Allergen Delivery Inhibitors: Characterisation of Potent and Selective Inhibitors of Der p 1 and Their Attenuation of Airway Responses to House Dust Mite Allergens


Abstract
:
1. Introduction
2. Results
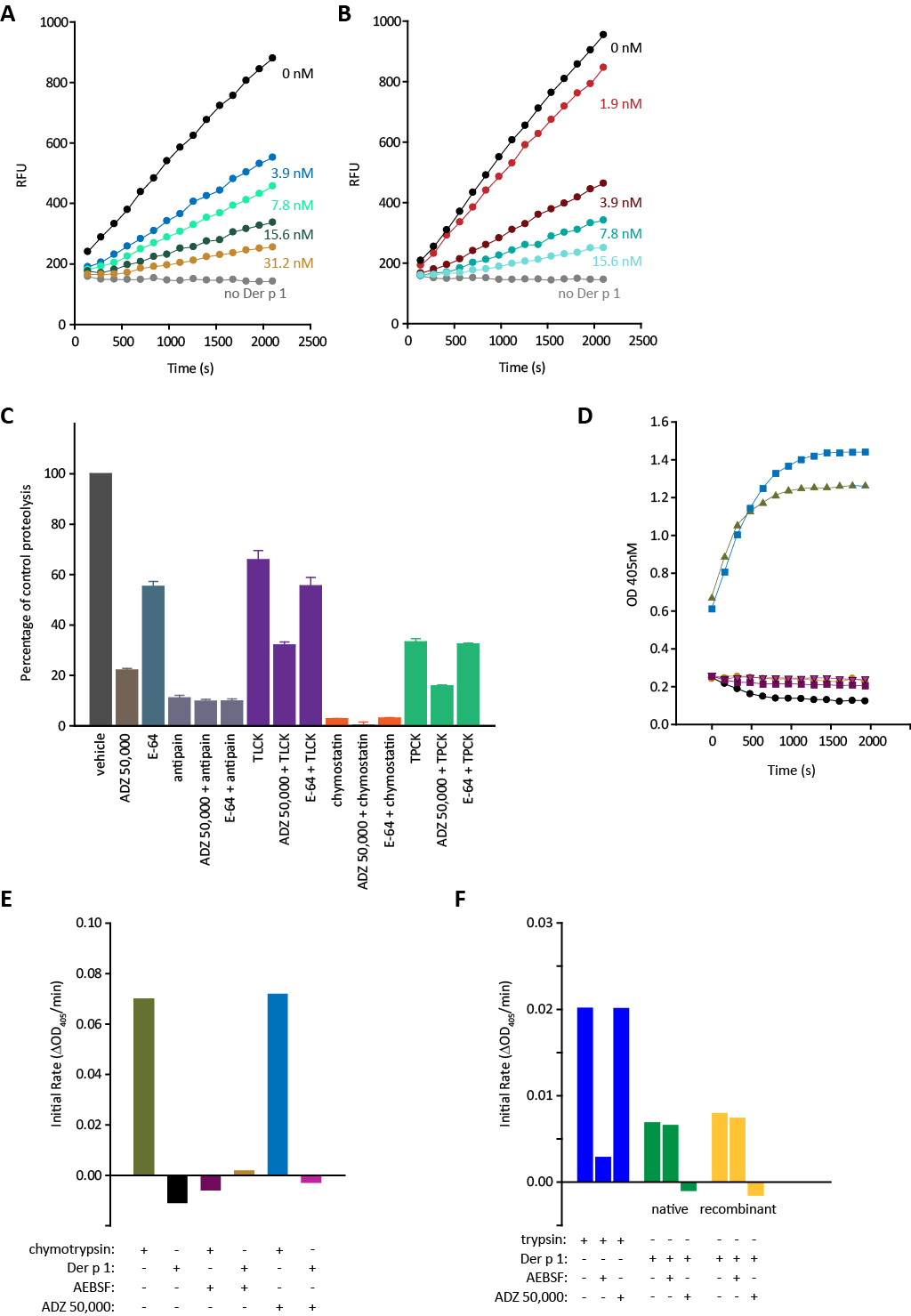
2.1. Degradation of ADZ 50,059 by Group 1 HDM Allergens and Its Inhibition
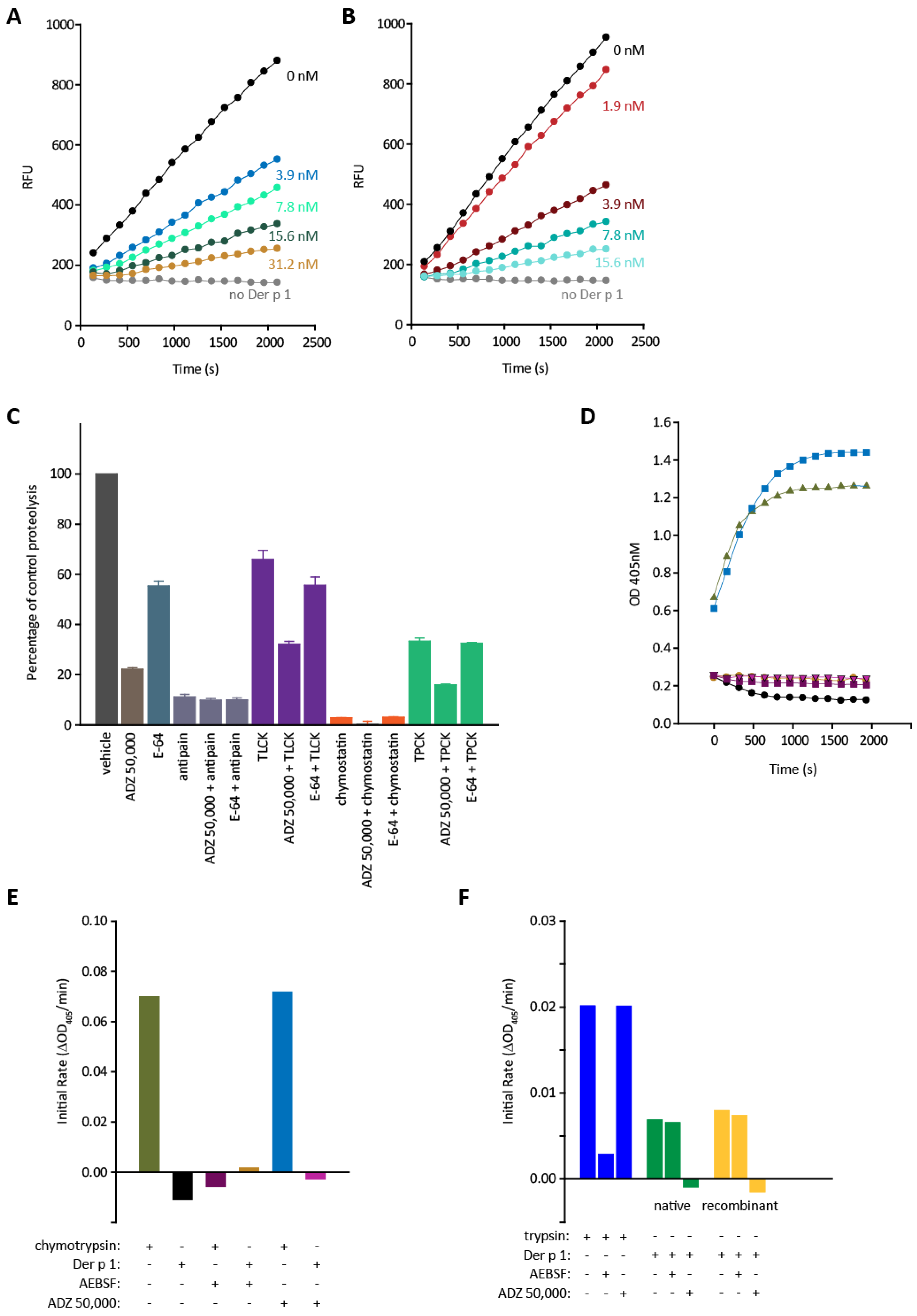
2.2. Effects of Der p 1 and Der p 2 on Barrier Properties of Human Airway Epithelial Cells and Its Modulation by ADZ 50,000
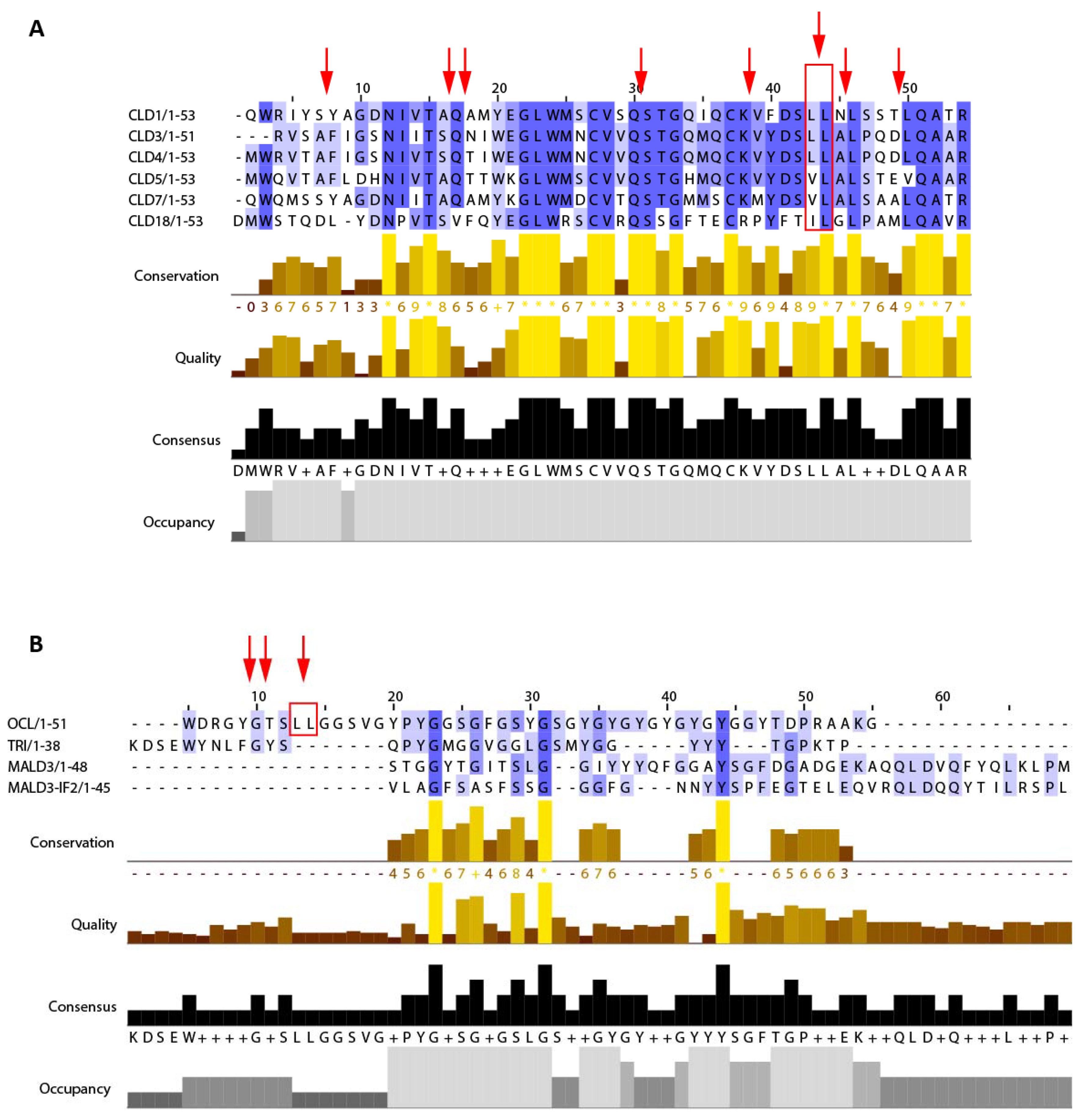
2.3. Cleavage of CLD1.1 by Der p 1
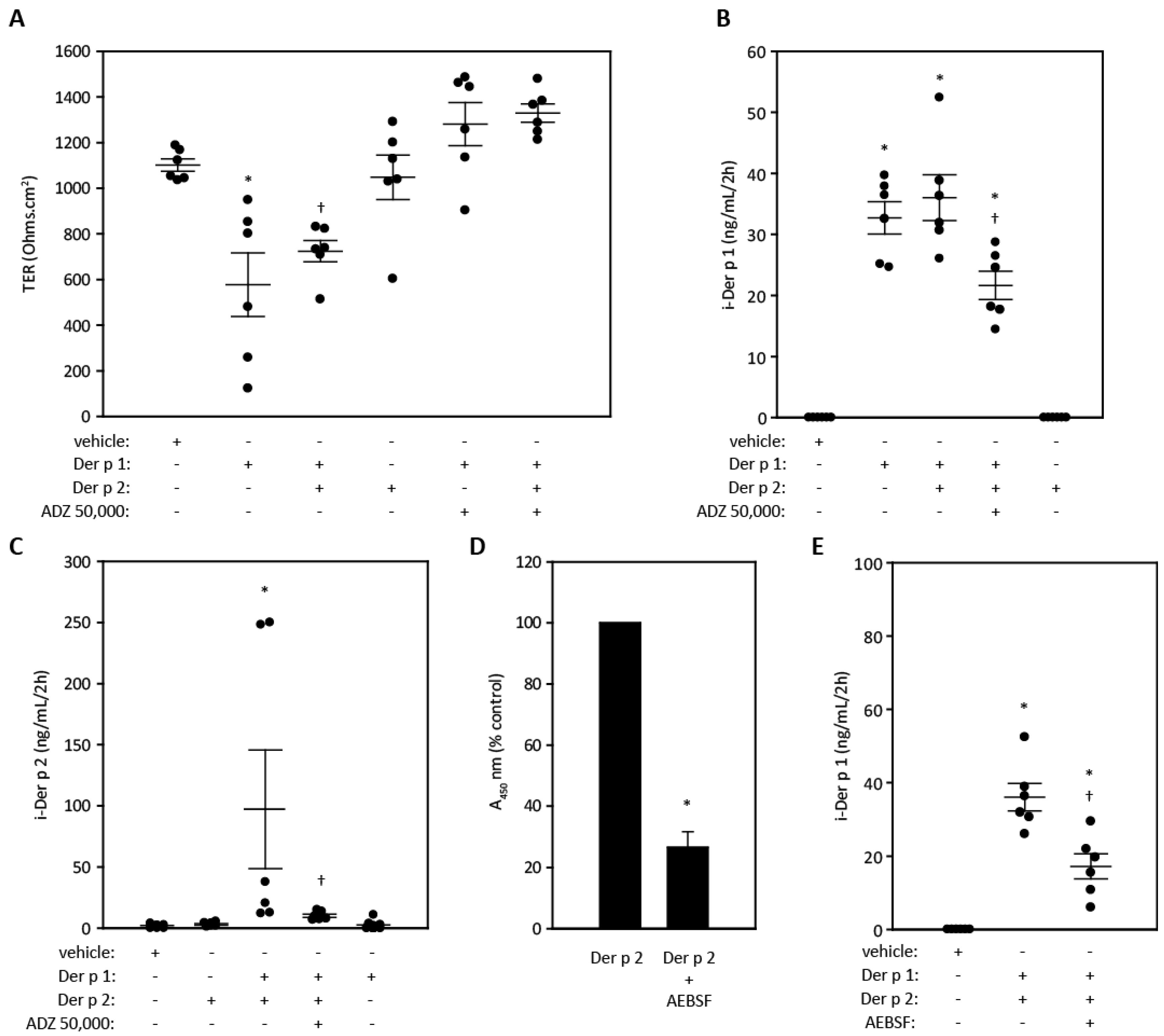
2.4. Cytokine Release from Airway Epithelial Cells and Its Modulation By ADZ 50,000
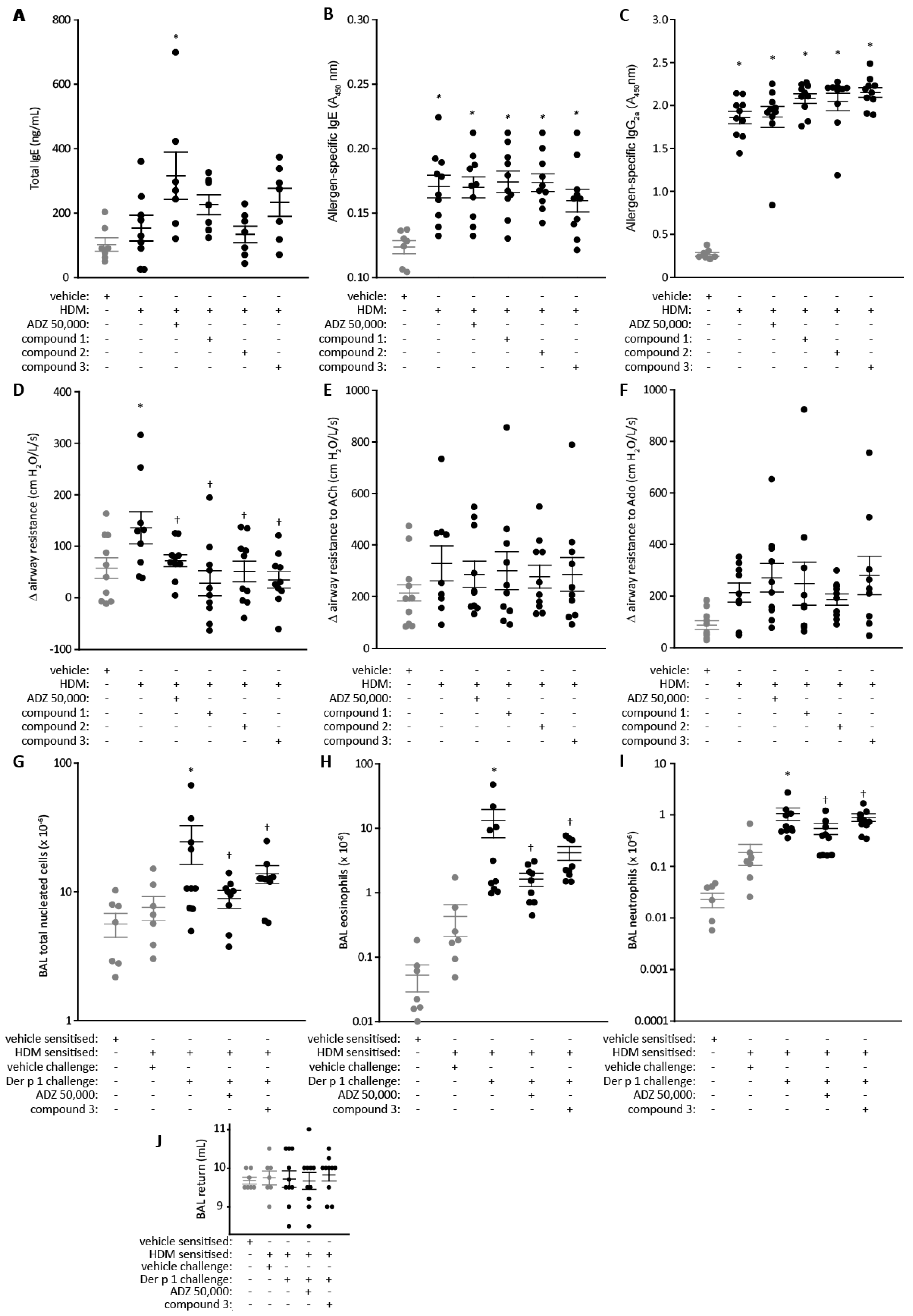
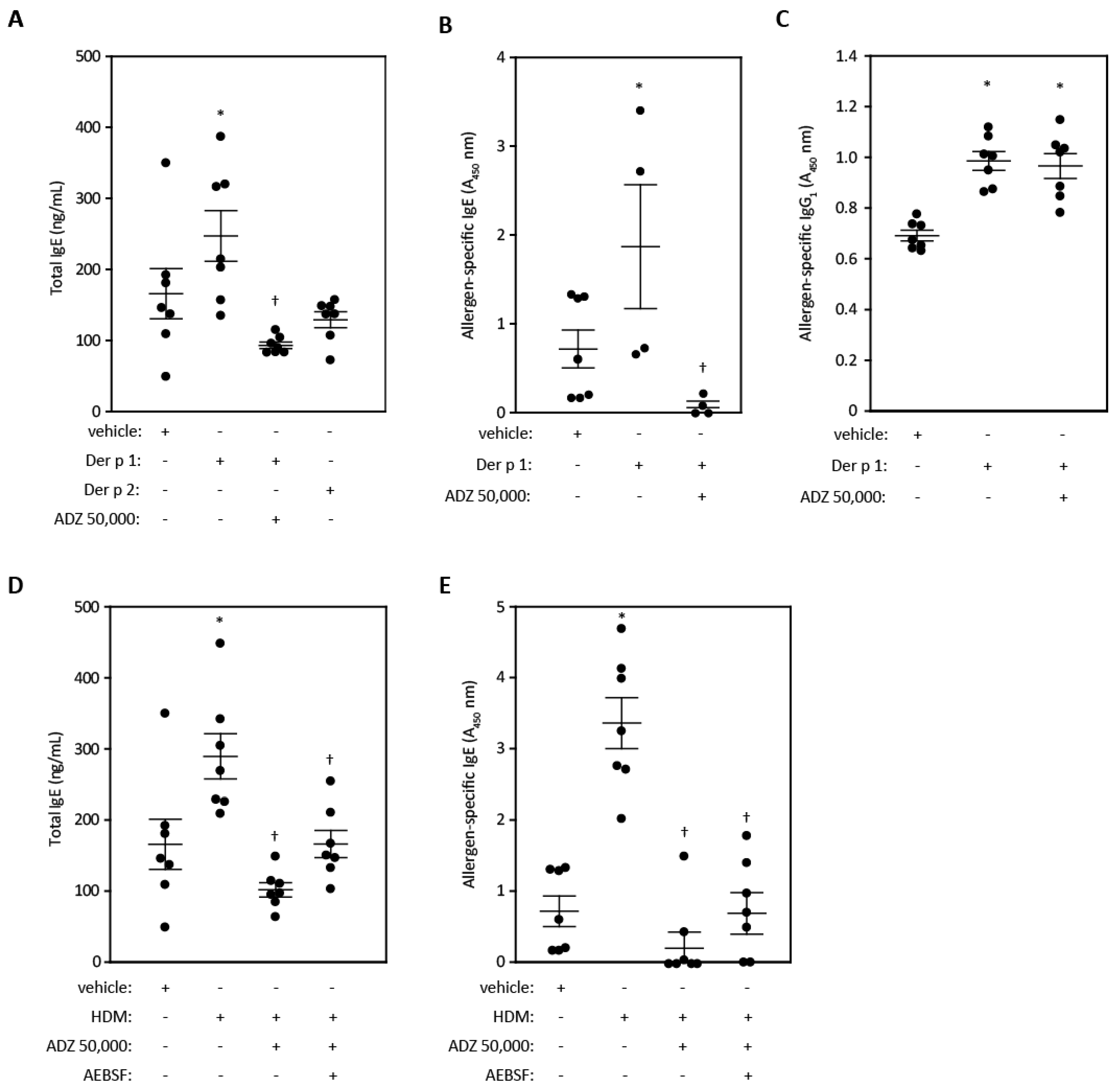
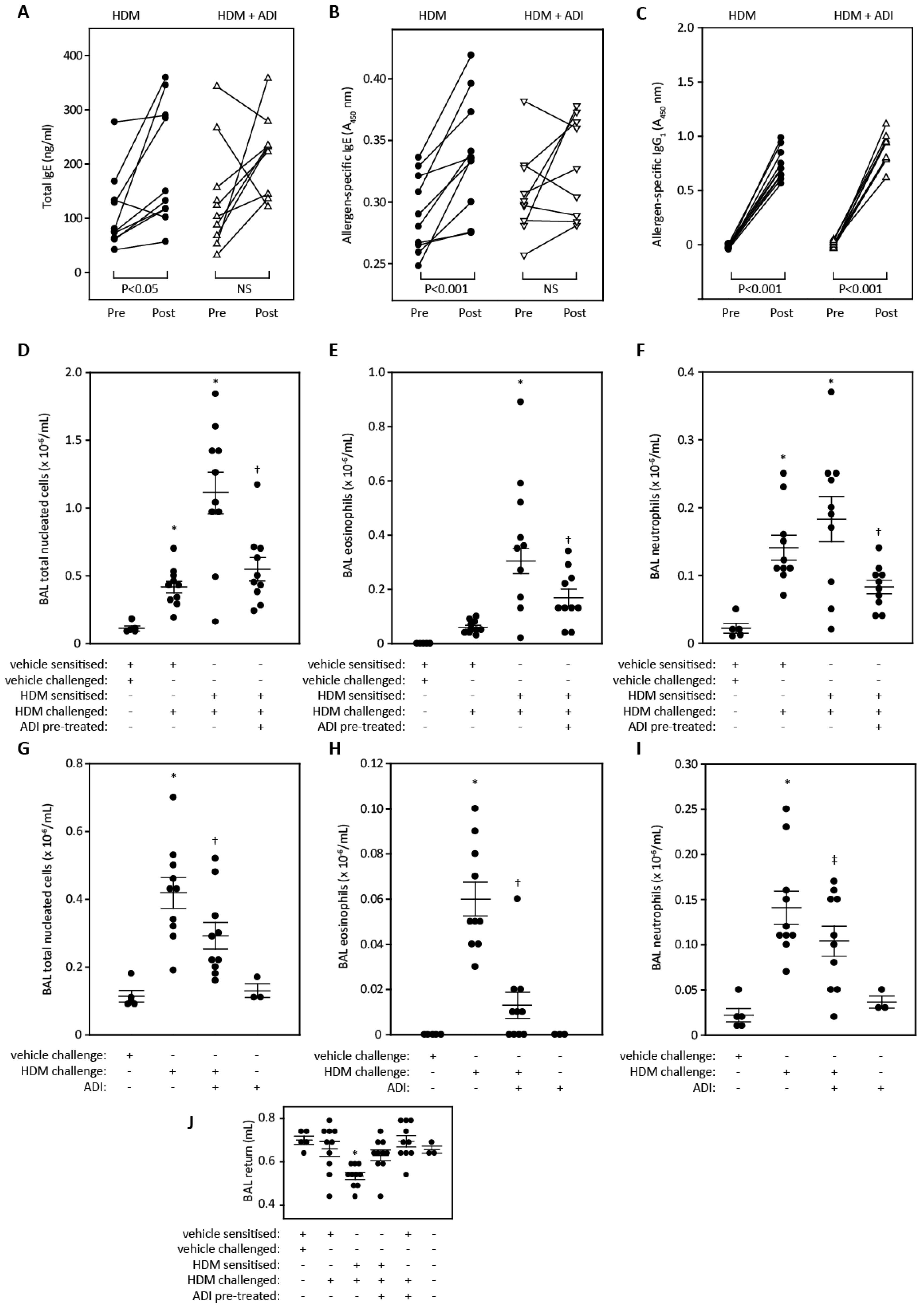
2.5. Lung Function and Cell Recruitment Studies in Sensitized Rats
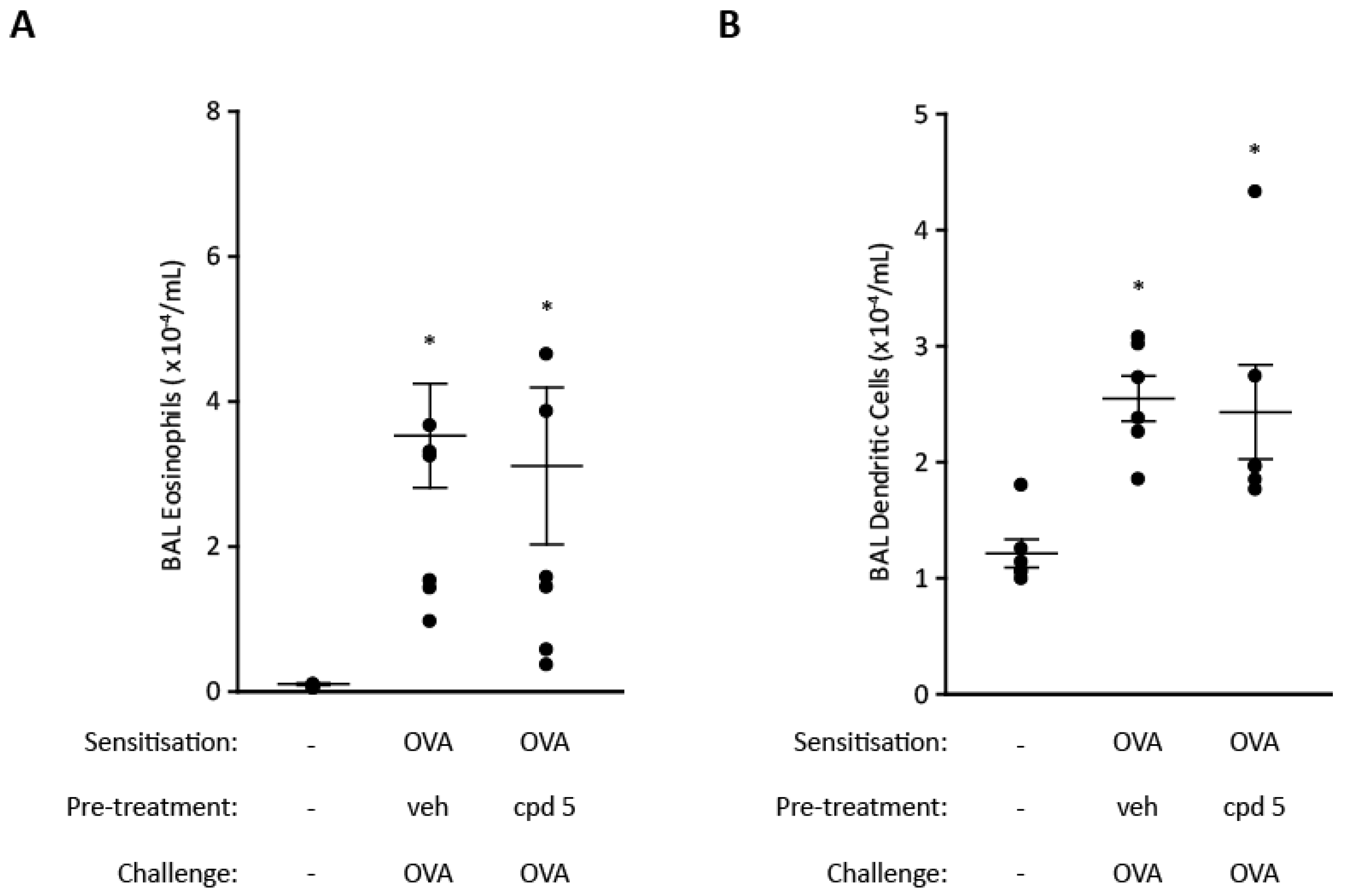
2.6. Cell Recruitment Studies in Mice
3. Discussion
4. Methods and Materials
4.1. Materials
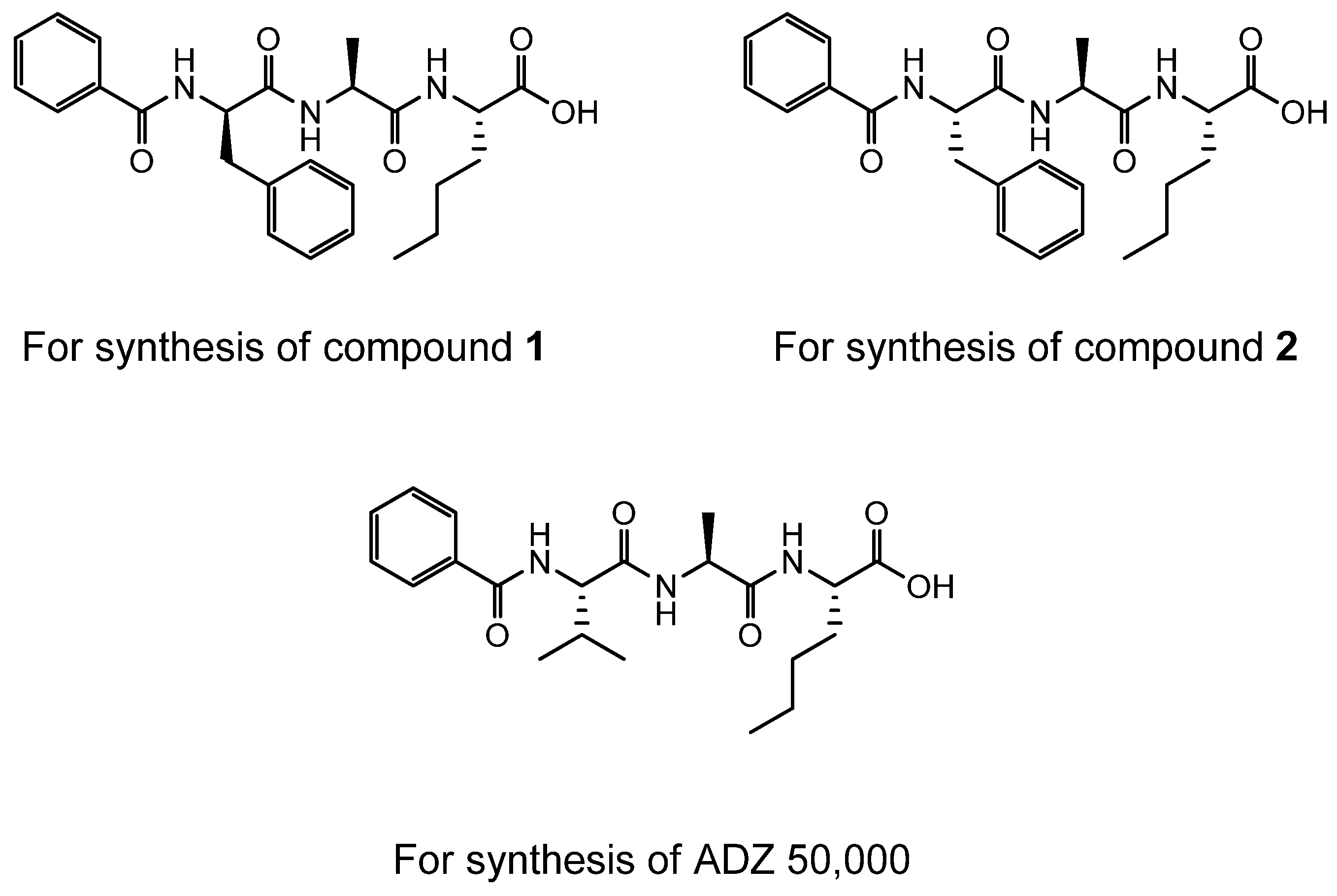
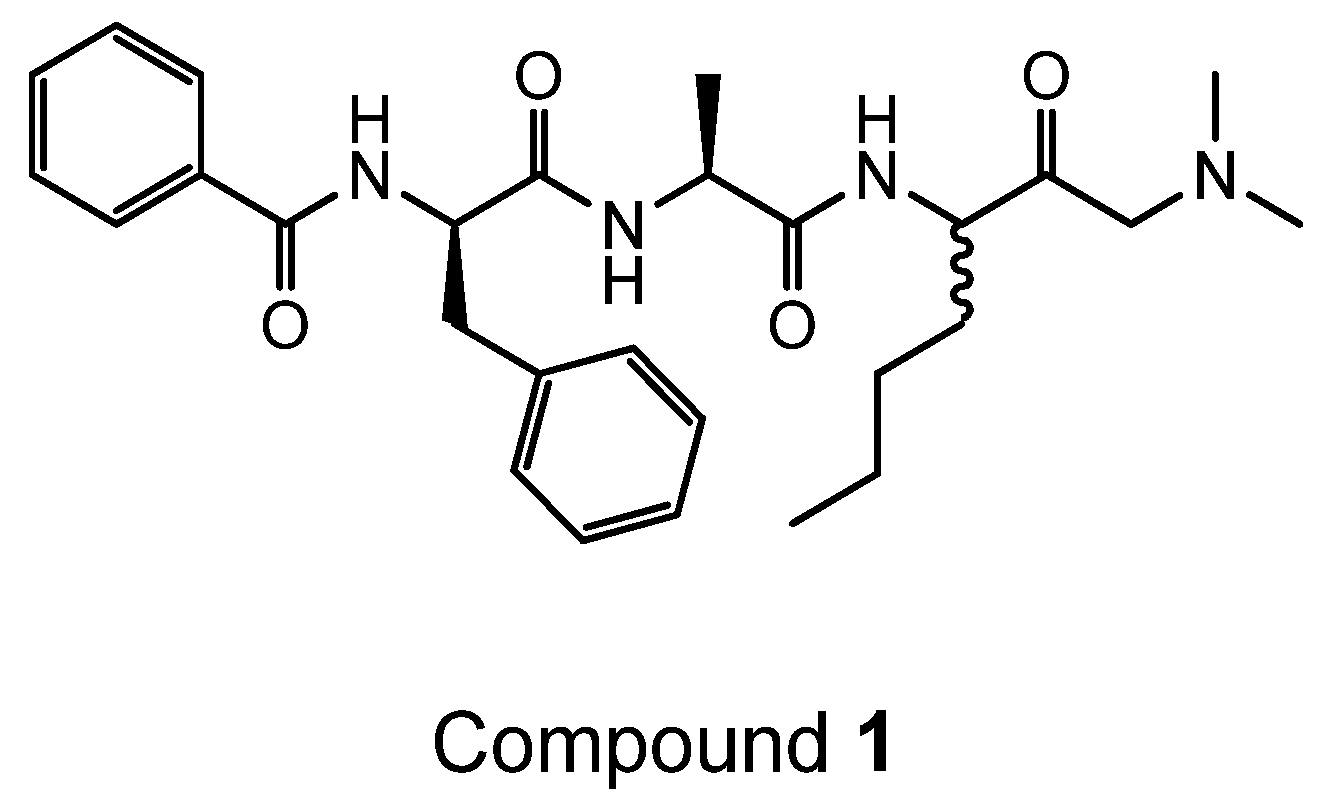
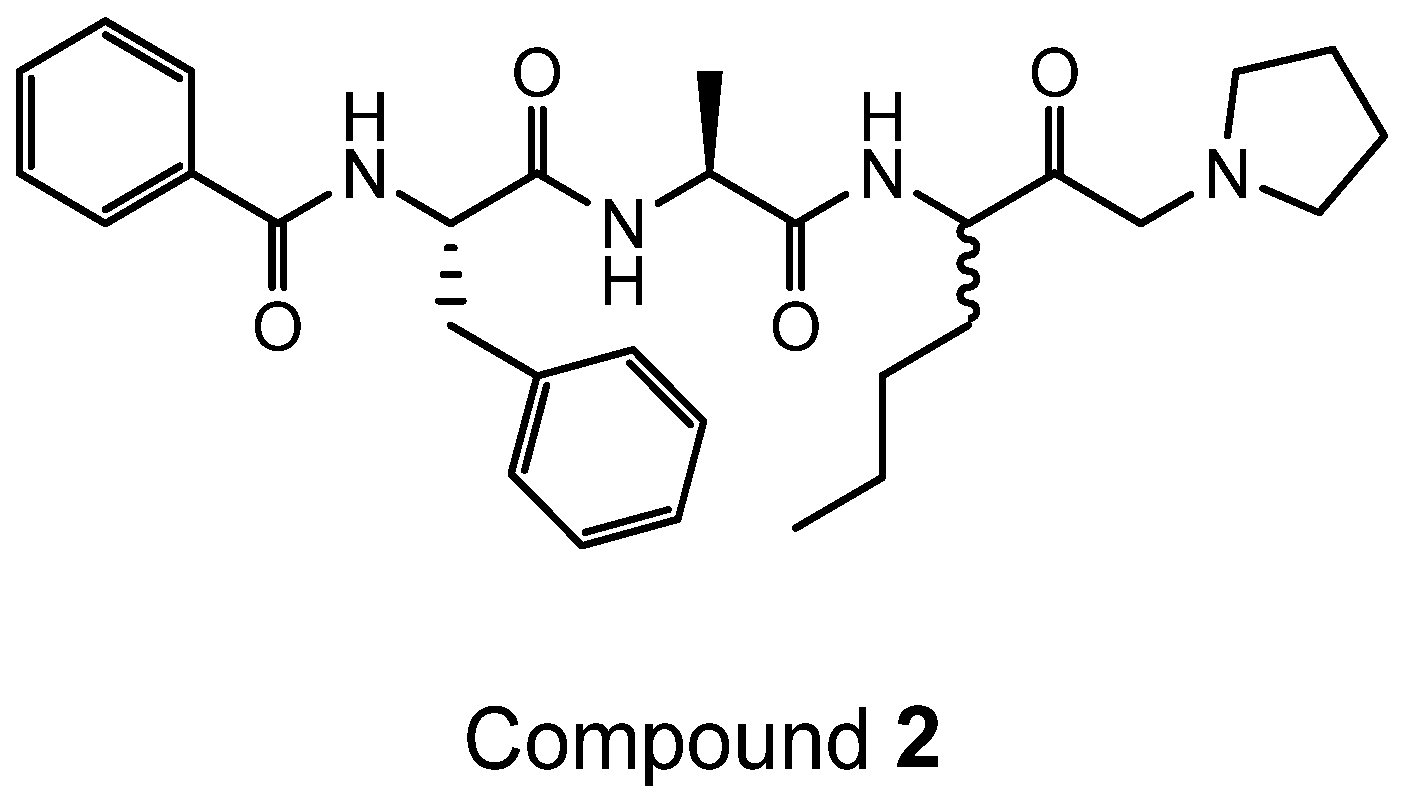
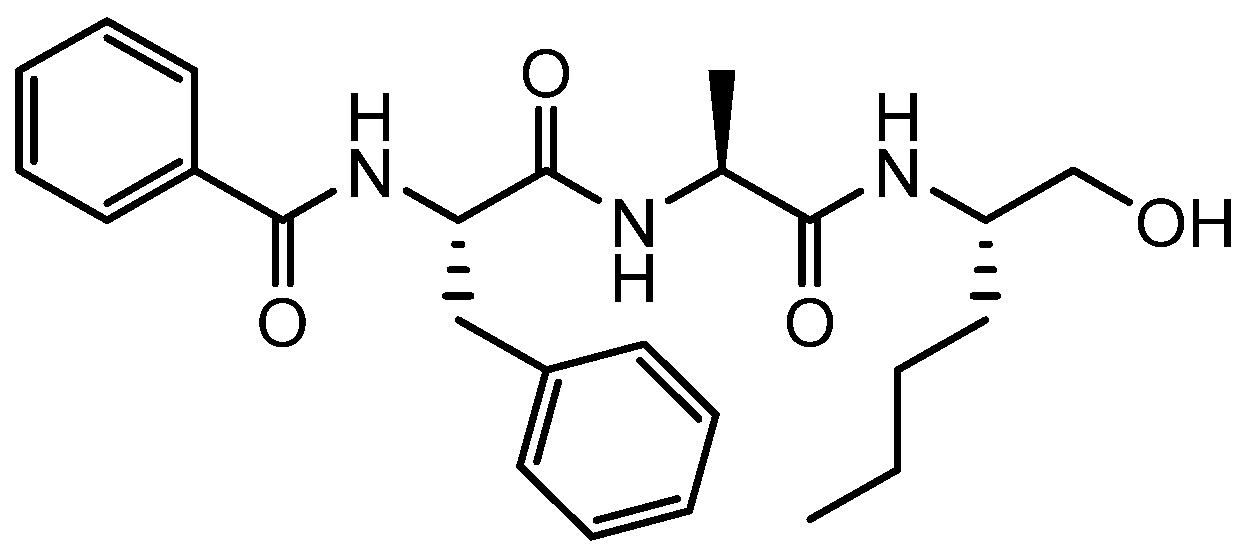
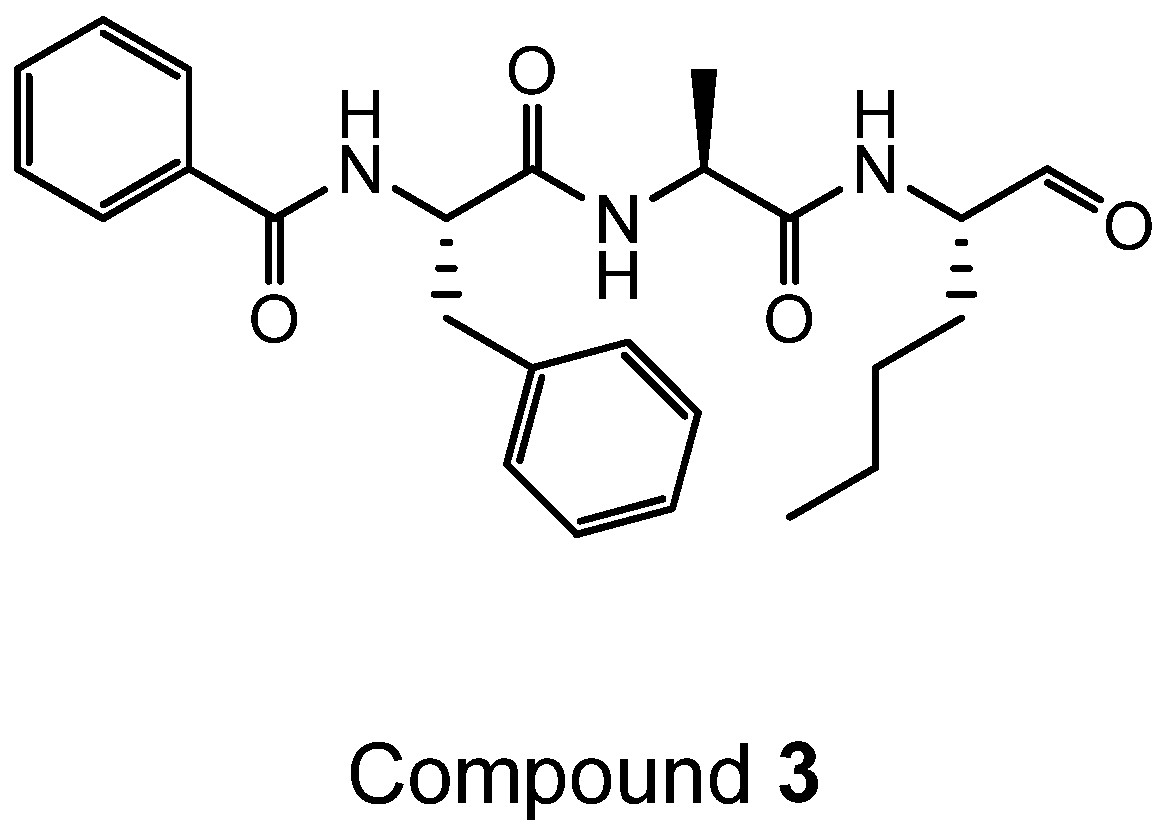
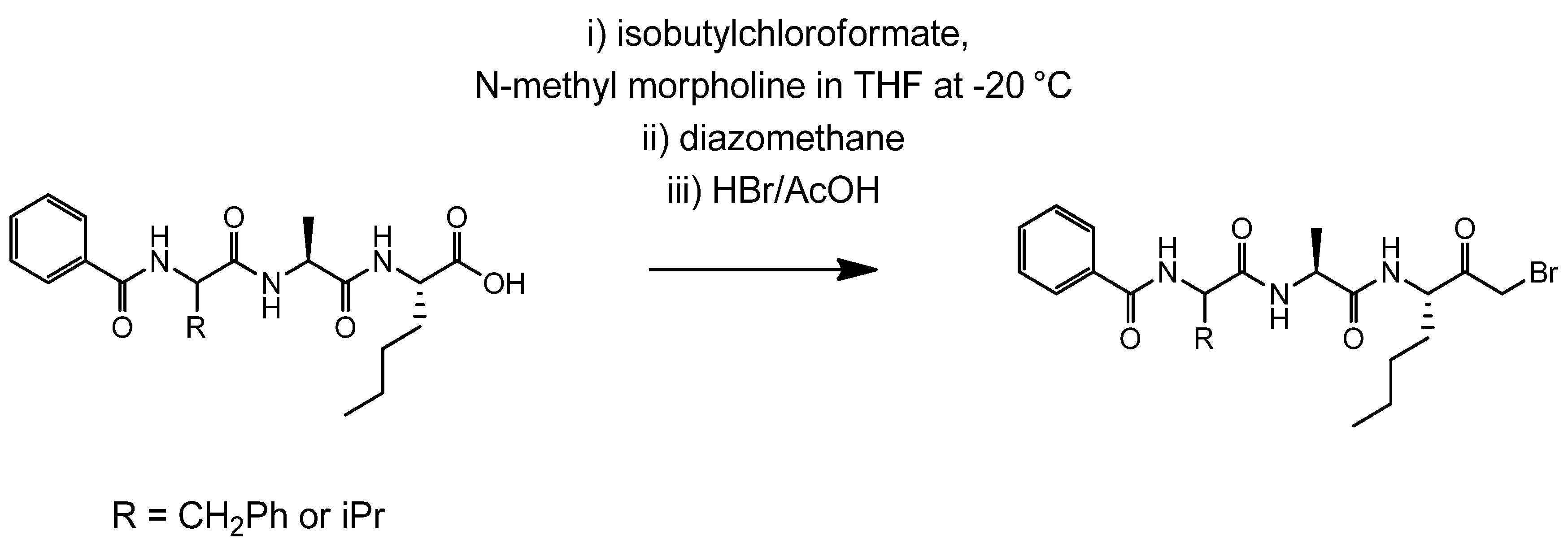
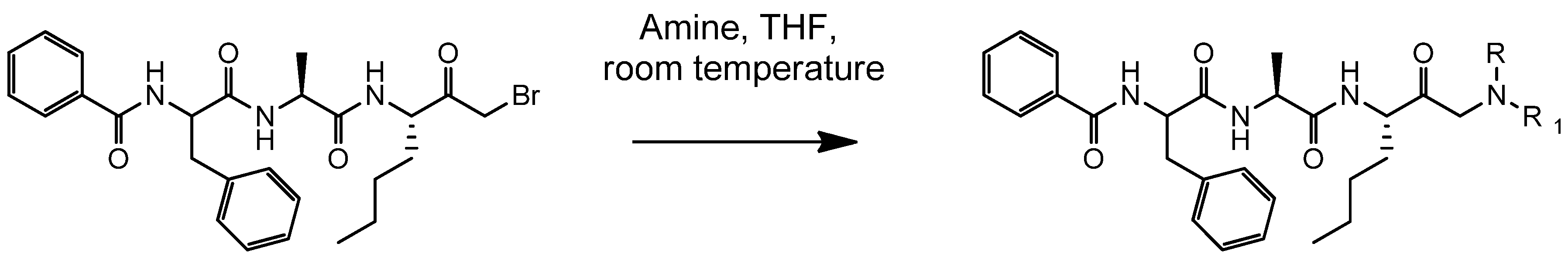
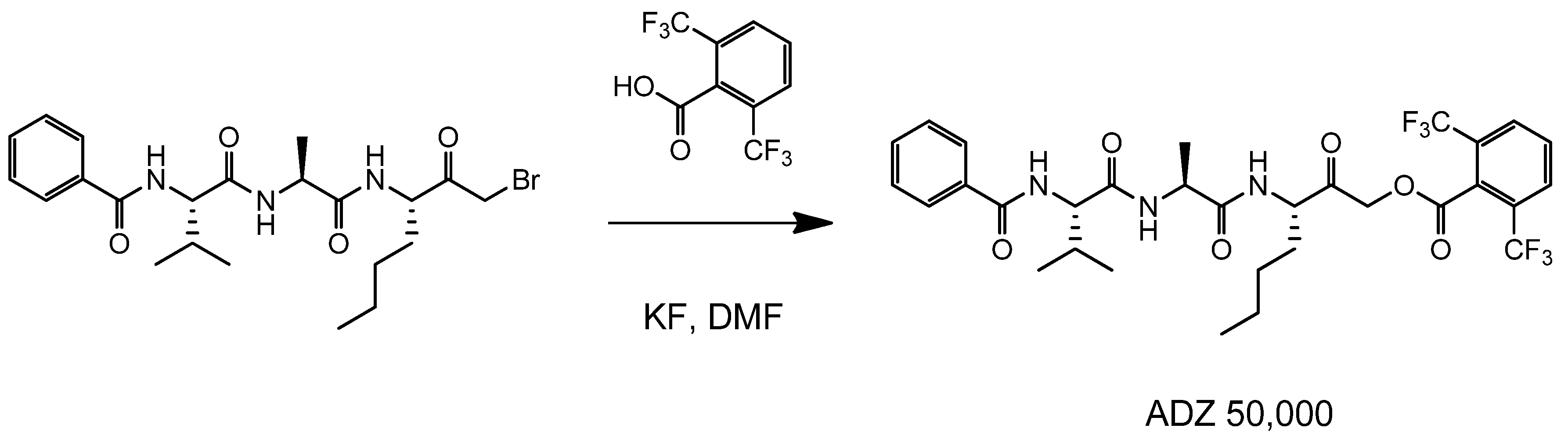
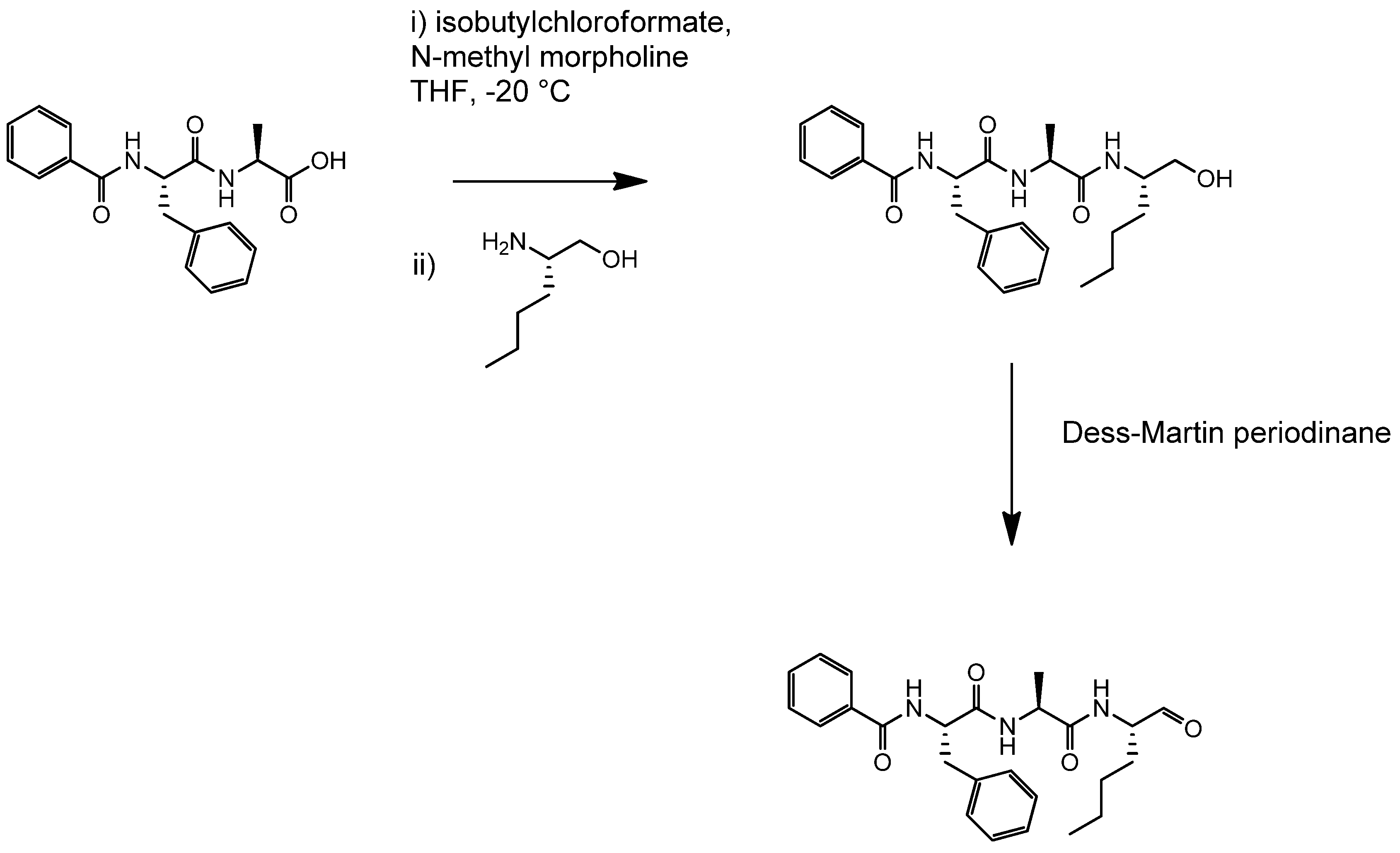
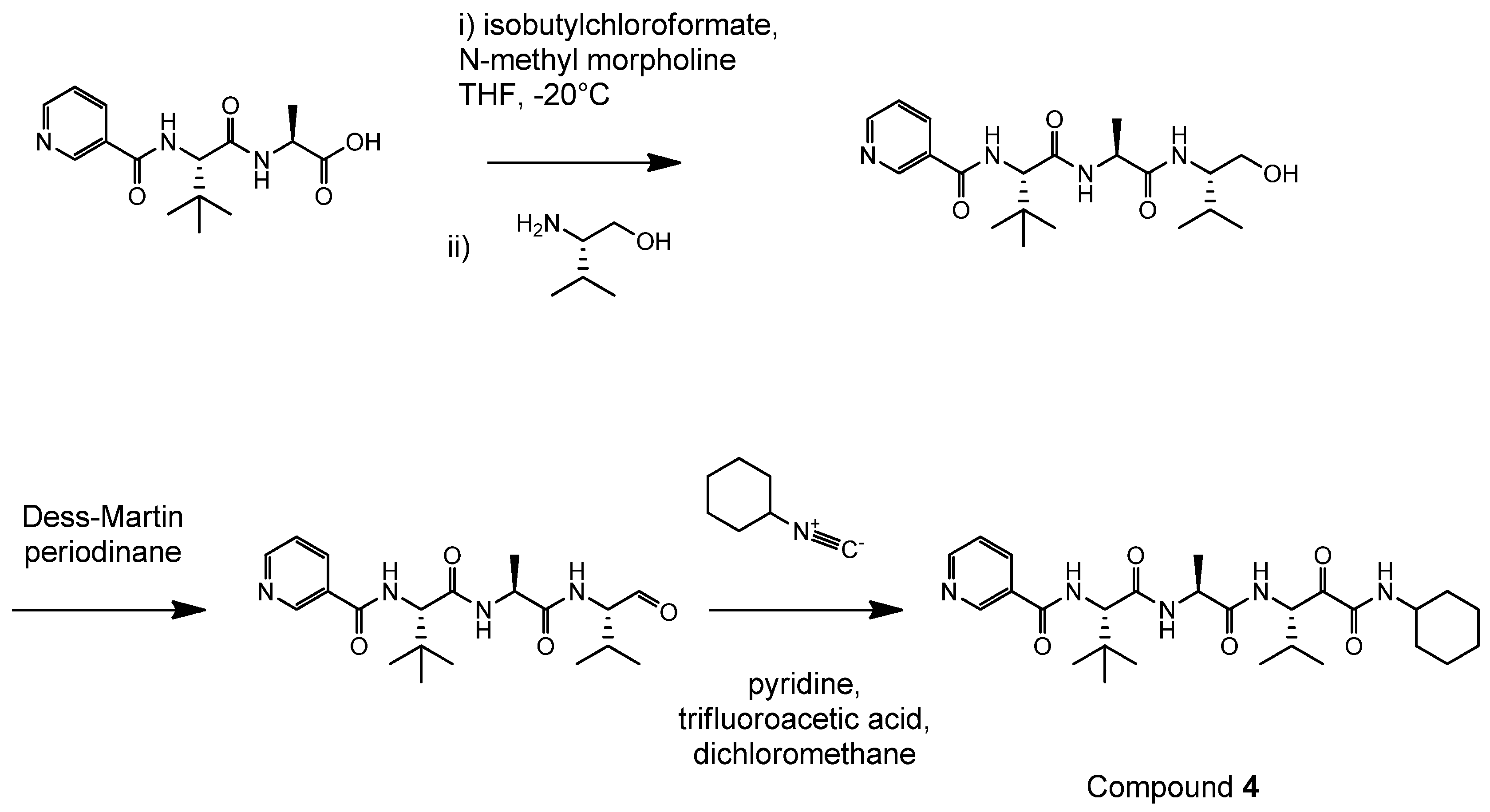
4.2. Synthesis of Der p 1 Substrate and Specific Inhibitors





4.3. Preparation of HDM Allergen and Purification of Der p 1
4.4. Der p 1 and Der f 1 Enzyme Activity Assays
4.5. Assay of Cathepsin B
4.6. Assay of Cathepsin H
4.7. Assay of Cathepsin K
4.8. Assays for Cathepsin L and Cathepsin S
4.9. Assay of Thrombin
4.10. Assay of Trypsin
4.11. Assay of Chymotrypsin
4.12. Analysis of Inhibitor Kinetics
4.13. Expression of Claudin 1.1
4.14. Cell Culture
4.15. Studies Performed In Vivo
4.16. Allergic Responses in Rats
4.17. Allergic Sensitization Studies in Mice
4.18. ELISA Measurements
4.19. BAL Analysis by Flow Cytometry
4.20. Data Presentation and Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Abz | aminobenzoyl |
| ACh | acetylcholine |
| ADI | allergen delivery inhibitor |
| Ado | adenosine |
| ADZ 50,059 | (3S,6S,9S,12S,15S,18S)-1-(2-aminophenyl)-9-butyl-18-carbamoyl-15-(4-hydroxy-3-nitrobenzyl)-12-(hydroxymethyl)-3-isopropyl-6-methyl-1,4,7,10,13,16-hexaoxo-2,5,8,11,14,17-hexaazaicosan-20-oic acid |
| AEBSF | 4-(2-aminoethyl)benzenesulphonyl fluoride |
| AMC | 7-amino-4-methylcoumarin |
| BAL | bronchoalveolar lavage |
| Bz | benzoyl |
| CH2Ph | benzyl |
| CLD1.1 | extracellular domain 1 of claudin-1 |
| DCM | dichloromethane |
| Der f | Dermatophagoides farinae |
| Der p | Dermatophagoides pteronyssinus |
| DMF | dimethylformamide |
| DMSO | dimethyl sulphoxide |
| DNP | 2,4-dinitrophenol |
| DTT | dithiothreitol |
| E-64 | trans-epoxysucciny-l-leucyl-amido (4-guanidino) butane |
| EDTA | ethylenediaminetetraacetic acid |
| EMEM | minimum essential medium with Earle’s salts |
| GM-CSF | granulocyte macrophage-colony stimulating factor |
| HBSS | Hanks’ balanced salt solution |
| HDM | house dust mite |
