Modulation of Insulin Resistance and the Adipocyte-Skeletal Muscle Cell Cross-Talk by LCn-3PUFA



Abstract
:
1. Introduction
2. Results
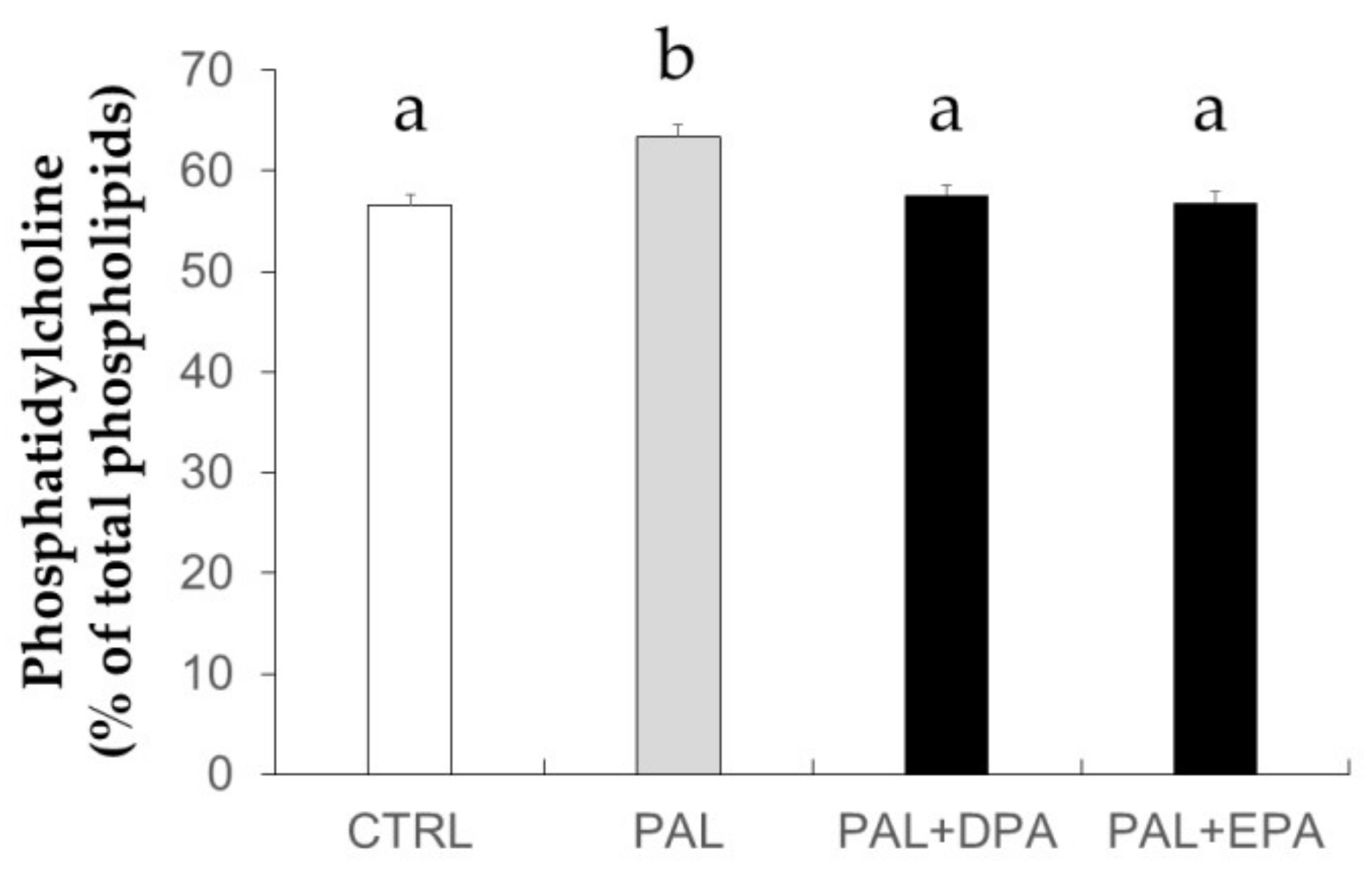
