Knockdown of LXRα Inhibits Goat Intramuscular Preadipocyte Differentiation

 ,
,
Abstract
:1. Introduction
2. Results
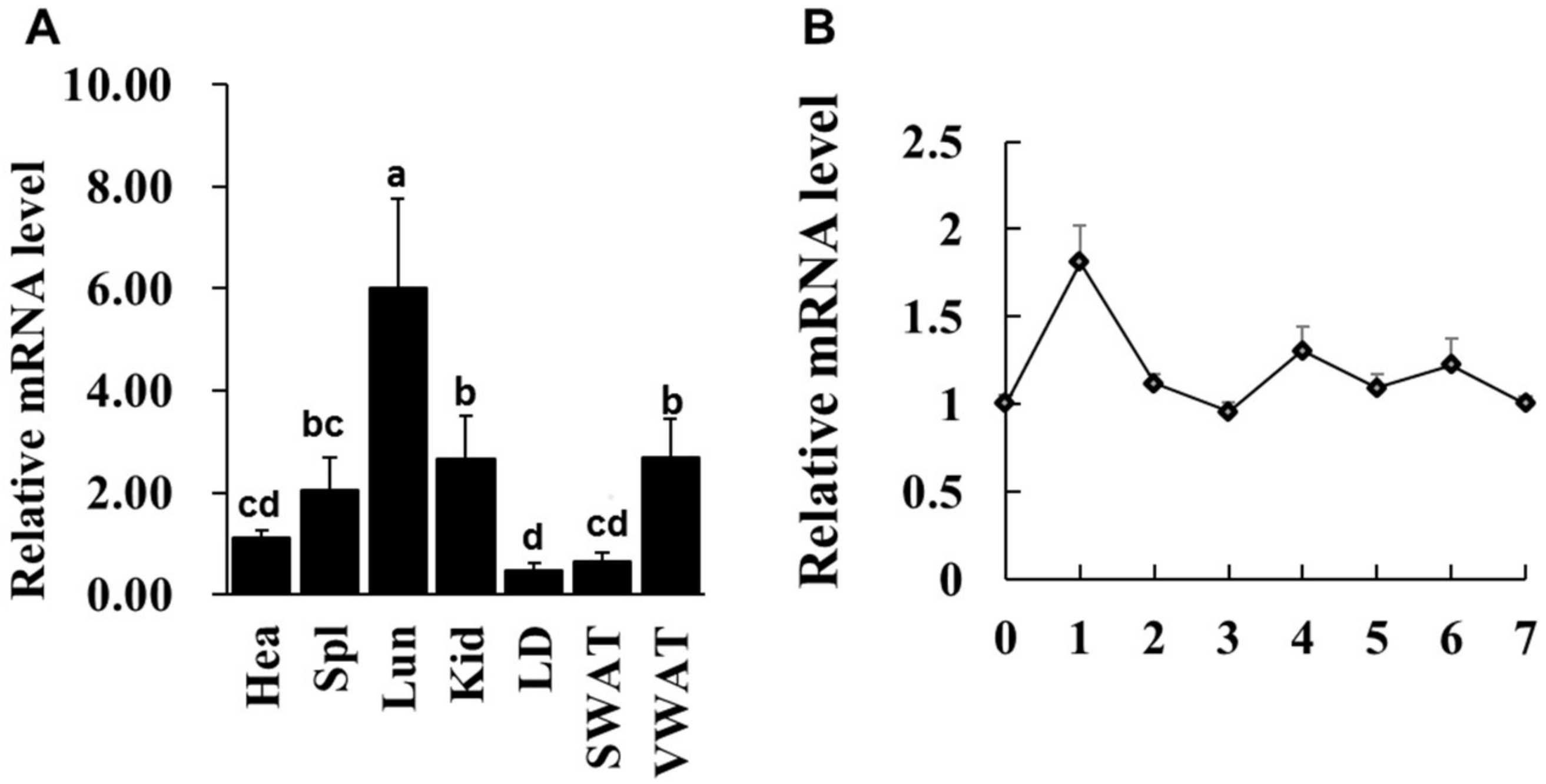
2.1. The Expression Pattern of LXRα in Goat Various Tissues and During Intramuscular Adipocytes Differentiation
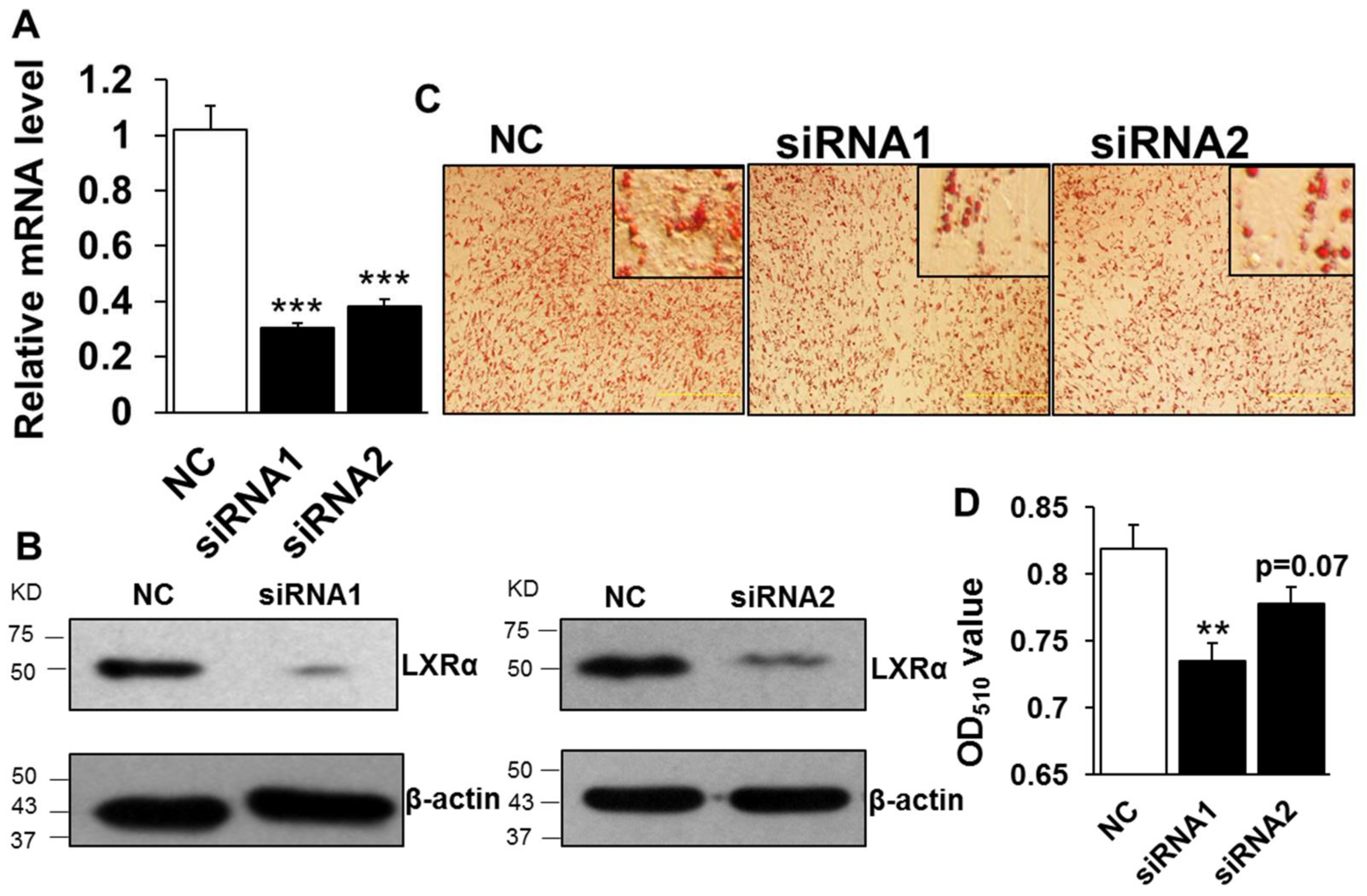
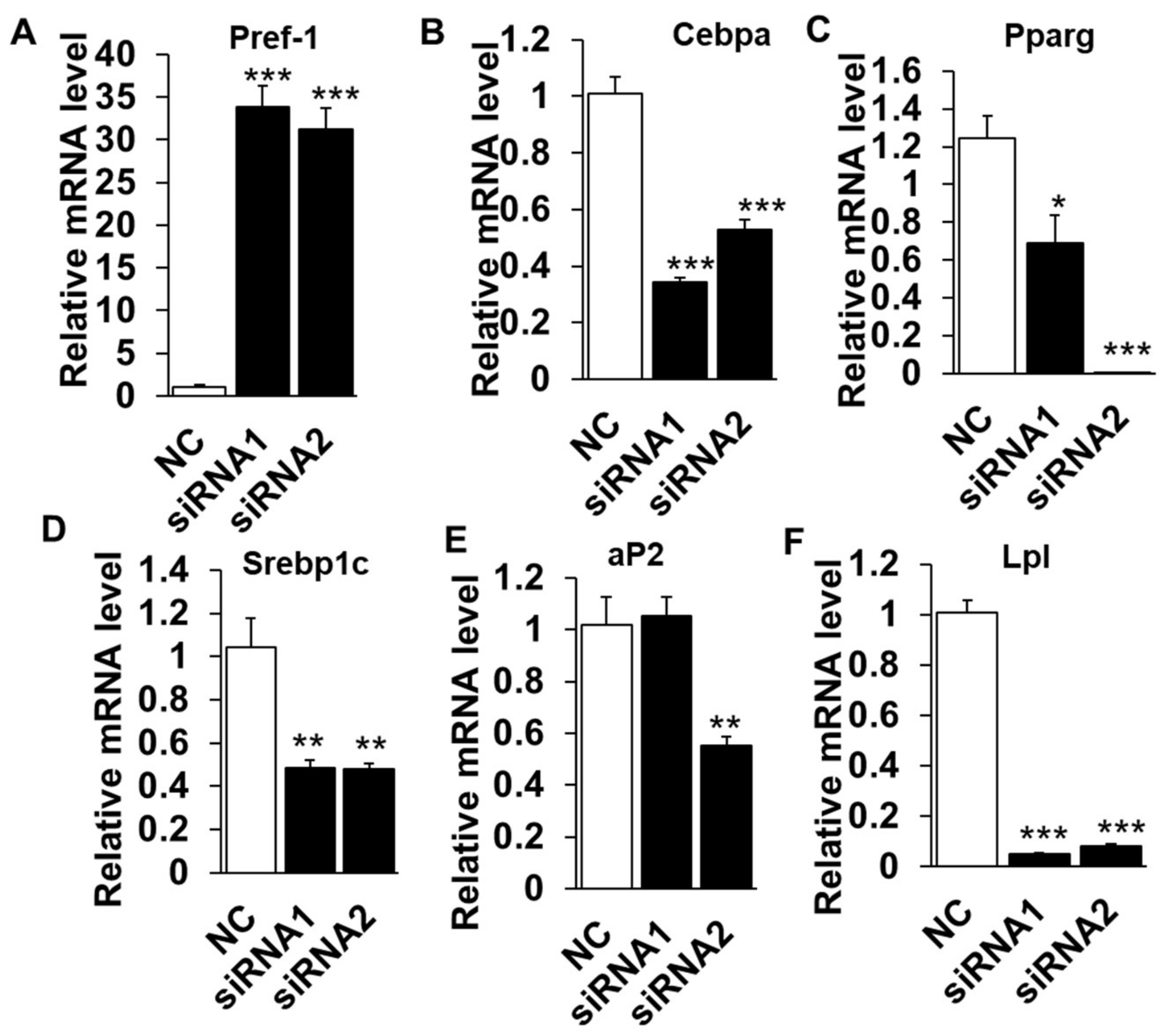
2.2. Loss Function of LXRα Suppresses Intramuscular Adipocytes Differentiation
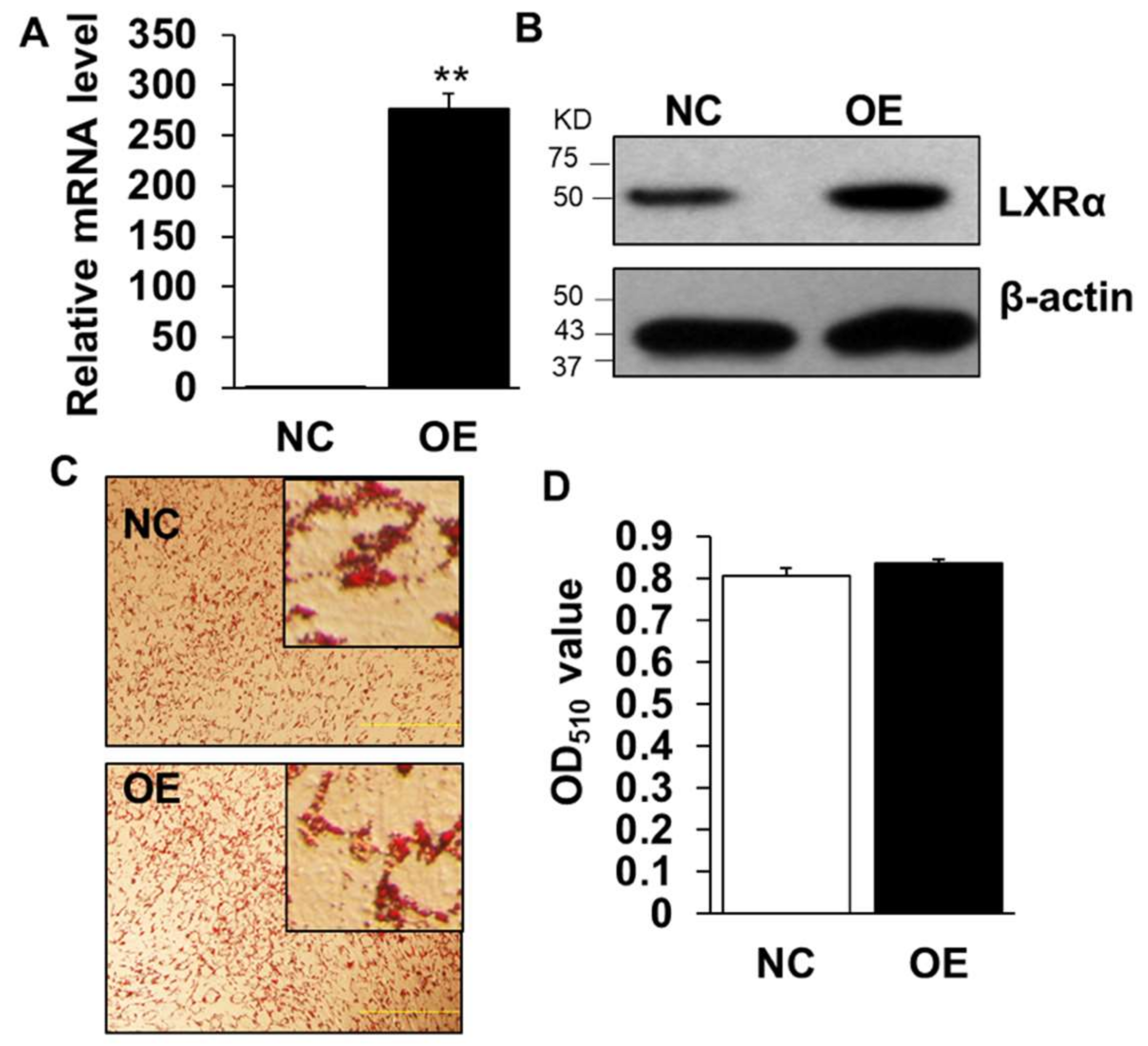
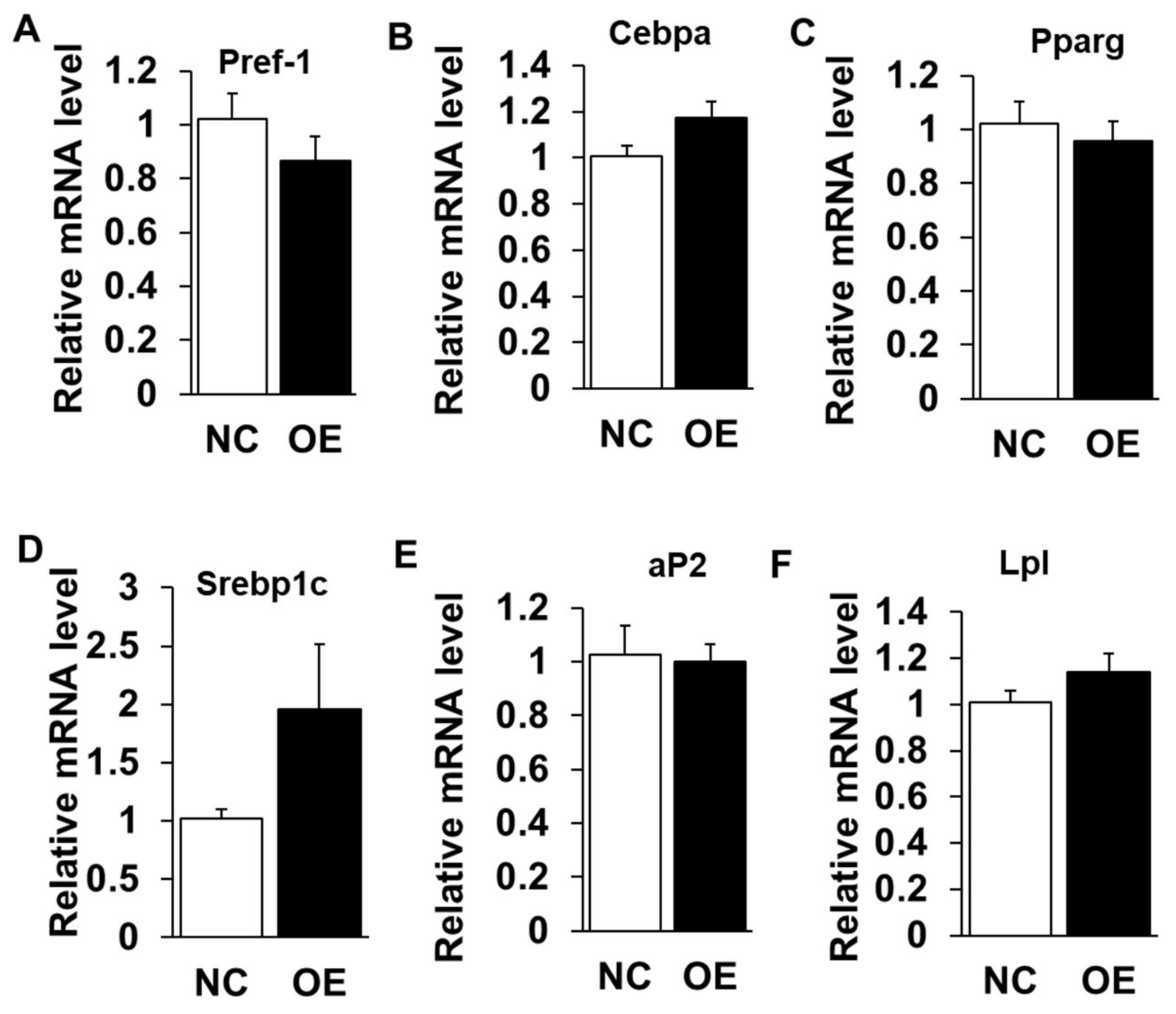
2.3. Overexpression of LXRα Does Not Affect Intramuscular Adipocytes Differentiation
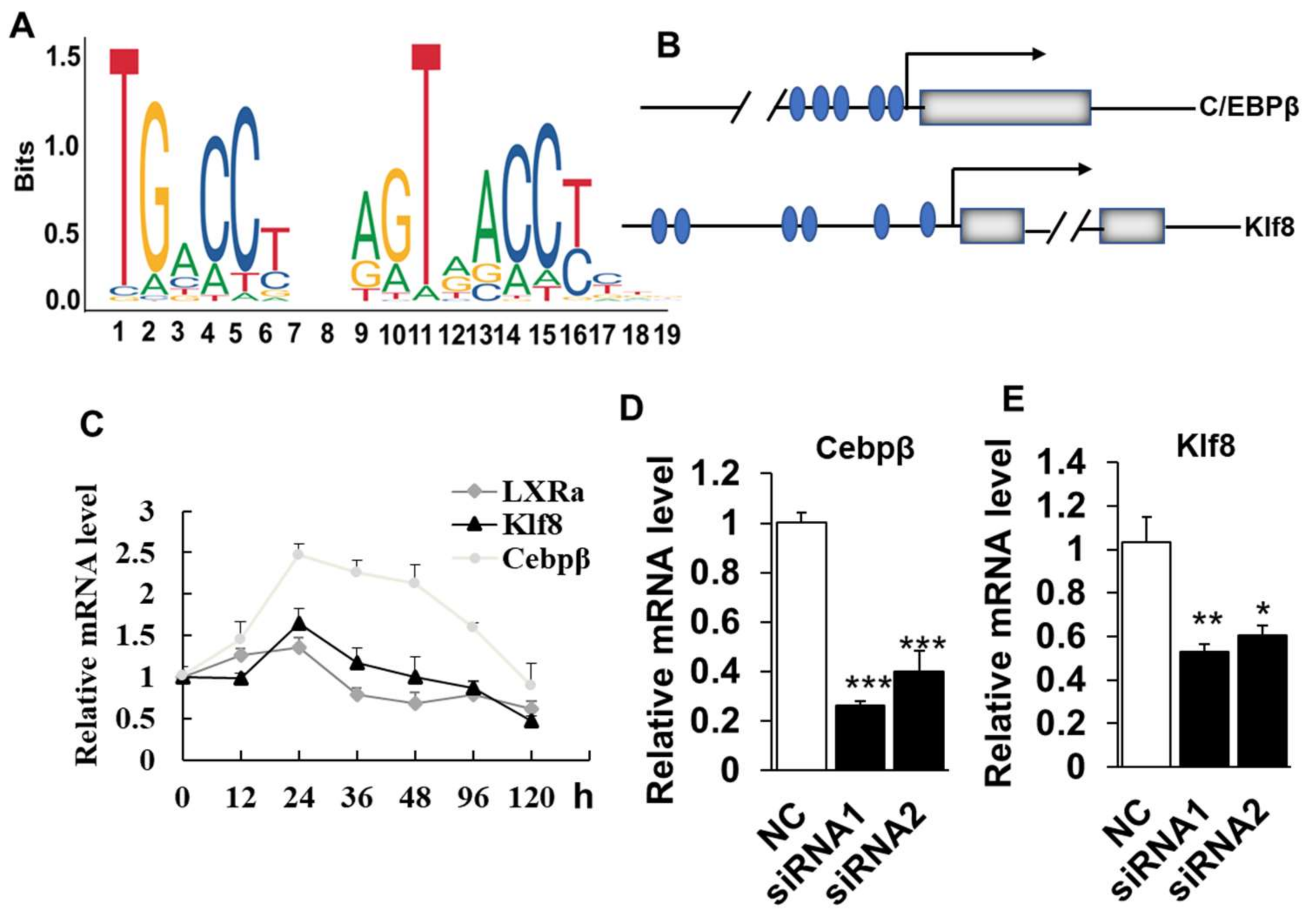
2.4. LXRα Effects on Intramuscular Adipocytes Differentiation through Upregulation of Cebpβ and Klf8
3. Discussion
4. Materials and Methods
4.1. Animal and Cell Culture
4.2. Adenovirus Generation
4.3. Chemical Synthesis of siRNA
4.4. Cell Induction, Transfection and Infection
4.5. Oil Red O Staining
4.6. Total RNA Extraction and Quantitative Real-Time PCR (qPCR)
4.7. Protein Extraction and Western Blot Analysis
4.8. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| IMF | Intramuscular fat |
| LXRα | Liver X receptor α |
| Cebpα | CCAAT enhancer binding protein α |
| Cebpβ | CCAAT enhancer binding protein β |
| Pparg | Peroxisome proliferator activated receptor gamma |
| Srebp1c | Sterol regulatory element binding protein isoform 1c |
| Klf8 | Kruppel like factor 8 |
| aP2 | Fatty acids binding protein 4 |
| Lpl | Lipoprotein lipase |
| SWAT | Subcutaneous white adipose tissue |