| HEPES | 4-(2-hydroxyethyl)-1-piperazine ethanesulphonic acid |
| HPLC | high performance liquid chromatography |
| HTS | high-throughput screening |
| i.p. | intraperitoneal |
| iPr | isopropyl |
| IL | interleukin |
| IPTG | isopropyl β-d-1-thiogalactopyranoside |
| i.t. | intratracheal |
| LC-MS | liquid chromatography-mass spectrometry |
| MALDI-TOF | matrix-assisted laser desorption ionisation time-of-flight mass spectrometry |
| MeCN | acetonitrile |
| MES | 2-morpholinoethanesulphonic acid |
| NaAc-AcH | sodium acetate-acetic acid |
| NMR | nuclear magnetic resonance |
| OVA | ovalbumin |
| PAGE | polyacrylamide gel electrophoresis |
| PAR | protease-activated receptor |
| PBS | phosphate-buffered saline |
| SDS | sodium dodecyl sulphate |
| TER | transepithelial electrical resistance |
| THF | tetrahydrofuran |
| TJ | tight junction |
| TLCK | Nα-p-tosyl-l-lysine choromethyl ketone |
| TPCK | N-p-tosyl-l-phenylalanine chloromethyl ketone |
References
- Stewart, G.A.; Richardson, J.P.; Zhang, J.; Robinson, C. The structure and function of allergens. In Middleton’s Allergy—Principles and Practice; Adkinson, N.F., Bochner, B.S., Burks, A.W., Busse, W.W., Holgate, S.T., Lemanske, R.F., O’Hehir, R.E., Eds.; Elsevier Saunders: Philadelphia, PA, USA, 2014; Volume 8, pp. 398–429. [Google Scholar]
- Zhang, J.; Chen, J.; Newton, G.K.; Perrior, T.R.; Robinson, C. Allergen Delivery Inhibitors: A rationale for targeting sentinel innate immune signaling of Group 1 house dust mite allergens through structure-based protease inhibitor design. Mol. Pharmacol. 2018, 94, 1007–1030. [Google Scholar] [CrossRef] [PubMed]
- Fattouh, R.; Pouladi, M.A.; Alvarez, D.; Johnson, J.R.; Walker, T.D.; Goncharova, S.; Inman, M.D.; Jordana, M. House dust mite facilitates ovalbumin-specific allergic sensitization and airway inflammation. Am. J. Respir. Crit. Care Med. 2005, 172, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Gough, L.; Schulz, O.; Sewell, H.F.; Shakib, F. The cysteine protease activity of the major dust mite allergen Der p 1 selectively enhances the immunoglobulin E antibody response. J. Exp. Med. 1999, 190, 1897–1902. [Google Scholar] [CrossRef] [PubMed]
- Gough, L.; Sewell, H.F.; Shakib, F. The proteolytic activity of the major dust mite allergen Der p 1 enhances the IgE antibody response to a bystander antigen. Clin. Exp. Allergy 2001, 31, 1594–1598. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.Y.; Tam, M.F.; Chou, H.; Peng, H.J.; Su, S.N.; Perng, D.W.; Shen, H.D. Pen ch 13 allergen induces secretion of mediators and degradation of occludin protein of human lung epithelial cells. Allergy 2006, 61, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, M.; Rossi, G.A.; Cozzani, S.; Pulvirenti, G.; Fasce, L. Age-dependent tendency to become sensitized to other classes of aeroallergens in atopic asthmatic children. Ann. Allergy Asthma Immunol. 1999, 83, 335–340. [Google Scholar] [CrossRef]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. EAACI Molecular Allergology User’s Guide. Pediatric Allergy Immunol. 2016, 27 (Suppl. 23), 1–250. [Google Scholar] [CrossRef] [PubMed]