2.1. Effects of LCn-3PUFA Treatments on Membrane Phosphatidylcholine Content and Fatty Acid Profile
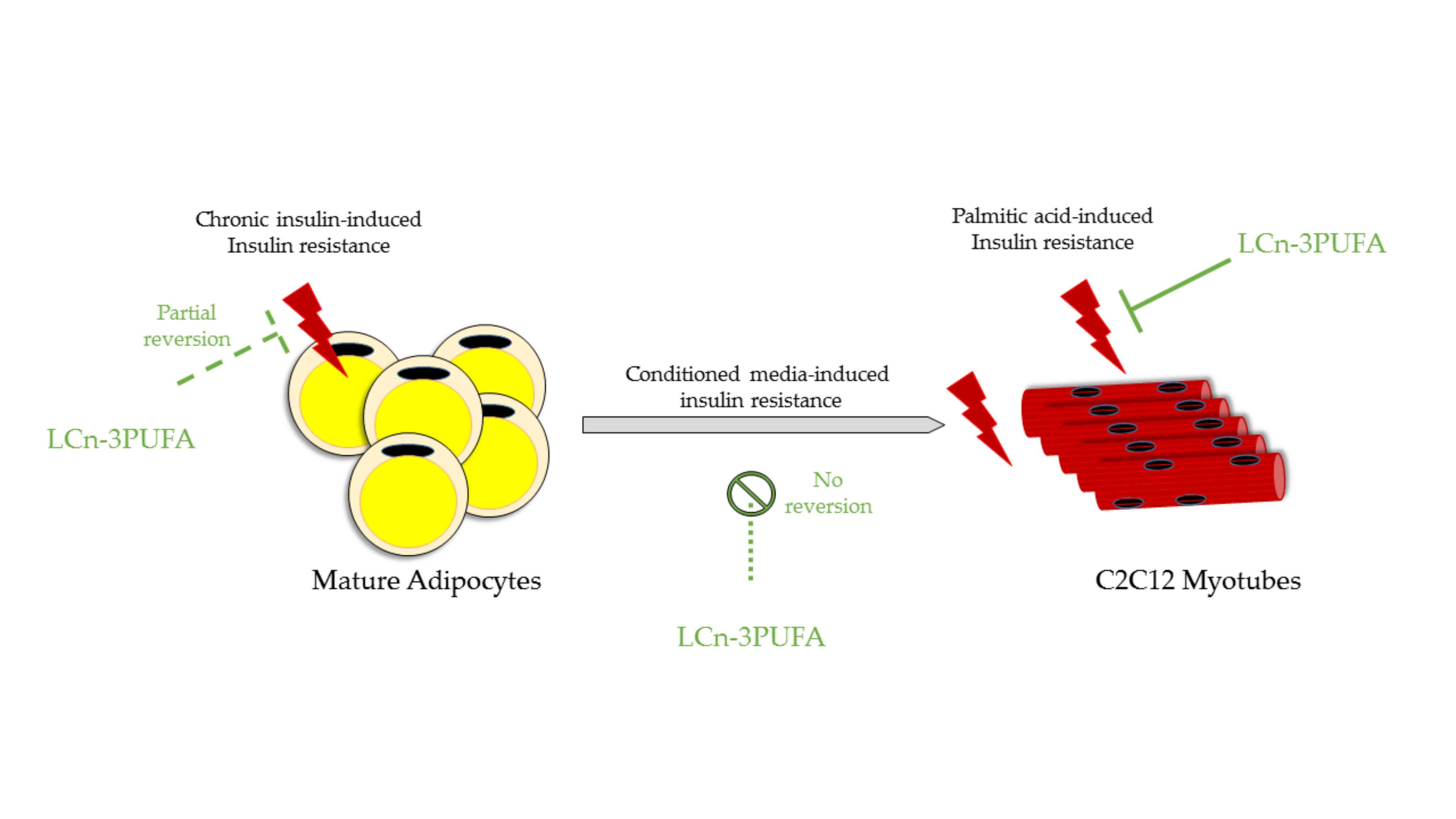
2.2. Effect of Fatty Acid Treatments on Insulin Resistance in C2C12 Muscle Cells