| VWAT | Visceral white adipose tissues |
| KO | Knockout |
| WT | Wild type |
| LD | Longissimus dorsi |
| NC | Negative control |
| RXRα | Retinoid X receptor α |
| ApoE | Apolipoprotein E |
References
- Fernandez, X.; Monin, G.; Talmant, A.; Mourot, J.; Lebret, B. Influence of intramuscular fat content on the quality of pig meat-2. Consumer acceptability of m. longissimus lumborum. Meat Sci. 1999, 53, 67–72. [Google Scholar] [CrossRef]
- Cheng, W.W.; Cheng, J.H.; Sun, D.W.; Pu, H.B. Marbling Analysis for Evaluating Meat Quality: Methods and Techniques. Compr. Rev. Food Sci. Food Saf. 2015, 14, 523–535. [Google Scholar] [CrossRef] [Green Version]
- Addison, O.; Marcus, R.L.; LaStayo, P.C.; Ryan, A.S. Intermuscular Fat: A Review of the Consequences and Causes. Int. J. Endocrinol. 2014, 2014, 309570. [Google Scholar] [CrossRef] [PubMed]
- Hocquette, J.F.; Gondret, F.; Baeza, E.; Medale, F.; Jurie, C.; Pethick, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Lowe, C.E.; O’Rahilly, S.; Rochford, J.J. Adipogenesis at a glance. J. Cell Sci. 2011, 124, 2681–2686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.H.; Lu, J.X.; Chen, Y.; Zhao, Y.Q.; Guo, P.H.; Yang, J.T.; Zang, R.X. Comparison of the adipogenesis in intramuscular and subcutaneous adipocytes from Bamei and Landrace pigs. Biochem. Cell Biol. 2014, 92, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.X.; Wang, H.H.; Jiang, B.C.; Zhao, Y.Y.; Xie, Z.R.; Xiong, K.; Chen, J. Global comparison of gene expression between subcutaneous and intramuscular adipose tissue of mature Erhualian pig. Genet. Mol. Res. 2013, 12, 5085–5101. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Fu, S.Y.; Gao, H.Y.; Zheng, Z.Q.; Meng, X.R.; A, N.; Sa, R.; Zhang, W.G.; Li, J.Q. Transcriptome Analysis of Intramuscular Preadipocytes and Matureadipocyte in Cashmere Goats. Acta Veterinaria et Zootechnica Sinica 2014, 45, 714–721. [Google Scholar]
- Sheng, X.H.; Ni, H.M.; Liu, Y.H.; Li, J.Y.; Zhang, L.P.; Guo, Y. RNA-seq analysis of bovine intramuscular, subcutaneous and perirenal adipose tissues. Mol. Biol. Rep. 2014, 41, 1631–1637. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.X.; Mo, D.L.; Zhang, Y.; Zhang, Y.; Cong, P.Q.; Xiao, S.Q.; He, Z.Y.; Liu, X.H.; Chen, Y.S. MicroRNAome Comparison between Intramuscular and Subcutaneous Vascular Stem Cell Adipogenesis. PLOS ONE 2012, 7, e45410. [Google Scholar] [CrossRef] [PubMed]
- Schulman, I.G. Liver X receptors link lipid metabolism and inflammation. FEBS Lett. 2017, 591, 2978–2991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korach-Andre, M.; Gustafsson, J.A. Liver X receptors as regulators of metabolism. Biomol. Concepts 2015, 6, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Tontonoz, P. Liver X receptors in lipid metabolism: opportunities for drug discovery. Nat. Rev. Drug Discov. 2014, 13, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Archer, A.; Laurencikiene, J.; Ahmed, O.; Steffensen, K.R.; Parini, P.; Gustafsson, J.A.; Korach-Andre, M. Skeletal muscle as a target of LXR agonist after long-term treatment: focus on lipid homeostasis. Am. J. Physiol.-Endoc. Metab. 2014, 306, E494–E502. [Google Scholar] [CrossRef] [PubMed]