- Purello-D’Ambrosio, F.; Gangemi, S.; Merendino, R.A.; Isola, S.; Puccinelli, P.; Parmiani, S.; Ricciardi, L. Prevention of new sensitizations in monosensitized subjects submitted to specific immunotherapy or not. A retrospective study. Clin. Exp. Allergy 2001, 31, 1295–1302. [Google Scholar] [CrossRef] [PubMed]
- Wickman, M.; Lupinek, C.; Andersson, N.; Belgrave, D.; Asarnoj, A.; Benet, M.; Pinart, M.; Wieser, S.; Garcia-Aymerich, J.; Baar, A.; et al. Detection of IgE Reactivity to a Handful of Allergen Molecules in Early Childhood Predicts Respiratory Allergy in Adolescence. EBioMedicine 2017, 26, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.; Zhang, J.; Newton, G.K.; Perrior, T.R. Nonhuman targets in allergic lung conditions. Future Med. Chem. 2013, 5, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, J.; Robinson, C. Fundamental events in the interaction between house dust mite group 1 allergens and innate defences in the airways. Int. J. Mol. Sci. 2018. submitted for publication. [Google Scholar]
- Pichavant, M.; Charbonnier, A.S.; Taront, S.; Brichet, A.; Wallaert, B.; Pestel, J.; Tonnel, A.B.; Gosset, P. Asthmatic bronchial epithelium activated by the proteolytic allergen Der p 1 increases selective dendritic cell recruitment. J. Allergy Clin. Immunol. 2005, 115, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Winton, H.L.; Soeller, C.; Tovey, E.R.; Gruenert, D.C.; Thompson, P.J.; Stewart, G.A.; Taylor, G.W.; Garrod, D.R.; Cannell, M.B.; et al. Der p 1 facilitates transepithelial allergen delivery by disruption of tight junctions. J. Clin. Investig. 1999, 104, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweerus, K.; Lachowicz-Scroggins, M.; Gordon, E.; LaFemina, M.; Huang, X.; Parikh, M.; Kanegai, C.; Fahy, J.V.; Frank, J.A. Claudin-18 deficiency is associated with airway epithelial barrier dysfunction and asthma. J. Allergy Clin. Immunol. 2017, 139, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Kubo, A.; Nagao, K.; Yokouchi, M.; Sasaki, H.; Amagai, M. External antigen uptake by Langerhans cells with reorganization of epidermal tight junction barriers. J. Exp. Med. 2009, 206, 2937–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, K.; Kojima, T.; Go, M.; Murata, M.; Ichimiya, S.; Himi, T.; Sawada, N. HLA-DR- and CD11c-positive dendritic cells penetrate beyond well-developed epithelial tight junctions in human nasal mucosa of allergic rhinitis. J. Histochem. Cytochem. 2005, 53, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Brandner, J.M.; Zorn-Kruppa, M.; Yoshida, T.; Moll, I.; Beck, L.A.; De Benedetto, A. Epidermal tight junctions in health and disease. Tissue Barriers 2015, 3, e974451. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, A.; Slifka, M.K.; Rafaels, N.M.; Kuo, I.H.; Georas, S.N.; Boguniewicz, M.; Hata, T.; Schneider, L.C.; Hanifin, J.M.; Gallo, R.L.; et al. Reductions in claudin-1 may enhance susceptibility to herpes simplex virus 1 infections in atopic dermatitis. J. Allergy Clin. Immunol. 2011, 128, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, J.; Tachie-Menson, T.; Shukla, N.; Garrod, D.R.; Robinson, C. Allergen-dependent oxidant formation requires purinoceptor activation of ADAM 10 and prothrombin. J. Allergy Clin. Immunol. 2017, 139, 2023–2026. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, J.; Allen-Philbey, K.; Perera Baruhupolage, C.; Tachie-Menson, T.; Mangat, S.C.; Garrod, D.R.; Robinson, C. Innate generation of thrombin and intracellular oxidants in airway epithelium by allergen Der p 1. J. Allergy Clin. Immunol. 2016, 138, 1224–1227. [Google Scholar] [CrossRef] [PubMed]