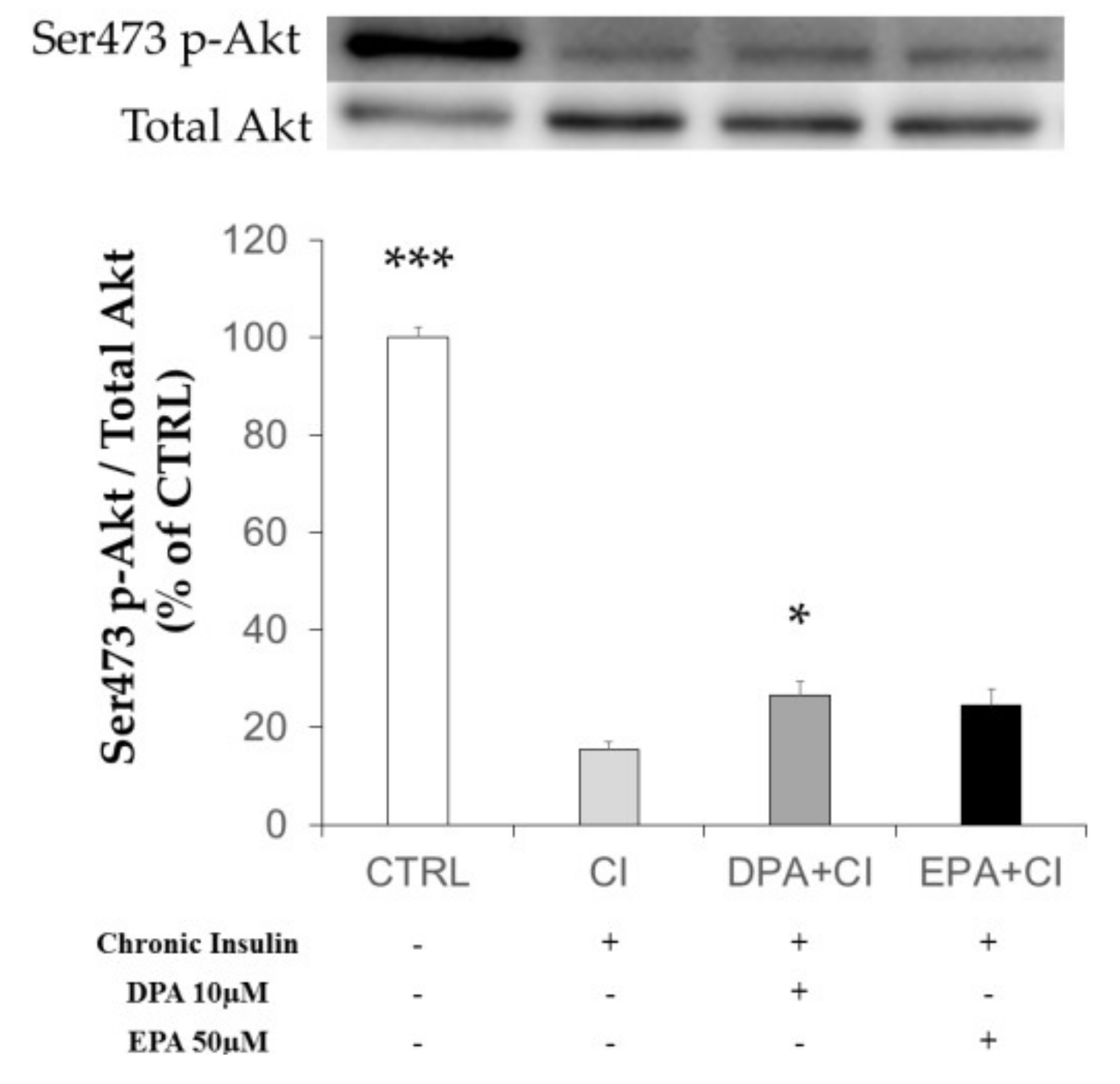
2.3. Chronic Insulin Induced Insulin Resistance in 3T3-L1 Adipocytes
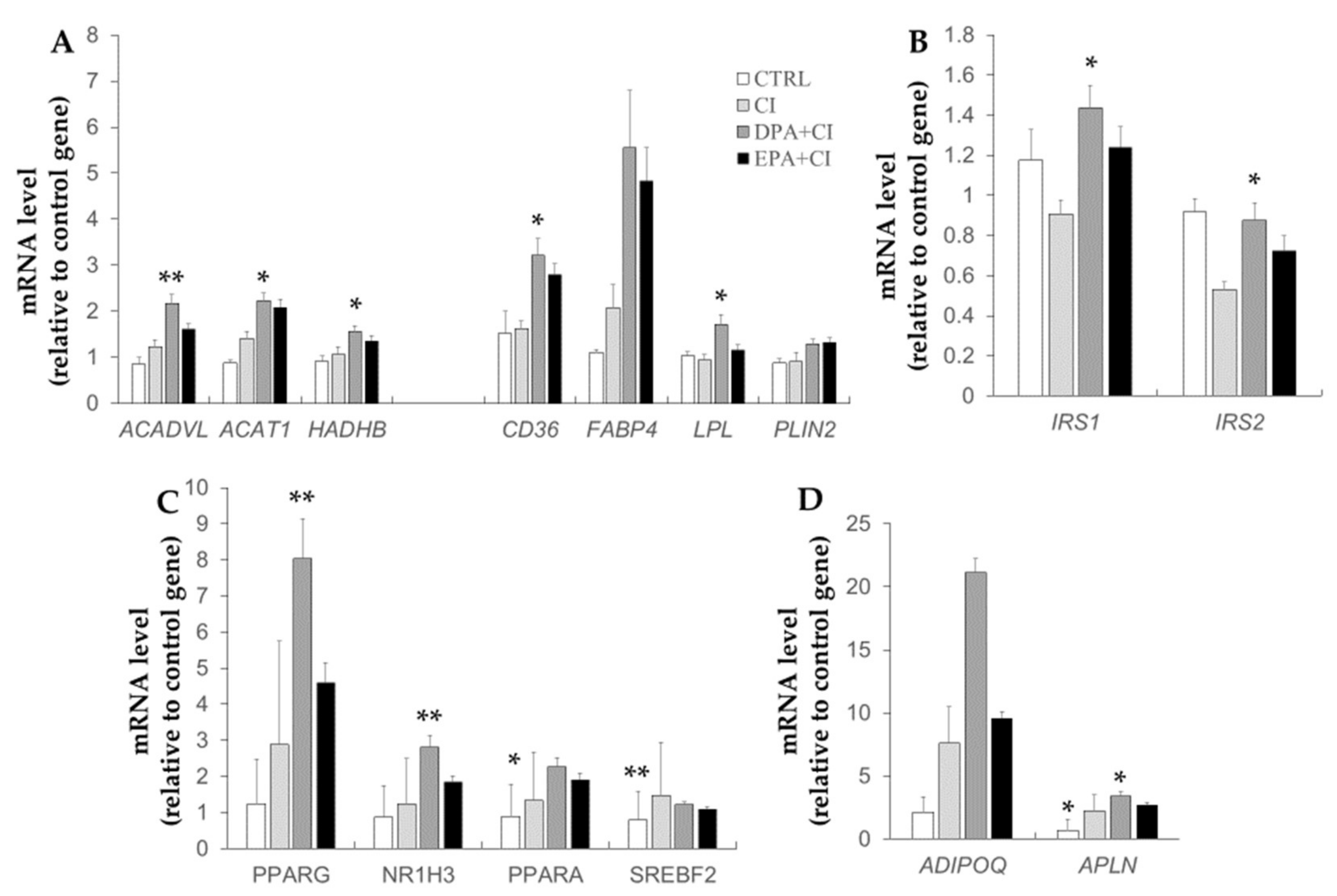