- Dib, L.; Bugge, A.; Collins, S. LXRα fuels fatty acid-stimulated oxygen consumption in white adipocytes. J. Lipid Res. 2014, 55, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Ulven, S.M.; Dalen, K.T.; Gustafsson, J.A.; Nebb, H.I. LXR is crucial in lipid metabolism. Prostag. Leukot. Ess. 2005, 73, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Stenson, B.M.; Ryden, M.; Venteclef, N.; Dahlman, I.; Pettersson, A.M.; Mairal, A.; Astrom, G.; Blomqvist, L.; Wang, V.; Jocken, J.W.; et al. Liver X receptor (LXR) regulates human adipocyte lipolysis. J. Biol. Chem. 2011, 286, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Spiegelman, B.M. Cell biology of fat storage. Mol. Biol. Cell 2016, 27, 2523–2527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Fornes, O.; Stigliani, A.; Gheorghe, M.; Castro-Mondragon, J.A.; van der Lee, R.; Bessy, A.; Cheneby, J.; Kulkarni, S.R.; Tan, G.; et al. JASPAR 2018: update of the open-access database of transcription factor binding profiles and its web framework. Nucleic Acids Res 2018, 46, D1284. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.B.; Miao, Y.F.; Wu, W.F.; Li, Y.; D’Errico, F.; Su, W.; Burns, A.R.; Huang, B.; Maneix, L.; Warner, M.; et al. Ablation of Liver X receptors alpha and beta leads to spontaneous peripheral squamous cell lung cancer in mice. Proc. Natl. Acad. Sci. USA 2016, 113, 7614–7619. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M. Subcutaneous and visceral adipose tissue: structural and functional differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Albrecht, E.; Ender, K.; Zhao, R.Q.; Wegner, J. Computer image analysis of intramuscular adipocytes and marbling in the longissimus muscle of cattle. J. Anim. Sci. 2006, 84, 3251–3258. [Google Scholar] [CrossRef] [PubMed]
- Hausman, G.J.; Basu, U.; Du, M.; Fernyhough-Culver, M.; Dodson, M.V. Intermuscular and intramuscular adipose tissues: Bad vs. good adipose tissues. Adipocyte 2014, 3, 242–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldai, N.; Najera, A.I.; Dugan, M.E.; Celaya, R.; Osoro, K. Characterisation of intramuscular, intermuscular and subcutaneous adipose tissues in yearling bulls of different genetic groups. Meat. Sci. 2007, 76, 682–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahnalampi, M.; Heinäniemi, M.; Sinkkonen, L.; Wabitsch, M.; Carlberg, C. Time-resolved expression profiling of the nuclear receptor superfamily in human adipogenesis. PLoS ONE 2010, 5, e12991. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Kim, K.A.; Kim, J.H.; Sul, H.S. Pref-1, a preadipocyte secreted factor that inhibits adipogenesis. J. Nutr. 2006, 136, 2953–2956. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Muise, A.M.; Lyons, P.J.; Ro, H.S. Regulation of adipogenesis by a transcriptional repressor that modulates MAPK activation. J. Biol. Chem. 2001, 276, 10199–10206. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Puigserver, P.; Spiegelman, B.M. Transcriptional activation of adipogenesis. Curr. Opin. Cell Biol. 1999, 11, 689–694. [Google Scholar] [CrossRef]