- Newton, G.K.; Perrior, T.R.; Jenkins, K.; Major, M.R.; Key, R.E.; Stewart, M.R.; Firth-Clark, S.; Lloyd, S.M.; Zhang, J.; Francis-Newton, N.J.; et al. The discovery of potent, selective, and reversible inhibitors of the house dust mite peptidase allergen Der p 1: An innovative approach to the treatment of allergic asthma. J. Med. Chem. 2014, 57, 9447–9462. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.; Noon, L. Further observation on the treatment of hay-fever by hypodermic inoculation of pollen vaccine. Lancet 1911, 2, 1572–1573. [Google Scholar] [CrossRef]
- Wan, H.; Winton, H.L.; Soeller, C.; Stewart, G.A.; Thompson, P.J.; Gruenert, D.C.; Cannell, M.B.; Garrod, D.R.; Robinson, C. Tight junction properties of the immortalized human bronchial epithelial cell lines Calu-3 and 16HBE14o-. Eur. Respir. J. 2000, 15, 1058–1068. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Winton, H.L.; Soeller, C.; Taylor, G.W.; Gruenert, D.C.; Thompson, P.J.; Cannell, M.B.; Stewart, G.A.; Garrod, D.R.; Robinson, C. The transmembrane protein occludin of epithelial tight junctions is a functional target for serine peptidases from faecal pellets of Dermatophagoides pteronyssinus. Clin. Exp. Allergy 2001, 31, 279–294. [Google Scholar] [CrossRef] [PubMed]
- Asokananthan, N.; Graham, P.T.; Stewart, D.J.; Bakker, A.J.; Eidne, K.A.; Thompson, P.J.; Stewart, G.A. House dust mite allergens induce proinflammatory cytokines from respiratory epithelial cells: The cysteine protease allergen, Der p 1, activates protease-activated receptor (PAR)-2 and inactivates PAR-1. J. Immunol. 2002, 169, 4572–4578. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H. Function of proton channels in lung epithelia. WIREs Membr. Transp. Signal. 2012, 1, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Song, Y.; Verkman, A.S. Airway surface liquid pH in well-differentiated airway epithelial cell cultures and mouse trachea. Am. J. Physiol. Cell Physiol. 2001, 281, C1504–C1511. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Thiagarajah, J.; Verkman, A.S. Sodium and chloride concentrations, pH, and depth of airway surface liquid in distal airways. J. Gen. Physiol. 2003, 122, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Schulz, O.; Sewell, H.F.; Shakib, F. A sensitive fluorescent assay for measuring the cysteine protease activity of Der p 1, a major allergen from the dust mite Dermatophagoides pteronyssinus. Mol. Pathol. 1998, 51, 222–224. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.F.; Fang, K.; Malik, R.; Synder, A.; Malhotra, N.; Platts-Mills, T.A.E.; Gaston, B. Endogenous airway acidification. Implications for asthma pathophysiology. Am. J. Respir. Crit. Care Med. 2000, 161, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, J.; Mangat, S.C.; Perera Baruhupolage, C.; Garrod, D.R.; Robinson, C. Pathways of airway oxidant formation by house dust mite allergens and viral RNA converge through myosin motors, pannexons and Toll-like receptor 4. Immun. Inflamm. Dis. 2018, 6, 276–296. [Google Scholar] [CrossRef] [PubMed]
