Endocrinology 1975, 96, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Sturza, A.; Duicu, O.M.; Vaduva, A.; Dănilă, M.D.; Noveanu, L.; Varró, A.; Muntean, D.M. Monoamine oxidases are novel sources of cardiovascular oxidative stress in experimental diabetes. Can. J. Physiol. Pharmacol. 2015, 93, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- DOLE, V.P.; MEINERTZ, H. Microdetermination of long-chain fatty acids in plasma and tissues. J. Biol. Chem. 1960, 235, 2595–2599. [Google Scholar] [PubMed]
- Grès, S.; Canteiro, S.; Mercader, J.; Carpéné, C. Oxidation of high doses of serotonin favors lipid accumulation in mouse and human fat cells. Mol. Nutr. Food Res. 2013, 57, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Panchuk-Voloshina, N. A one-step fluorometric method for the continuous measurement of monoamine oxidase activity. Anal. Biochem. 1997, 253, 169–174. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | CON | SUC | SUC + PHE | p Value |
|---|---|---|---|---|
| Body weight at baseline (g) | 28.3 ± 1.2 | 28.2 ± 0.6 | 28.2 ± 0.8 | |
| Body weight at week 11 (g) | 32.6 ± 1.4 | 36.1 ± 0.7 # | 34.1 ± 0.6 | 0.032 |
| Ac. chow intake (g) | 238 ± 17 | 231 ± 8 | ||
| Ac. sucrose intake (g) | 37.5 ± 1.2 | 32.1 ± 2.5 | ||
| Ac. energy intake (kcal) | 910 ± 54 | 867 ± 35 | ||
| Energy efficiency (g/Mcal) | 8.5 ± 1.3 | 8.3 ± 3 | ||
| Adiposomatic index (%) | 6.0 ± 0.5 | 8.0 ± 0.7 | 7.2 ± 0.7 | |
| pWAT | ||||
| mass (%) | 1.0 ± 0.1 | 1.3 ± 0.1 | 1.1 ± 0.1 | |
| eWAT | ||||
| mass (%) | 3.0 ± 0.2 | 3.6 ± 0.3 | 3.2 ± 0.3 | |
| lipid (mg) | 692 ± 83 | 1166 ± 119 # | 996 ± 105 | 0.023 |
| protein (mg) | 10.2 ± 1.4 | 10.2 ± 0.4 | 9.5 ± 1.6 | |
| DNA (µg) | 46.4 ± 5.4 | 40.8 ± 4.6 | 33.3 ± 2.9 | |
| iWAT | ||||
| mass (%) | 1.9 ± 0.1 | 2.8 ± 0.2 # | 2.4 ± 0.2 | 0.043 |
| lipid (mg) | 431 ± 51 | 782 ± 89 # | 676 ± 89 | 0.031 |
| protein (mg) | 9.3 ± 1.5 | 11.4 ± 1.3 | 12.9 ± 1.8 | |
| DNA (µg) | 35.1 ± 3.0 | 58.2 ± 4.9 # | 63.1 ± 6.4 §§ | 0.003 |
| iBAT | ||||
| mass (%) | 0.31 ± 0.03 | 0.51 ± 0.04 # | 0.41 ± 0.04 | 0.032 |
| Liver | ||||
| mass (%) | 4.2 ± 0.3 | 4.8 ± 0.1 # | 4.8 ± 0.02 § | 0.013 |
| lipid (mg) | 51.6 ± 6.7 | 57.2 ± 11.0 | 66.7 ± 8.3 | |
| protein (mg) | 248 ± 2 | 277 ± 15 | 247 ± 12 | |
| DNA (µg) | 2177 ± 211 | 2088 ± 132 | 1803 ± 84 | |
| TAG (mg/g tissue) | 0.050 ± 0.003 | 0.168 ± 0.021 # | 0.175 ± 0.019 §§ | 0.008 |
| Skeletal muscle | ||||
| lipid (mg/g tissue) | 17.3 ± 2.6 | 23.2 ± 2.9 | 23.5 ± 2.8 | |
| Heart | ||||
| mass (%) | 0.45 ± 0.03 | 0.47 ± 0.01 | 0.44 ± 0.01 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carpéné, C.; Gómez-Zorita, S.; Chaplin, A.; Mercader, J. Metabolic Effects of Oral Phenelzine Treatment on High-Sucrose-Drinking Mice. Int. J. Mol. Sci. 2018, 19, 2904. https://doi.org/10.3390/ijms19102904
Carpéné C, Gómez-Zorita S, Chaplin A, Mercader J. Metabolic Effects of Oral Phenelzine Treatment on High-Sucrose-Drinking Mice. International Journal of Molecular Sciences. 2018; 19(10):2904. https://doi.org/10.3390/ijms19102904
Chicago/Turabian StyleCarpéné, Christian, Saioa Gómez-Zorita, Alice Chaplin, and Josep Mercader. 2018. "Metabolic Effects of Oral Phenelzine Treatment on High-Sucrose-Drinking Mice" International Journal of Molecular Sciences 19, no. 10: 2904. https://doi.org/10.3390/ijms19102904
APA StyleCarpéné, C., Gómez-Zorita, S., Chaplin, A., & Mercader, J. (2018). Metabolic Effects of Oral Phenelzine Treatment on High-Sucrose-Drinking Mice. International Journal of Molecular Sciences, 19(10), 2904. https://doi.org/10.3390/ijms19102904