AWRK6, a Novel GLP-1 Receptor Agonist, Attenuates Diabetes by Stimulating Insulin Secretion



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
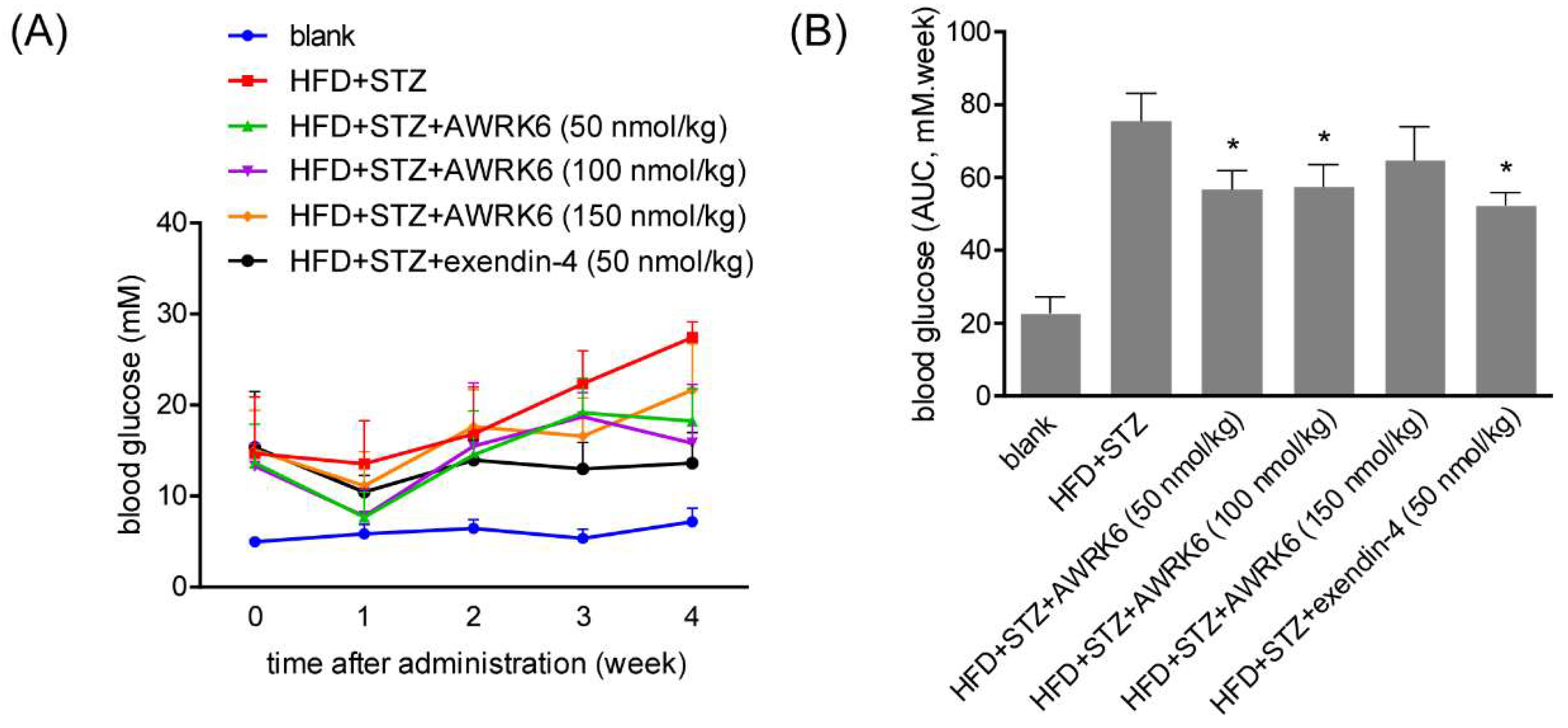
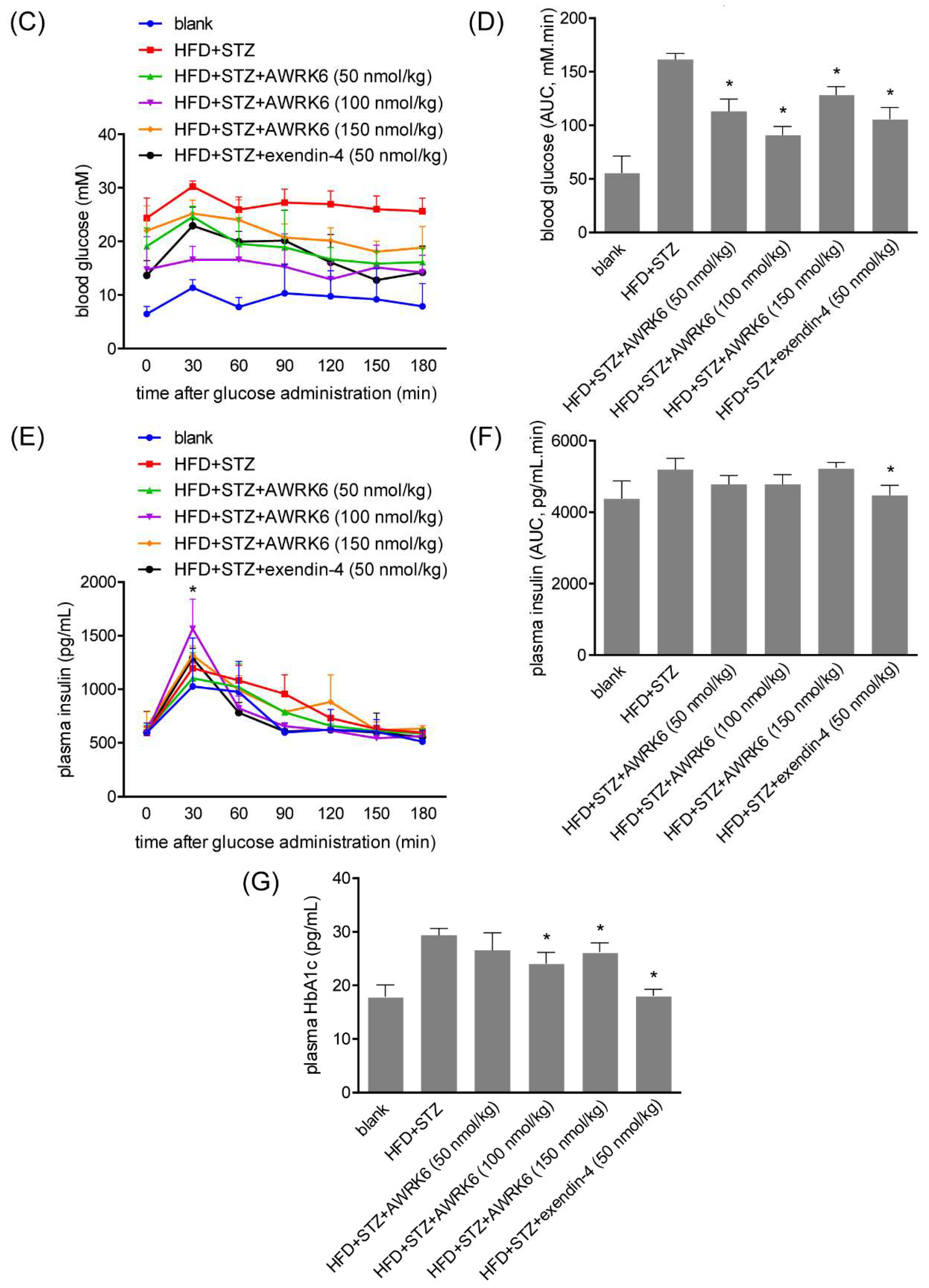
2.1. AWRK6 Reduced Blood Glucose in Diabetic Mice
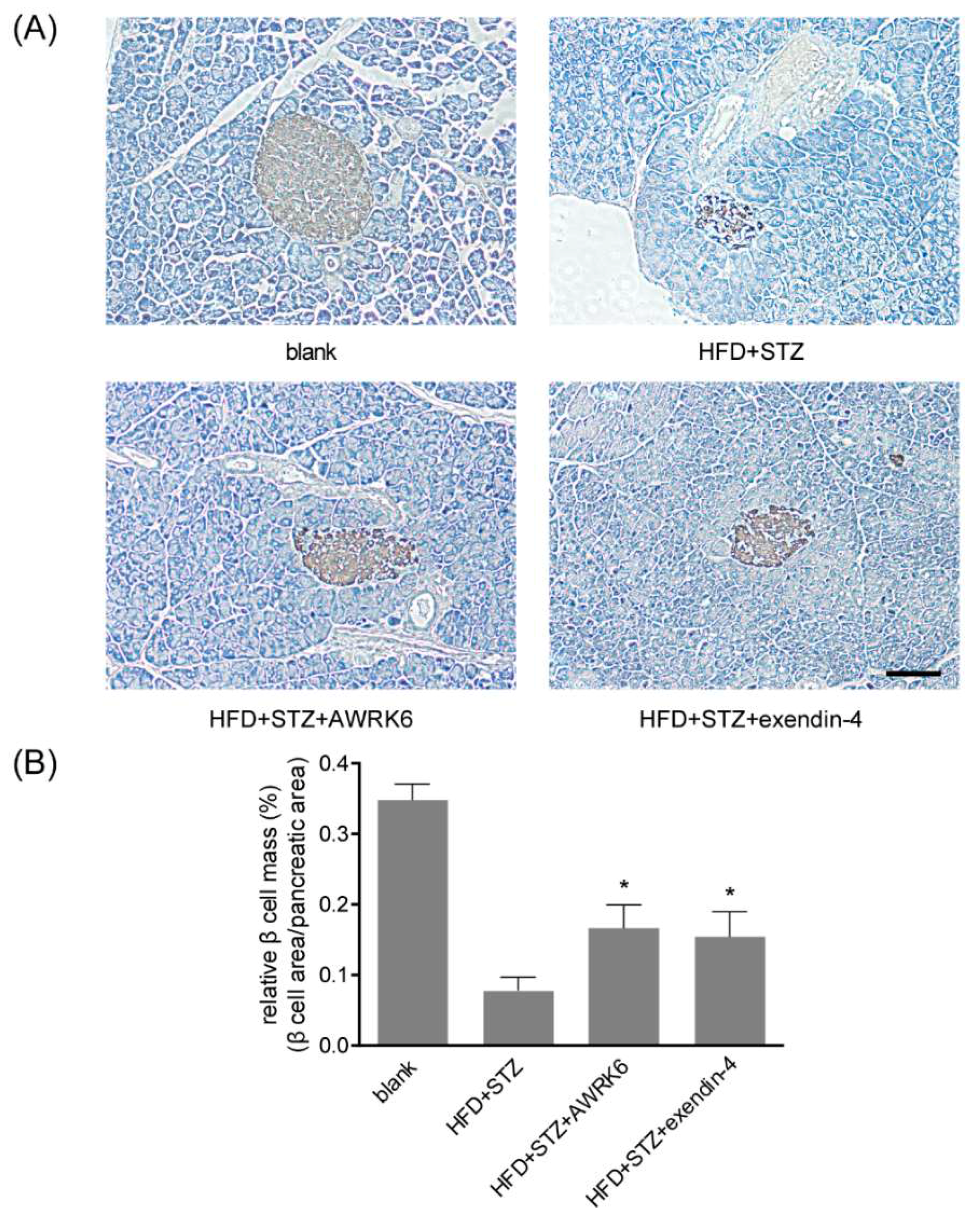
2.2. AWRK6 Increased β Cell Mass in Diabetic Mice
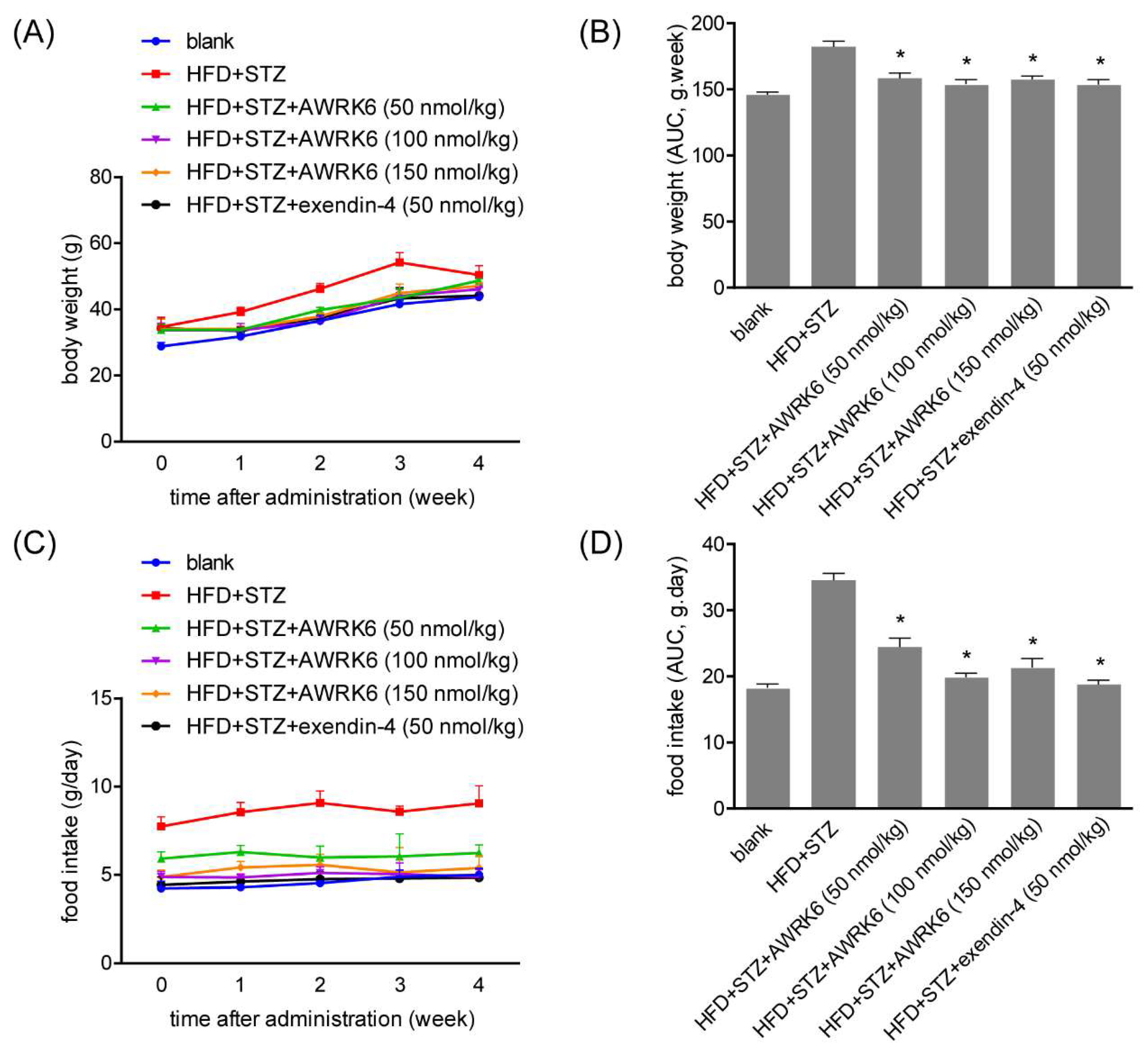
2.3. AWRK6 Decreased Food Intake and Body Weight
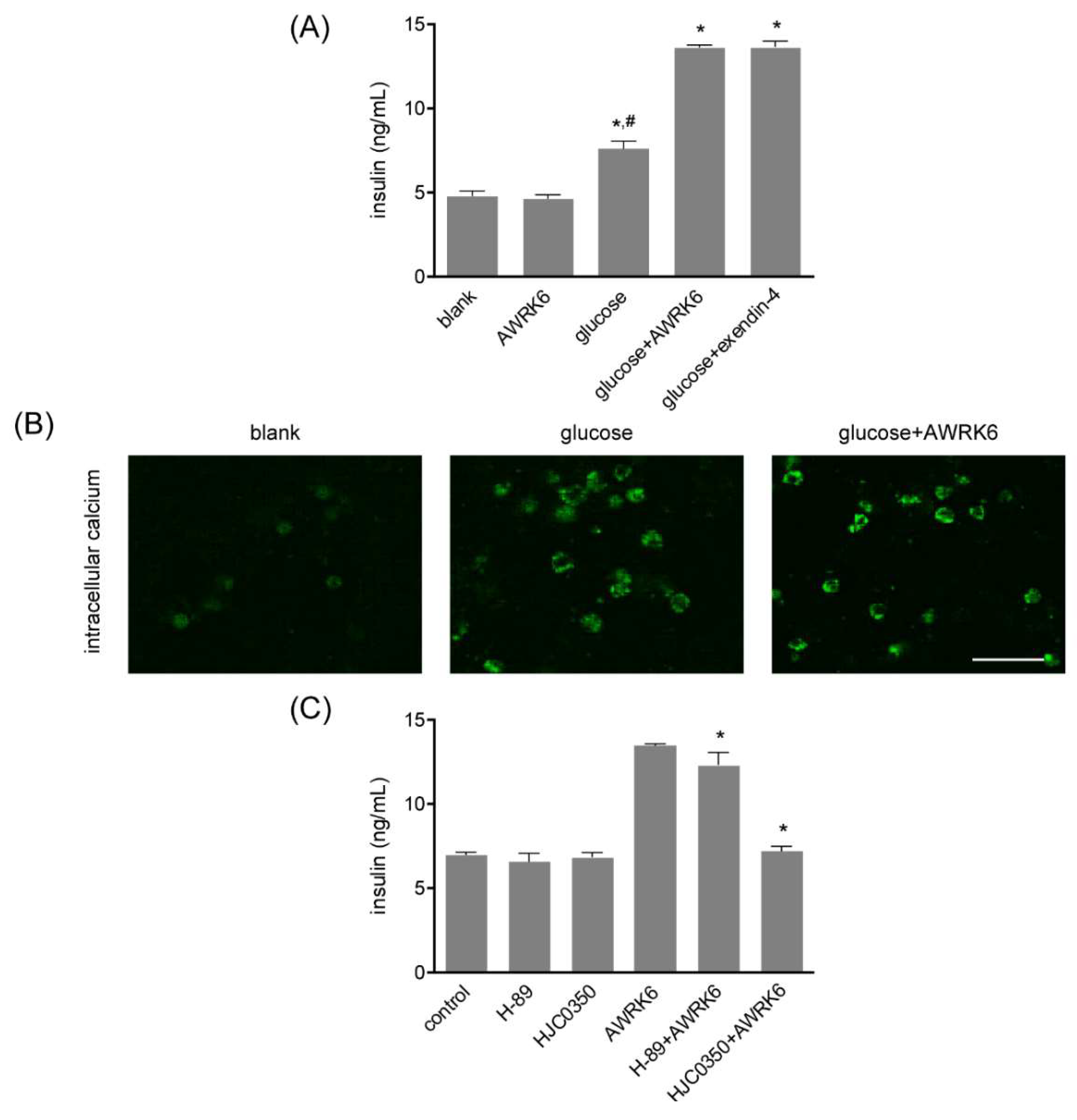
2.4. AWRK6 Induced Insulin in a cAMP-Dependent Manner
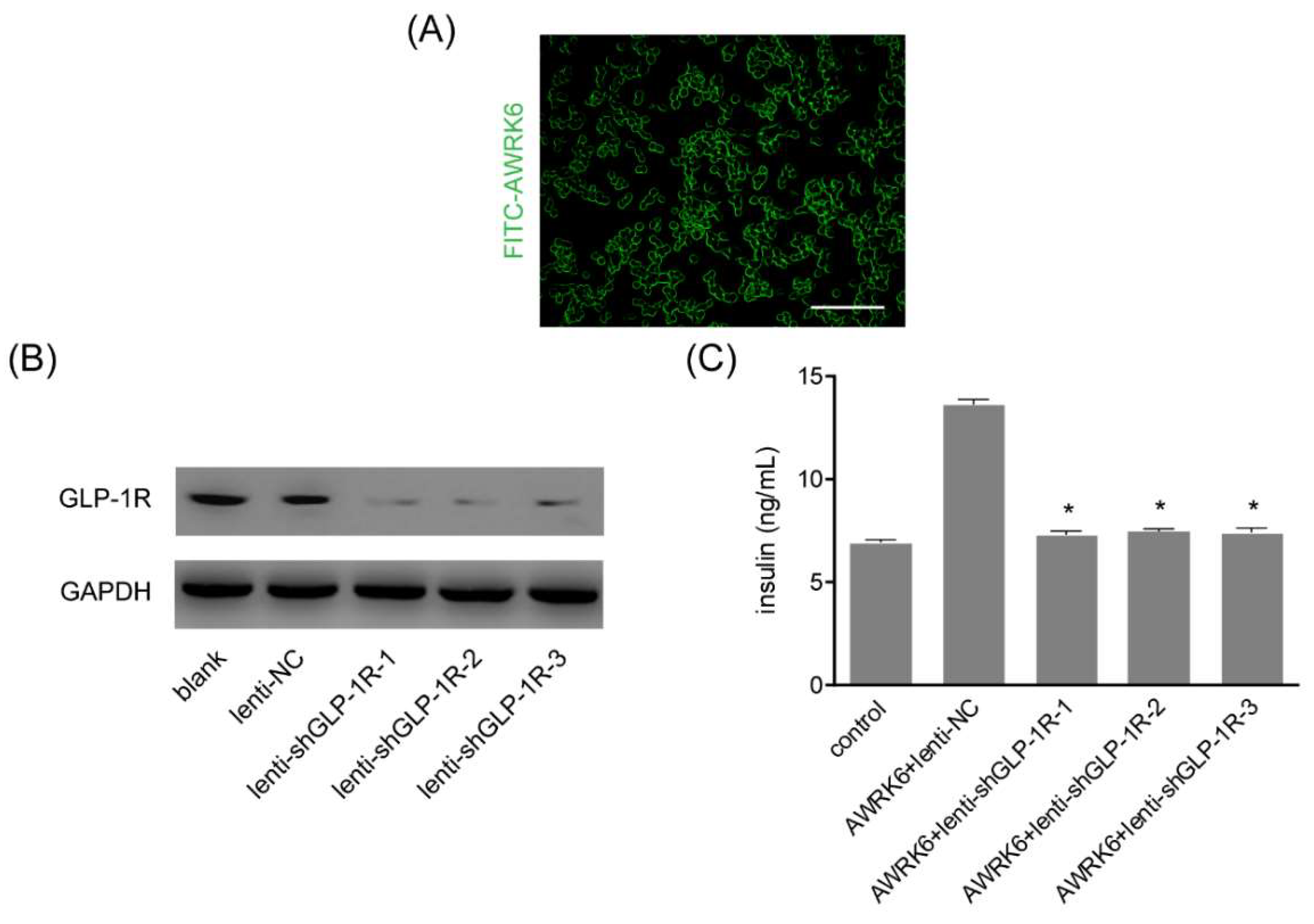
2.5. AWRK6 Was a GLP-1R Agonist
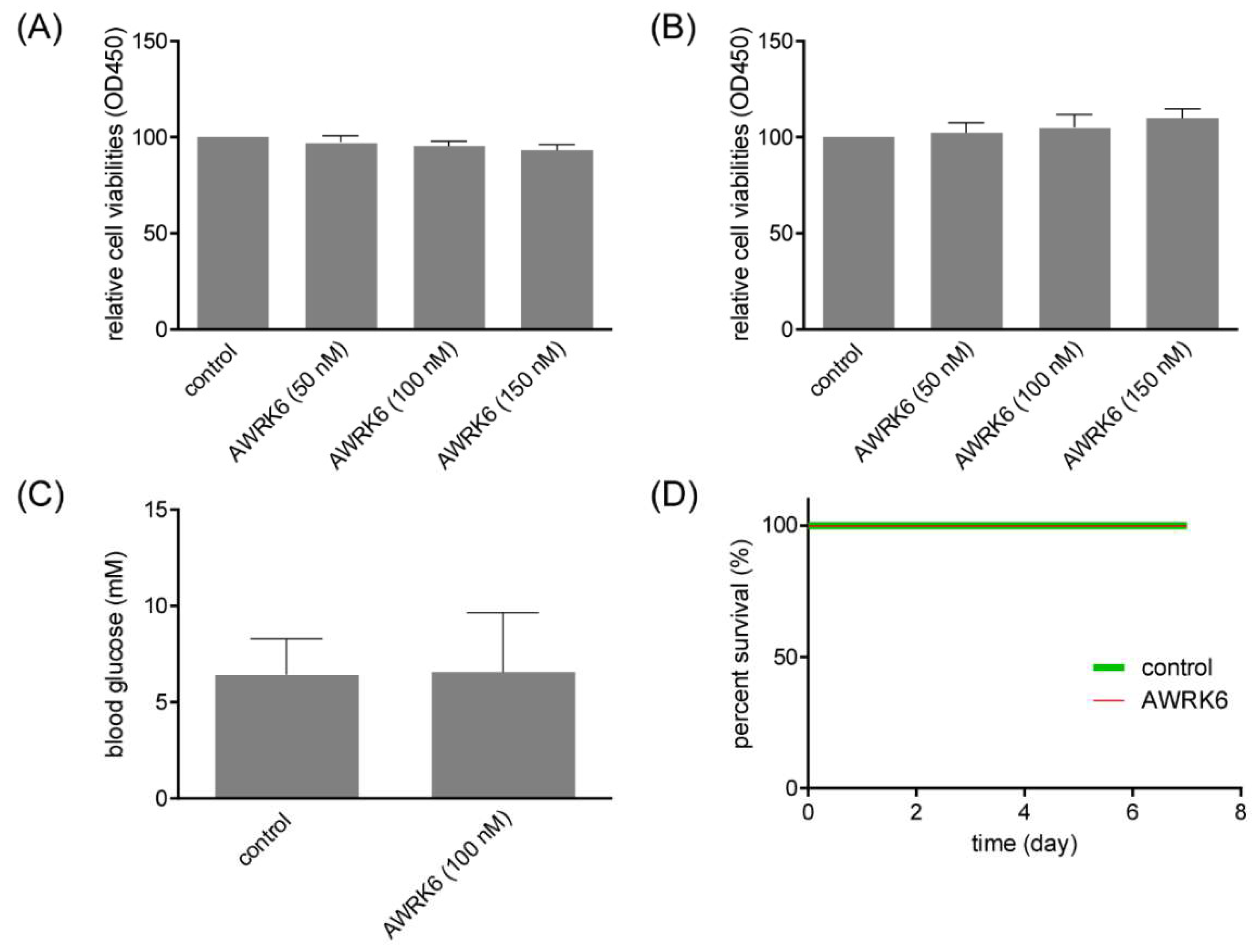
2.6. AWRK6 Showed No Toxicity in Vitro and in Vivo
3. Discussion
4. Materials and Methods
4.1. Peptides
4.2. Mice Model
4.3. Blood Glucose, Insulin, and HbA1c Measurement
4.4. Histopathological Examination
4.5. Cell Culture
4.6. Intracellular Calcium Determination
4.7. GLP-1R Silencing by Lentivirus
4.8. Western Blotting