- Gough, L.; Campbell, E.; Bayley, D.; Van Heeke, G.; Shakib, F. Proteolytic activity of the house dust mite allergen Der p 1 enhances allergenicity in a mouse inhalation model. Clin. Exp. Allergy 2003, 33, 1159–1163. [Google Scholar] [CrossRef] [PubMed]
- Kheradmand, F.; Kiss, A.; Xu, J.; Lee, S.H.; Kolattukudy, P.E.; Corry, D.B. A protease-activated pathway underlying Th cell type 2 activation and allergic lung disease. J. Immunol. 2002, 169, 5904–5911. [Google Scholar] [CrossRef] [PubMed]
- Kurup, V.P.; Xia, J.Q.; Shen, H.D.; Rickaby, D.A.; Henderson, J.D., Jr.; Fink, J.N.; Chou, H.; Kelly, K.J.; Dawson, C.A. Alkaline serine proteinase from Aspergillus fumigatus has synergistic effects on Asp-f-2-induced immune response in mice. Int. Arch. Allergy Immunol. 2002, 129, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.; Farmer, K.; MacDonald, L.; Kalsheker, N.; Pritchard, D.; Haslett, C.; Lamb, J.; Sallenave, J.M. House dust mite Der p 1 downregulates defenses of the lung by inactivating elastase inhibitors. Am. J. Respir. Cell Mol. Biol. 2003, 29, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Kalsheker, N.A.; Deam, S.; Chambers, L.; Sreedharan, S.; Brocklehurst, K.; Lomas, D.A. The house dust mite allergen Der p1 catalytically inactivates alpha 1-antitrypsin by specific reactive centre loop cleavage: A mechanism that promotes airway inflammation and asthma. Biochem. Biophys. Res. Commun. 1996, 221, 59–61. [Google Scholar] [CrossRef] [PubMed]
- He, J.S.; Subramaniam, S.; Narang, V.; Srinivasan, K.; Saunders, S.P.; Carbajo, D.; Wen-Shan, T.; Hidayah Hamadee, N.; Lum, J.; Lee, A.; et al. IgG1 memory B cells keep the memory of IgE responses. Nat. Commun. 2017, 8, 641. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.W.; Tjota, M.Y.; Sperling, A.I. The contribution of allergen-specific IgG to the development of th2-mediated airway inflammation. J. Allergy 2012, 2012, 236075. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.C.; Horton, D.; Harrop, R.; Peachell, P.T.; Helm, B.A. Potential allergens stimulate the release of mediators of the allergic response from cells of mast cell lineage in the absence of sensitization with antigen-specific IgE. Eur. J. Immunol. 1996, 26, 2972–2980. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.B.; Lerner, E.A. Activation of mas-related G-protein-coupled receptors by the house dust mite cysteine protease Der p1 provides a new mechanism linking allergy and inflammation. J. Biol. Chem. 2017, 292, 17399–17406. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, J.L. Observations on itching produced by cowhage, and on the part played by histamine as a mediator of the itch sensation. Br. J. Pharmacol. Chemother. 1953, 8, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Winton, H.L.; Soeller, C.; Gruenert, D.C.; Thompson, P.J.; Cannell, M.B.; Stewart, G.A.; Garrod, D.R.; Robinson, C. Quantitative structural and biochemical analyses of tight junction dynamics following exposure of epithelial cells to house dust mite allergen Der p 1. Clin. Exp. Allergy 2000, 30, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. Allergens and the airway epithelium response: Gateway to allergic sensitization. J. Allergy Clin. Immunol. 