2.4. Effects of Chronic Insulin and LCn-3PUFA Treatments on Gene Expression in 3T3-L1 Adipocytes
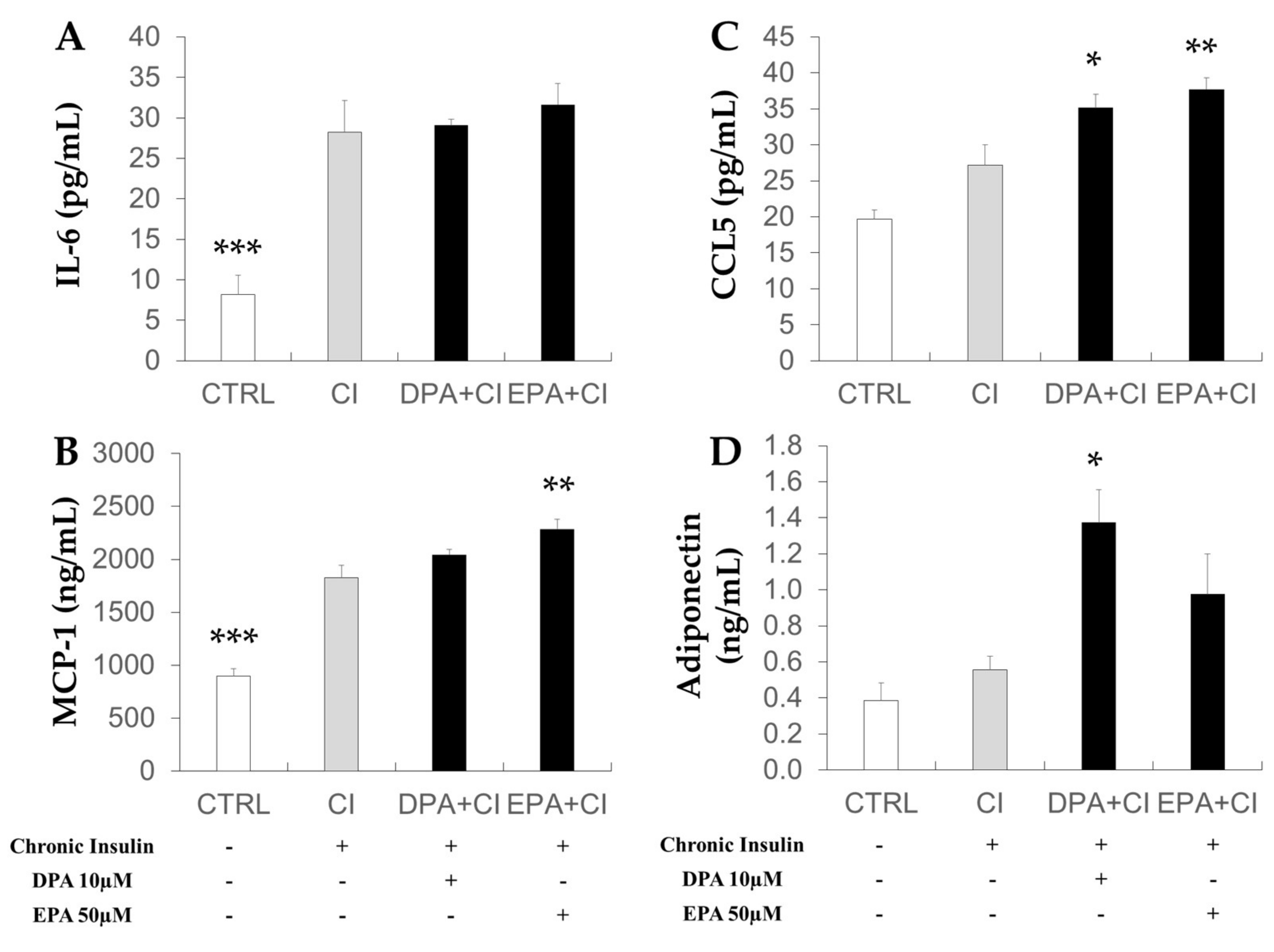
2.5. Effects of Chronic Insulin and LCn-3PUFA Treatments on Mature Adipocyte Secretion
2.6. Effect of Adipocyte-Conditioned Media on Akt Phosphorylation in C2C12 Muscle Cells
3. Discussion
4. Material and Methods
4.1. 3T3-L1 Cell Culture and Treatment.