- Matsushita, K.; Morello, F.; Zhang, Z.; Masuda, T.; Iwanaga, S.; Steffensen, K.R.; Gustafsson, J.Å.; Pratt, R.E.; Dzau, V.J. Nuclear hormone receptor LXRα inhibits adipocyte differentiation of mesenchymal stem cells with Wnt/beta-catenin signaling. Lab. Invest. 2016, 96, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.B.; Moon, H.M.; Kim, W.S.; Lee, Y.S.; Jeong, H.W.; Yoo, E.J.; Ham, J.; Kang, H.; Park, M.G.; Steffensen, K.R.; et al. Activated liver X receptors stimulate adipocyte differentiation through induction of peroxisome proliferator-activated receptor gamma expression. Mol. Cell Biol. 2004, 24, 3430–3444. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Ye, F.; Gui, C.; Luo, H.; Cai, J.; Shen, J.; Chen, K.; Shen, X.; Jiang, H. Ligand-binding regulation of LXR/RXR and LXR/PPAR heterodimerizations: SPR technology-based kinetic analysis correlated with molecular dynamics simulation. Protein Sci. 2005, 14, 812–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willy, P.J.; Mangelsdorf, D.J. Unique requirements for retinoid-dependent transcriptional activation by the orphan receptor LXR. Genes Dev. 1997, 11, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Laffitte, B.A.; Repa, J.J.; Joseph, S.B.; Wilpitz, D.C.; Kast, H.R.; Mangelsdorf, D.J.; Tontonoz, P. LXRs control lipid-inducible expression of the apolipoprotein E gene in macrophages and adipocytes. Proc. Natl. Acad. Sci. USA 2001, 98, 