2014, 134, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Daugherty, B.L.; Mateescu, M.; Patel, A.S.; Wade, K.; Kimura, S.; Gonzales, L.W.; Guttentag, S.; Ballard, P.L.; Koval, M. Developmental regulation of claudin localization by fetal alveolar epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, L1266–1273. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Chiba, S.; Ebina, M.; Furuse, M.; Nukiwa, T. Altered expression of tight junction molecules in alveolar septa in lung injury and fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L193–L205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soini, Y. Claudins in lung diseases. Respir. Res. 2011, 12, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balda, M.S.; Whitney, J.A.; Flores, C.; Gonzalez, S.; Cereijido, M.; Matter, K. Functional dissociation of paracellular permeability and transepithelial electrical resistance and disruption of the apical-basolateral intramembrane diffusion barrier by expression of a mutant tight junction membrane protein. J. Cell Biol. 1996, 134, 1031–1049. [Google Scholar] [CrossRef] [PubMed]
- Balda, M.S.; Flores-Maldonado, C.; Cereijido, M.; Matter, K. Multiple domains of occludin are involved in the regulation of paracellular permeability. J. Cell Biochem. 2000, 78, 85–96. [Google Scholar] [CrossRef]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, C.R.; Horton, H.; Jones, R.M.; Pritchard, D.I. Heterogeneous proteolytic specificity and activity of the house dust mite proteinase allergen Der p I. Clin. Exp. Allergy 1997, 27, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Meno, K.; Thorsted, P.B.; Ipsen, H.; Kristensen, O.; Larsen, J.N.; Spangfort, M.D.; Gajhede, M.; Lund, K. The crystal structure of recombinant proDer p 1, a major house dust mite proteolytic allergen. J. Immunol. 2005, 175, 3835–3845. [Google Scholar] [CrossRef] [PubMed]
- de Halleux, S.; Stura, E.; VanderElst, L.; Carlier, V.; Jacquemin, M.; Saint-Remy, J.M. Three-dimensional structure and IgE-binding properties of mature fully active Der p 1, a clinically relevant major allergen. J. Allergy Clin. Immunol. 2006, 117, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Ring, P.C.; Wan, H.; Schou, C.; Kroll Kristensen, A.; Roepstorff, P.; Robinson, C. The 18-kDa form of cat allergen Felis domesticus 1 (Fel d 1) is associated with gelatin- and fibronectin-degrading activity. Clin. Exp. Allergy 2000, 30, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.C.; White, P. Basic Procedures. In Fmoc Solid Phase Peptide Synthesis. A Practical Approach; Chan, W.C., White, P., Eds.; Oxford University Press: Oxford, UK, 2000; Volume 1, pp. 41–76. [Google Scholar]
- Zhang, J.; Robinson, C. Novel method for the purification of house dust mite allergen Der p 1 and its use in structure-based chemical design of novel inhibitors. In Allergy: Methods and Protocols, 2nd ed.; Lympany, P., Jones, M.G., Eds.; Springer Science & Business Media: New York, NY, USA, 2019; in press. [Google Scholar]