4.2. Induction of Insulin Resistance in 3T3-L1 Adipocytes
4.3. C2C12 Muscle Cell Culture.
4.4. BSA-Bound PAL Solution for Muscle Cell Treatment with Fatty Acids
4.5. Fatty Acid Treatment
4.6. Treatment of Myotubes with Conditioned Media
4.7. Protein Quantification and Western Blotting
4.8. Reverse Transcriptase Polymerase Chain Reaction
4.9. Phospholipid Extraction
4.10. Phospholipid Separation and Collection by HPLC
4.11. Fatty Acid Profile of PC Fraction
4.12. Quantification of Adipokines and Non-Esterified Fatty Acids in Conditioned Media from Mature Adipocytes
4.13. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AS160 | Akt Substrate of 160 kDa |
| BSA | Bovine Serum Albumin |
| CM | Conditioned Media |
| DG | Diacylglycerol |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DHA | Docosahexaenoic Acid |
| DPA | Docosapentaenoic Acid |
| ECL | Enhanced Chemiluminescence |
| EPA | Eicosapentaenoic Acid |
| EtOH | Ethanol |
| FA | Fatty Acid |
| FBS | Fetal Bovine Serum |
| GAPDH | Glyceraldehyde 3-Phosphate Dehydrogenase |
| HILIC | Hydrophilic Interaction Liquid Chromatography |
| HPLC | High Performance Liquid Chromatography |
| HPRT | Hypoxanthine Guanine Phosphoribosyltransferase |
| IBMX | Isobutyl-Methyl-Xanthine |
| IL-6 | Interleukine-6 |
| IR | Insulin Resistance |
| IRS | Insulin Receptor Substrate |
| LCn-3PUFA | Long Chain n-3 Polyunsaturated Fatty Acids |
| MCP-1 | Monocyte Chemoattractant Protein-1 |
| METS | Metabolic Syndrome |
| PAL | Palmitic acid |
| PC | Phosphatidyl-Choline |
| PKC | Protein Kinase C |
| PL | Phospholipid |
| PPAR | Peroxisome Proliferator-Activated Receptor |
| RNA | Ribonucleic Acid |
References
- Part, P.; Englert, M.; Lambrecht, M.; Taneva, P.; Lazarova, Y. The 2012 Ageing Report: Economic and Budgetary Projections for the EU27 Member States (2010–2060); European Commission: Brussels, Belgium, 2012; Volume 18, pp. 25–36. [Google Scholar]
- Choi, K.M. Sarcopenia and sarcopenic obesity. Endocrinol. Metab. 2016, 31, 1054–1060. [Google Scholar] [Green Version]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–808. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M.; Della Gatta, P.; Suzuki, K.; Nieman, D.C. Cytokine expression and secretion by skeletal muscle cells: regulatory mechanisms and exercise effects. Exerc. Immunol. Rev. 2015, 21, 8–25. [Google Scholar] [PubMed]
- Wei, Y.; Chen, K.; Whaley-Connell, A.T.; Stump, C.S.; Ibdah, J.A.; Sowers, J.R. Skeletal muscle insulin resistance: Role of inflammatory cytokines and reactive oxygen species. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R673–R680. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Liu, B.; Liang, C.; Li, Y.; Song, Y.H. Cytokine signaling in skeletal muscle wasting. Trends Endocrinol. Metab. 2016, 27, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Arner, P.; Ryden, M. Fatty acids, obesity and insulin resistance. Obes. Facts 2015, 8, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Szendroedi, J.; Yoshimura, T.; Phielix, E.; Koliaki, C.; Marcucci, M.; Zhang, D.; Jelenik, T.; Muller, J.; Herder, C.; Nowotny, P.; et al. Role of diacylglycerol activation of PKCtheta in lipid-induced muscle insulin resistance in humans. Proc. Natl. Acad. Sci. USA 2014, 111, 9597–9602. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Keung, W.; Samokhvalov, V.; Wang, W.; Lopaschuk, G.D. Role of fatty acid uptake and fatty acid beta-oxidation in mediating insulin resistance in heart and skeletal muscle. Biochim. Biophys. Acta 2010, 1801, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Pinel, A.; Rigaudiere, J.P.; Laillet, B.; Pouyet, C.; Malpuech-Brugere, C.; Prip-Buus, C.; Morio, B.; Capel, F. N-3PUFA differentially modulate palmitate-induced lipotoxicity through alterations of its metabolism in C2C12 muscle cells. Biochim. Biophys. Acta 2015, 1861, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Bryner, R.W.; Woodworth-Hobbs, M.E.; Williamson, D.L.; Alway, S.E. Docosahexaenoic acid protects muscle cells from palmitate-induced atrophy. ISRN Obes. 2012, 2012, 647348. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Lemaitre, R.N.; King, I.B.; Song, X.; Huang, H.; Sacks, F.M.; Rimm, E.B.; Wang, M.; Siscovick, D.S. Plasma phospholipid long-chain omega-3 fatty acids and total and cause-specific mortality in older adults: A cohort study. Ann. Intern. Med. 2013, 158, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Norris, P.C.; Dennis, E.A. Omega-3 fatty acids cause dramatic changes in TLR4 and purinergic eicosanoid signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 8517–8522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinel, A.; Pitois, E.; Rigaudiere, J.P.; Jouve, C.; de Saint-Vincent, S.; Laillet, B.; Montaurier, C.; Huertas, A.; Morio, B.; Capel, F. EPA prevents fat mass expansion and metabolic disturbances in mice fed with a Western diet. J. Lipid Res. 2016, 57, 1382–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmendorf, J.S. Fluidity of insulin action. Mol. Biotechnol. 2004, 27, 127–138. [Google Scholar] [CrossRef]
- Sakoda, H.; Ogihara, T.; Anai, M.; Funaki, M.; Inukai, K.; Katagiri, H.; Fukushima, Y.; Onishi, Y.; Ono, H.; Fujishiro, M.; et al. Dexamethasone-induced insulin resistance in 3T3-L1 adipocytes is due to inhibition of glucose transport rather than insulin signal transduction. Diabetes 2000, 49, 1700–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, K.; Kawabata, K.; Yamashita, T.; Kawasaki, K.; Yamamoto, N.; Ashida, H. Ameliorative effects of polyunsaturated fatty acids against palmitic acid-induced insulin resistance in L6 skeletal muscle cells. Lipids Health Dis. 2012, 11, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capel, F.; Acquaviva, C.; Pitois, E.; Laillet, B.; Rigaudiere, J.P.; Jouve, C.; Pouyet, C.; Gladine, C.; Comte, B.; Vianey Saban, C.; et al. DHA at nutritional doses restores insulin sensitivity in skeletal muscle by preventing lipotoxicity and inflammation. J. Nutr. Biochem. 2015, 26, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Kozka, I.J.; Clark, A.E.; Holman, G.D. Chronic treatment with insulin selectively down-regulates cell-surface GLUT4 glucose transporters in 3T3-L1 adipocytes. J. Biol. Chem. 1991, 266, 11726–11731. [Google Scholar] [PubMed]
- Kontrova, K.; Zidkova, J.; Bartos, B.; Skop, V.; Sajdok, J.; Kazdova, L.; Mikulik, K.; Mlejnek, P.; Zidek, V.; Pravenec, M. CD36 regulates fatty acid composition and sensitivity to insulin in 3T3-L1 adipocytes. Physiol. Res. 2007, 56, 493–496. [Google Scholar] [PubMed]
- Madsen, L.; Petersen, R.K.; Kristiansen, K. Regulation of adipocyte differentiation and function by polyunsaturated fatty acids. Biochim. Biophys. Acta 2005, 1740, 266–286. [Google Scholar] [CrossRef] [PubMed]
- Miki, H.; Yamauchi, T.; Suzuki, R.; Komeda, K.; Tsuchida, A.; Kubota, N.; Terauchi, Y.; Kamon, J.; Kaburagi, Y.; Matsui, J.; et al. Essential role of insulin receptor substrate 1 (IRS-1) and IRS-2 in adipocyte differentiation. Mol. Cell. Biol. 2001, 21, 2521–2532. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Della-Fera, M.; Lin, J.; Baile, C.A. Docosahexaenoic acid inhibits adipocyte differentiation and induces apoptosis in 3T3-L1 preadipocytes. J. Nutr. 2006, 136, 2965–2969. [Google Scholar] [CrossRef] [PubMed]
- Murali, G.; Desouza, C.V.; Clevenger, M.E.; Ramalingam, R.; Saraswathi, V. Differential effects of eicosapentaenoic acid and docosahexaenoic acid in promoting the differentiation of 3T3-L1 preadipocytes. Leuk. Essent. 2014, 90, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Melloni, E.; Zauli, G.; Celeghini, C.; Volpi, I.; Secchiero, P. Release of a specific set of proinflammatory adipokines by differentiating 3T3-L1 cells. Nutrition 2013, 29, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Prostek, A.; Gajewska, M.; Balasinska, B. The influence of eicosapentaenoic acid and docosahexaenoic acid on expression of genes connected with metabolism and secretory functions of ageing 3T3-L1 adipocytes. Prostag. Lipid Mediat. 2016, 125, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Seyoum, B.; Fite, A.; Abou-Samra, A.B. Effects of 3T3 adipocytes on interleukin-6 expression and insulin signaling in L6 skeletal muscle cells. Biochem. Biophys. Res. Commun. 2011, 410, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Sell, H.; Dietze-Schroeder, D.; Kaiser, U.; Eckel, J. Monocyte chemotactic protein-1 is a potential player in the negative cross-talk between adipose tissue and skeletal muscle. Endocrinology 2006, 147, 2458–2467. [Google Scholar] [CrossRef] [PubMed]