4.9. Cell Viability Assay
4.10. LDH Release Assay
4.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Puchulu, F.M. Definition, Diagnosis and Classification of Diabetes Mellitus. In Dermatology and Diabetes; Cohen Sabban, E.N., Puchulu, F.M., Cusi, K., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 7–18. [Google Scholar]
- Zaccardi, F.; Webb, D.R.; Yates, T.; Davies, M.J. Pathophysiology of type 1 and type 2 diabetes mellitus: A 90-year perspective. Postgrad. Med. J. 2016, 92, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Hu, F.B. The global implications of diabetes and cancer. Lancet 2014, 383, 1947–1948. [Google Scholar] [CrossRef]
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Datta-Nemdharry, P.; Thomson, A.; Beynon, J.; Donegan, K. Patterns of anti-diabetic medication use in patients with type 2 diabetes mellitus in England and Wales. Pharmacoepidemiol. Drug Saf. 2016, 26, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Flory, J.H.; Small, D.S.; Cassano, P.A.; Brillon, D.J.; Mushlin, A.I.; Hennessy, S. Comparative effectiveness of oral diabetes drug combinations in reducing glycosylated hemoglobin. J. Comp. Effect. Res. 2013, 3, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhu, J.; Prokop, L.J.; Murad, M.H. Pharmacologic Therapy of Diabetes and Overall Cancer Risk and Mortality: A Meta-Analysis of 265 Studies. Sci. Rep. 2015, 5, 10147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval, D.A.; D’Alessio, D.A. Physiology of Proglucagon Peptides: Role of Glucagon and GLP-1 in Health and Disease. Physiol. Rev. 2015, 95, 513–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Li, L.; Deng, K.; Xu, C.; Busse, J.W.; Vandvik, P.O.; Li, S.; Guyatt, G.H.; Sun, X. Incretin based treatments and mortality in patients with type 2 diabetes: Systematic review and meta-analysis. BMJ 2017, 357, j2499. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.J. GLP-1 receptor agonists for individualized treatment of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2012, 8, 728. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Cooper, M.E.; Del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet 2014, 383, 1068–1083. [Google Scholar] [CrossRef]
- Prasad-Reddy, L.; Isaacs, D. A clinical review of GLP-1 receptor agonists: Efficacy and safety in diabetes and beyond. Drugs Context 2015, 4, 212283. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.L.; Li, Q.; Song, S.S.; Feng, K.; Zhang, D.B.; Wang, Q.Y.; Chen, Y.H. Characterization of antimicrobial peptides isolated from the skin of the Chinese frog, Rana dybowskii. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 154, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Jin, L.; Wang, H.; Tai, S.; Liu, H.; Zhang, D. AWRK6, A Synthetic Cationic Peptide Derived from Antimicrobial Peptide Dybowskin-2CDYa, Inhibits Lipopolysaccharide-Induced Inflammatory Response. Int. J. Mol. Sci. 2018, 19, 600. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Wang, Z.; Jin, L.; Li, H.; Hwang, J.W.; Hanrahan, J.; Wang, Q. Development of a novel antimicrobial peptide AWRK6. Bang. J. Pharmacol. 2016, 11, 460. [Google Scholar] [Green Version]
- Conlon, J.M.; Mechkarska, M.; Abdel-Wahab, Y.H.; Flatt, P.R. Peptides from frog skin with potential for development into agents for Type 2 diabetes therapy. Peptides 2018, 100, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Vasu, S.; Ojo, O.O.; Moffett, R.C.; Conlon, J.M.; Flatt, P.R.; Abdel-Wahab, Y.H.A. Anti-diabetic actions of esculentin-2CHa(1–30) and its stable analogues in a diet-induced model of obesity-diabetes. Amino Acids 2017, 49, 1705–1717. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, F.M.; Dejene, E.A.; Corbin, K.L.; Nunemaker, C.S. Stress-induced dissociations between intracellular calcium signaling and insulin secretion in pancreatic islets. Cell Calcium 2015, 57, 366–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trexler, A.J.; Taraska, J.W. Regulation of insulin