- Tian, W.X.; Tsou, C.L. Determination of the rate constant of enzyme modification by measuring the substrate reaction in the presence of the modifier. Biochemistry 1982, 21, 1028–1032. [Google Scholar] [CrossRef] [PubMed]
- Winton, H.L.; Wan, H.; Cannell, M.B.; Gruenert, D.C.; Thompson, P.J.; Garrod, D.R.; Stewart, G.A.; Robinson, C. Cell lines of pulmonary and non-pulmonary origin as tools to study the effects of house dust mite proteinases on the regulation of epithelial permeability. Clin. Exp. Allergy 1998, 28, 1273–1285. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Km (µM) |
|---|---|
| Der p 1 | 15.4 ± 2.1 |
| Der f 1 | 29.0 ± 2.6 |
| Cathepsin B | 40.9 ± 4.5 |
| Cathepsin L | 8.0 ± 1.3 |
| Cathepsin S | 12.3 ± 1.8 |
| Trypsin | 46.6 ± 2.9 |
| Chymotrypsin | 132.1 ± 22.7 |
| Enzyme | Kobs/[I] (M−1 s−1) |
|---|---|
| Der p 1 | 2 × 108 |
| Der f 1 | 1.9 × 108 |
| Cathepsin B | 3.7 × 107 |
| Cathepsin S | 2.1 × 107 |
| Cathepsin L | 1.7 × 106 |
| Cathepsin H | 1.5 × 106 |
| Cathepsin K | 1.6 × 104 |
| Thrombin | inactive at ≤50 µM |
| Trypsin | inactive at ≤50 µM |
| Chymotrypsin | inactive at ≤50 µM |
| TJ Loop and Identified Cleavage | S4 | S3 | S2 | S1 | S1′ | S2′ | S3′ | S4′ |
|---|---|---|---|---|---|---|---|---|
| CLD1.1 | ||||||||
| SLLN.LSST | Ser | Leu | Leu | Asn | Leu | Ser | Ser | Thr |
| FDSL.LNLS | Phe | Asp | Ser | Leu | Leu | Asn | Leu | Ser |
| LSST.LQAT | Leu | Ser | Ser | Thr | Leu | Gln | Ala | Thr |
| IVTA.QAMY | Ile | Val | Thr | Ala | Gln | Ala | Met | Tyr |
| IQCK.VFDS | Ile | Gln | Cys | Lys | Val | Phe | Asp | Ser |
| RIYS.YAGD | Arg | Ile | Tyr | Ser | Tyr | Ala | Gly | Asp |
| CVSQ.STGQ | Cys | Val | Ser | Gln | Ser | Thr | Gly | Gln |
| VTAQ.AMYE | Val | Thr | Ala | Gln | Ala | Met | Tyr | Glu |
| Occludin 1.1 | ||||||||
| GTSL.LGGS | Gly | Thr | Ser | Leu | Leu | Gly | Gly | Ser |
| CL | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 25 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 100 | 43 | 61 | 62 | 57 | 60 | 70 | 51 | 60 | 37 | 29 | 24 | 49 | 38 | 26 | 42 | 44 | 64 | 38 | 34 | 34 | 25 | 30 |
| 2 | 43 | 100 | 45 | 45 | 42 | 49 | 47 | 40 | 49 | 35 | 29 | 22 | 58 | 30 | 26 | 36 | 42 | 55 | 40 | 24 | 24 | 19 | 26 |
| 3 | 61 | 45 | 100 | 94 | 67 | 80 | 59 | 71 | 80 | 44 | 32 | 25 | 61 | 42 | 31 | 63 | 42 | 57 | 45 | 40 | 40 | 24 | 31 |
| 4 | 62 | 45 | 94 | 100 | 74 | 85 | 55 | 68 | 83 | 42 | 35 | 28 | 62 | 40 | 34 | 58 | 46 | 53 | 43 | 38 | 38 | 25 | 34 |
| 5 | 57 | 42 | 67 | 74 | 100 | 72 | 60 | 57 | 70 | 40 | 37 | 24 | 53 | 36 | 34 | 43 | 40 | 49 | 45 | 36 | 36 | 25 | 30 |
| 6 | 60 | 49 | 80 | 85 | 72 | 100 | 53 | 66 | 98 | 40 | 35 | 26 | 59 | 32 | 32 | 53 | 42 | 57 | 43 | 40 | 40 | 23 | 34 |
| 7 | 70 | 47 | 59 | 55 | 60 | 53 | 100 | 49 | 53 | 42 | 33 | 26 | 49 | 38 | 32 | 43 | 42 | 60 | 45 | 36 | 36 | 25 | 28 |
| 8 | 51 | 40 | 71 | 68 | 57 | 66 | 49 | 100 | 66 | 33 | 31 | 28 | 49 | 32 | 34 | 70 | 37 | 45 | 32 | 40 | 40 | 25 | 32 |
| 9 | 60 | 49 | 80 | 83 | 70 | 98 | 53 | 66 | 100 | 40 | 35 | 26 | 58 | 32 | 32 | 53 | 40 | 57 | 43 | 40 | 40 | 23 | 34 |