507–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Repa, J.J.; Liang, G.; Ou, J.; Bashmakov, Y.; Lobaccaro, J.M.; Shimomura, I.; Shan, B.; Brown, M.S.; Goldstein, J.L.; Mangelsdorf, D.J. Regulation of mouse sterol regulatory element-binding protein-1c gene (SREBP-1c) by oxysterol receptors, LXRα and LXRβ. Genes Dev. 2000, 14, 2819–2830. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, R.; Lin, Y. Allograft inflammatory factor-1 in grass carp (Ctenopharynogodon idella): Expression and response to cadmium exposure. Fish Shellfish Immunol. 2015, 47, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.F.; Xiong, Y.; Peng, Y.; Gao, Y.; Qin, J.; Chu, G.Y.; Pang, W.J.; Yang, G.S. miR-425-5p Inhibits Differentiation and Proliferation in Porcine Intramuscular Preadipocytes. Int. J. Mol. Sci. 2017, 18, 2101. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Yue, F.; Jia, Z.; Gao, Y.; Jin, W.; Hu, K.; Zhang, Y.; Zhu, D.; Yang, G.; Kuang, S. A novel brown adipocyte-enriched long non-coding RNA that is required for brown adipocyte differentiation and sufficient to drive thermogenic gene program in white adipocytes. Biochim. Biophys. Acta 2018, 1863, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Guo, L.; Wang, N.; Shi, H.; Wang, Y.; Li, H. Oleate promotes differentiation of chicken primary preadipocytes in vitro. Bioscience rep. 2014, 34, e00093. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5’-3’) | Sequence (5’-3’) | GenBank |
|---|---|---|---|
| Cebpα | CCGTGGACAAGAACAGCAAC | AGGCGGTCATTGTCACTGGT | XM_018062278.1 |
| LXRα | TCGGAGGTACAACCCTGGAA | ATGGCAATGAGCAAGACAAACT | NM_001285751.1 |