- Esfahani, M.; Movahedian, A.; Baranchi, M.; Goodarzi, M.T. Adiponectin: an adipokine with protective features against metabolic syndrome. Iran. J. Basic Med. Sci. 2015, 18, 430–442. [Google Scholar] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to extracellular vesicles: Biogenesis, RNA cargo selection, content, release, and uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Jalabert, A.; Vial, G.; Guay, C.; Wiklander, O.P.; Nordin, J.Z.; Aswad, H.; Forterre, A.; Meugnier, E.; Pesenti, S.; Regazzi, R.; et al. Exosome-like vesicles released from lipid-induced insulin-resistant muscles modulate gene expression and proliferation of beta recipient cells in mice. Diabetologia 2016, 59, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.; Riopel, M.; Bandyopadhyay, G.; Dong, Y.; Birmingham, A.; Seo, J.B.; Ofrecio, J.M.; Wollam, J.; Hernandez-Carretero, A.; Fu, W.; et al. Adipose tissue macrophage-derived exosomal miRNAs can modulate in vivo and in vitro insulin sensitivity. Cell 2017, 171, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Desvergne, B.; Michalik, L.; Wahli, W. Transcriptional regulation of metabolism. Physiol. Rev. 2006, 86, 465–514. [Google Scholar] [CrossRef] [PubMed]
- Sampath, H.; Ntambi, J.M. Polyunsaturated fatty acid regulation of genes of lipid metabolism. Annu. Rev. Nutr. 2005, 25, 317–340. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandehroo, M.; Knoch, B.; Muller, M.; Kersten, S. Peroxisome proliferator-activated receptor alpha target genes. PPAR Res. 2010, 2010, 612089. [Google Scholar] [CrossRef] [PubMed]
- Dray, C.; Knauf, C.; Daviaud, D.; Waget, A.; Boucher, J.; Buleon, M.; Cani, P.D.; Attane, C.; Guigne, C.; Carpene, C.; et al. Apelin stimulates glucose utilization in normal and obese insulin-resistant mice. Cell Metab. 2008, 8, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Lorente-Cebrian, S.; Bustos, M.; Marti, A.; Martinez, J.A.; Moreno-Aliaga, M.J. Eicosapentaenoic acid up-regulates apelin secretion and gene expression in 3T3-L1 adipocytes. Mol. Nutr. Food Res. 2010, 54, S104–S111. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell types | 3T3-L1 | C2C12 | ||||||
|---|---|---|---|---|---|---|---|---|
| Treatments | CTRL | CI | DPA+CI | EPA+CI | CTRL | PAL | PAL+DPA | PAL+EPA |
| %DPA | 0.69 ± 0.03b | 0.84 ± 0.02b | 2.29 ± 0.54a | 1.10 ± 0.10b | 0.48 ± 0.11b | n.d. | 2.93 ± 0.71a | 1 ± 0.25b |
| %EPA | 4.35 ± 1.30b | 4.74 ± 0.88b | 10.01 ± 2.5b | 24.44 ± 2.87a | 0.94 ± 0.36b | 0.77 ± 0.18b | 1.06 ± 0.19b | 27.13 ± 1.98a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinel, A.; Rigaudière, J.-P.; Jouve, C.; Capel, F. Modulation of Insulin Resistance and the Adipocyte-Skeletal Muscle Cell Cross-Talk by LCn-3PUFA. Int. J. Mol. Sci. 2018, 19, 2778. https://doi.org/10.3390/ijms19092778
Pinel A, Rigaudière J-P, Jouve C, Capel F. Modulation of Insulin Resistance and the Adipocyte-Skeletal Muscle Cell Cross-Talk by LCn-3PUFA. International Journal of Molecular Sciences. 2018; 19(9):2778. https://doi.org/10.3390/ijms19092778
Chicago/Turabian StylePinel, Alexandre, Jean-Paul Rigaudière, Chrystèle Jouve, and Frédéric Capel. 2018. "Modulation of Insulin Resistance and the Adipocyte-Skeletal Muscle Cell Cross-Talk by LCn-3PUFA" International Journal of Molecular Sciences 19, no. 9: 2778. https://doi.org/10.3390/ijms19092778
APA StylePinel, A., Rigaudière, J. -P., Jouve, C., & Capel, F. (2018). Modulation of Insulin Resistance and the Adipocyte-Skeletal Muscle Cell Cross-Talk by LCn-3PUFA. International Journal of Molecular Sciences, 19(9), 2778. https://doi.org/10.3390/ijms19092778