| 10 | 37 | 35 | 44 | 42 | 40 | 40 | 42 | 33 | 40 | 100 | 35 | 18 | 37 | 64 | 30 | 31 | 29 | 42 | 37 | 30 | 29 | 13 | 28 |
| 11 | 29 | 29 | 32 | 35 | 37 | 35 | 33 | 31 | 35 | 35 | 100 | 17 | 37 | 24 | 27 | 27 | 33 | 32 | 38 | 27 | 26 | 19 | 27 |
| 12 | 24 | 22 | 25 | 28 | 24 | 26 | 26 | 28 | 26 | 18 | 17 | 100 | 22 | 23 | 27 | 20 | 27 | 24 | 22 | 21 | 21 | 24 | 19 |
| 14 | 49 | 58 | 61 | 62 | 53 | 59 | 49 | 49 | 58 | 37 | 37 | 22 | 100 | 34 | 34 | 45 | 38 | 47 | 47 | 32 | 32 | 23 | 30 |
| 15 | 38 | 30 | 42 | 40 | 36 | 32 | 38 | 32 | 32 | 64 | 24 | 23 | 34 | 100 | 27 | 34 | 28 | 38 | 32 | 22 | 22 | 16 | 20 |
| 16 | 26 | 26 | 31 | 34 | 34 | 32 | 32 | 34 | 32 | 30 | 27 | 27 | 34 | 27 | 100 | 30 | 25 | 28 | 26 | 32 | 31 | 19 | 26 |
| 17 | 42 | 36 | 63 | 58 | 43 | 53 | 43 | 70 | 53 | 31 | 27 | 20 | 45 | 34 | 30 | 100 | 31 | 47 | 36 | 36 | 36 | 25 | 32 |
| 18 | 44 | 42 | 42 | 46 | 40 | 42 | 42 | 37 | 40 | 29 | 33 | 27 | 38 | 28 | 25 | 31 | 100 | 38 | 35 | 28 | 27 | 29 | 28 |
| 19 | 64 | 55 | 57 | 53 | 49 | 57 | 60 | 45 | 57 | 42 | 32 | 24 | 47 | 38 | 28 | 47 | 38 | 100 | 43 | 32 | 32 | 27 | 28 |
| 20 | 38 | 40 | 45 | 43 | 45 | 43 | 45 | 32 | 43 | 37 | 38 | 22 | 47 | 32 | 26 | 36 | 35 | 43 | 100 | 24 | 24 | 21 | 28 |
| 21 | 34 | 24 | 40 | 38 | 36 | 40 | 36 | 40 | 40 | 30 | 27 | 21 | 32 | 22 | 32 | 36 | 28 | 32 | 24 | 100 | 100 | 24 | 62 |
| 22 | 34 | 24 | 40 | 38 | 36 | 40 | 36 | 40 | 40 | 29 | 26 | 21 | 32 | 22 | 31 | 36 | 27 | 32 | 24 | 100 | 100 | 25 | 62 |
| 23 | 25 | 19 | 24 | 25 | 25 | 23 | 25 | 25 | 23 | 13 | 19 | 24 | 23 | 16 | 19 | 25 | 29 | 27 | 21 | 24 | 25 | 100 | 18 |
| 25 | 30 | 26 | 31 | 34 | 30 | 34 | 28 | 32 | 34 | 28 | 27 | 19 | 30 | 20 | 26 | 32 | 28 | 28 | 28 | 62 | 62 | 18 | 100 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Chen, J.; Zuo, J.; Newton, G.K.; Stewart, M.R.; Perrior, T.R.; Garrod, D.R.; Robinson, C. Allergen Delivery Inhibitors: Characterisation of Potent and Selective Inhibitors of Der p 1 and Their Attenuation of Airway Responses to House Dust Mite Allergens. Int. J. Mol. Sci. 2018, 19, 3166. https://doi.org/10.3390/ijms19103166
Zhang J, Chen J, Zuo J, Newton GK, Stewart MR, Perrior TR, Garrod DR, Robinson C. Allergen Delivery Inhibitors: Characterisation of Potent and Selective Inhibitors of Der p 1 and Their Attenuation of Airway Responses to House Dust Mite Allergens. International Journal of Molecular Sciences. 2018; 19(10):3166. https://doi.org/10.3390/ijms19103166
Chicago/Turabian StyleZhang, Jihui, Jie Chen, Jie Zuo, Gary K. Newton, Mark R. Stewart, Trevor R. Perrior, David R. Garrod, and Clive Robinson. 2018. "Allergen Delivery Inhibitors: Characterisation of Potent and Selective Inhibitors of Der p 1 and Their Attenuation of Airway Responses to House Dust Mite Allergens" International Journal of Molecular Sciences 19, no. 10: 3166. https://doi.org/10.3390/ijms19103166
APA StyleZhang, J., Chen, J., Zuo, J., Newton, G. K., Stewart, M. R., Perrior, T. R., Garrod, D. R., & Robinson, C. (2018). Allergen Delivery Inhibitors: Characterisation of Potent and Selective Inhibitors of Der p 1 and Their Attenuation of Airway Responses to House Dust Mite Allergens. International Journal of Molecular Sciences, 19(10), 3166. https://doi.org/10.3390/ijms19103166