| Cebpβ | CAAGAAGACGGTGGACAAGC | AACAAGTTCCGCAGGGTG | XM_018058020.1 |
| Srebp1c | AAGTGGTGGGCCTCTCTGA | GCAGGGGTTTCTCGGACT | NM_001285755.1 |
| Klf8 | GACTACAGCAAGAACCAGCAGC | CTCCTGTATGGATTCTGCGGT | KX247671 |
| aP2 | TGAAGTCACTCCAGATGACAGG | TGACACATTCCAGCACCAGC | NM_001285623.1 |
| Lpl | TCCTGGAGTGACGGAATCTGT | GACAGCCAGTCCACCACGAT | NM_001285607.1 |
| Pparg | AAGCGTCAGGGTTCCACTATG | GAACCTGATGGCGTTATGAGAC | NM_001285658.1 |
| Pref1 | CCGGCTTCATGGATAAGACCT | GCCTCGCACTTGTTGAGGAA | KP686197.1 |
| Ppia | ACAAAGTCCCGAAGACAGCAG | AAGTCACCACCCTGGCACAT | XM_005679322.2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Y.; Xu, Q.; Lin, S.; Wang, Y.; Lin, Y.; Zhu, J. Knockdown of LXRα Inhibits Goat Intramuscular Preadipocyte Differentiation. Int. J. Mol. Sci. 2018, 19, 3037. https://doi.org/10.3390/ijms19103037
Xiong Y, Xu Q, Lin S, Wang Y, Lin Y, Zhu J. Knockdown of LXRα Inhibits Goat Intramuscular Preadipocyte Differentiation. International Journal of Molecular Sciences. 2018; 19(10):3037. https://doi.org/10.3390/ijms19103037
Chicago/Turabian StyleXiong, Yan, Qing Xu, Sen Lin, Yong Wang, Yaqiu Lin, and Jiangjiang Zhu. 2018. "Knockdown of LXRα Inhibits Goat Intramuscular Preadipocyte Differentiation" International Journal of Molecular Sciences 19, no. 10: 3037. https://doi.org/10.3390/ijms19103037
APA StyleXiong, Y., Xu, Q., Lin, S., Wang, Y., Lin, Y., & Zhu, J. (2018). Knockdown of LXRα Inhibits Goat Intramuscular Preadipocyte Differentiation. International Journal of Molecular Sciences, 19(10), 3037. https://doi.org/10.3390/ijms19103037