exocytosis by calcium-dependent protein kinase C in β cells. Cell Calcium 2017, 67, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Arruda, A.P.; Hotamisligil, G.S. Calcium Homeostasis and Organelle Function in the Pathogenesis of Obesity and Diabetes. Cell Metabolism 2015, 22, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G.; Nakashima, R.; Yuan, Q.; Marks Andrew, R. Intracellular calcium release channels: An update. J. Physiol. 2017, 595, 3041–3051. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.M.; Rex, E.A.; Devaraneni, P.; Denton, J.S.; Boodhansingh, K.E.; DeLeon, D.D.; Stanley, C.A.; Shyng, S.L. Pharmacological Correction of Trafficking Defects in ATP-sensitive Potassium Channels Caused by Sulfonylurea Receptor 1 Mutations. J. Biol. Chem. 2016, 291, 21971–21983. [Google Scholar] [CrossRef] [PubMed]
- Tengholm, A.; Gylfe, E. cAMP signalling in insulin and glucagon secretion. Diabetes Obes. Metab. 2017, 19, 42–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodson, D.J.; Mitchell, R.K.; Marselli, L.; Pullen, T.J.; Gimeno Brias, S.; Semplici, F.; Everett, K.L.; Cooper, D.M.; Bugliani, M.; Marchetti, P.; et al. ADCY5 couples glucose to insulin secretion in human islets. Diabetes 2014, 63, 3009–3021. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, K.; Shibasaki, T.; Takahashi, H.; Seino, S. Structure and functional roles of Epac2 (Rapgef4). Gene 2016, 575, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J.; Nauck, M.A. The incretin system: Glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors in type 2 diabetes. Lancet 2006, 368, 1696–1705. [Google Scholar] [CrossRef]
- Kim, H.; Toyofuku, Y.; Lynn, F.C.; Chak, E.; Uchida, T.; Mizukami, H.; Fujitani, Y.; Kawamori, R.; Miyatsuka, T.; Kosaka, Y.; et al. Serotonin regulates pancreatic β cell mass during pregnancy. Nat. Med. 2010, 16, 804–808. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Zhao, C.; Jin, L.; Zhang, H.; Miao, Q.; Liu, H.; Zhang, D. AWRK6, a Novel GLP-1 Receptor Agonist, Attenuates Diabetes by Stimulating Insulin Secretion. Int. J. Mol. Sci. 2018, 19, 3053. https://doi.org/10.3390/ijms19103053
Wang Q, Zhao C, Jin L, Zhang H, Miao Q, Liu H, Zhang D. AWRK6, a Novel GLP-1 Receptor Agonist, Attenuates Diabetes by Stimulating Insulin Secretion. International Journal of Molecular Sciences. 2018; 19(10):3053. https://doi.org/10.3390/ijms19103053
Chicago/Turabian StyleWang, Qiuyu, Chunlin Zhao, Lili Jin, Hanyu Zhang, Qifan Miao, Hongsheng Liu, and Dianbao Zhang. 2018. "AWRK6, a Novel GLP-1 Receptor Agonist, Attenuates Diabetes by Stimulating Insulin Secretion" International Journal of Molecular Sciences 19, no. 10: 3053. https://doi.org/10.3390/ijms19103053
APA StyleWang, Q., Zhao, C., Jin, L., Zhang, H., Miao, Q., Liu, H., & Zhang, D. (2018). AWRK6, a Novel GLP-1 Receptor Agonist, Attenuates Diabetes by Stimulating Insulin Secretion. International Journal of Molecular Sciences, 19(10), 3053. https://doi.org/10.3390/ijms19103053