iTRAQ-Based Quantitative Proteome Revealed Metabolic Changes in Winter Turnip Rape (Brassica rapa L.) under Cold Stress

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
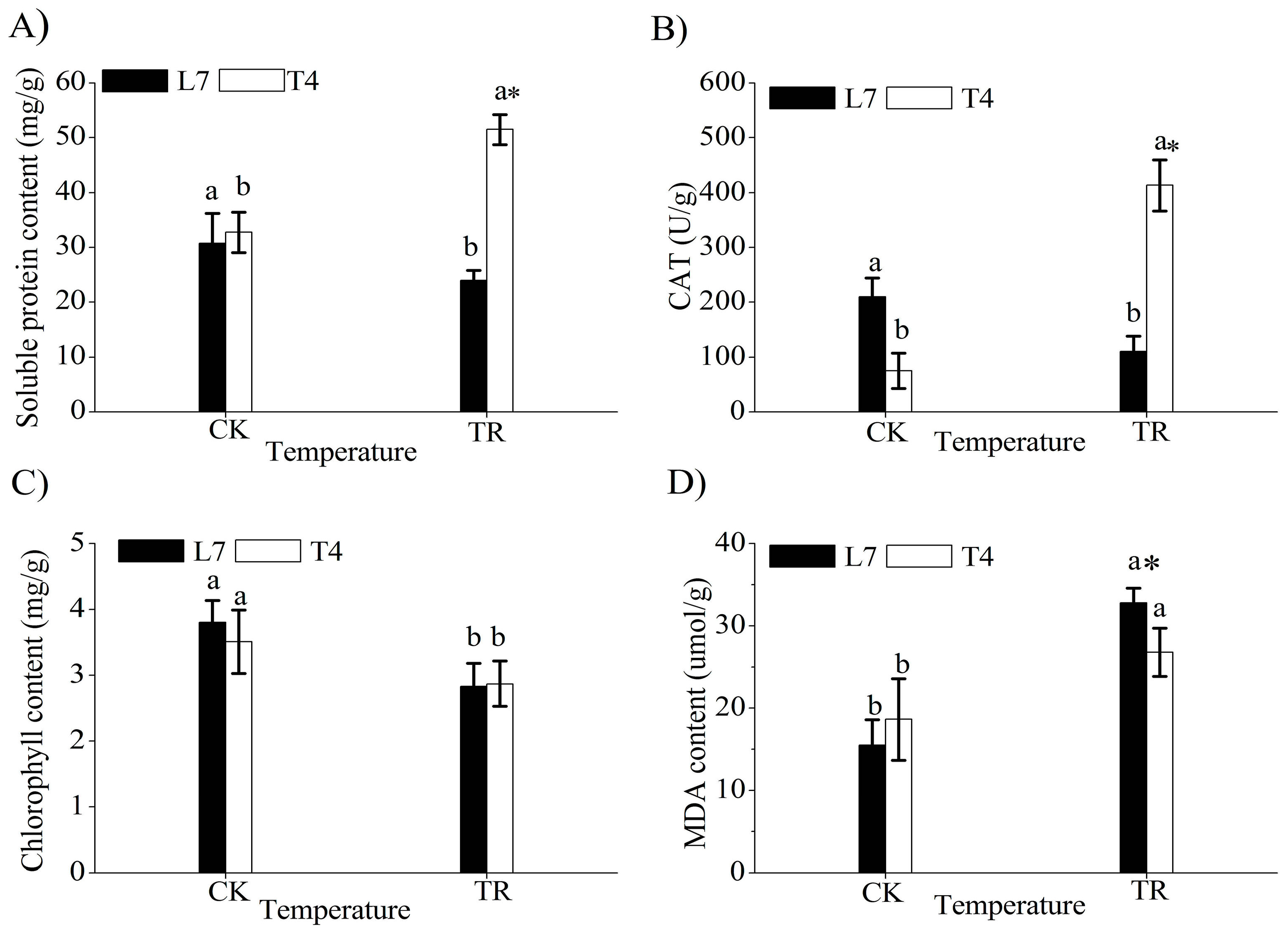
2.1. Analysis of Plant Growth and Physiological Indices under Cold Stress
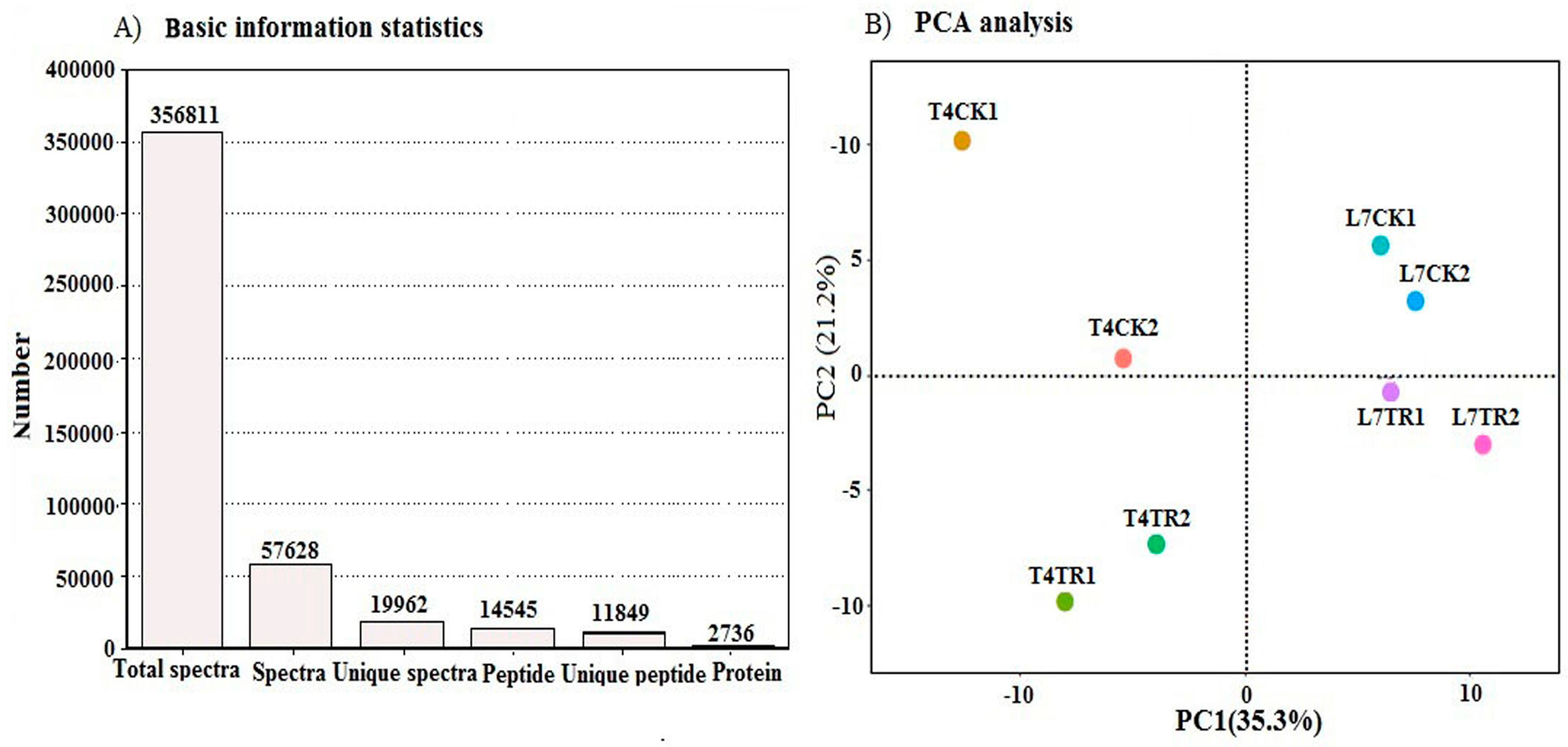
2.2. Protein Identification and Quantification
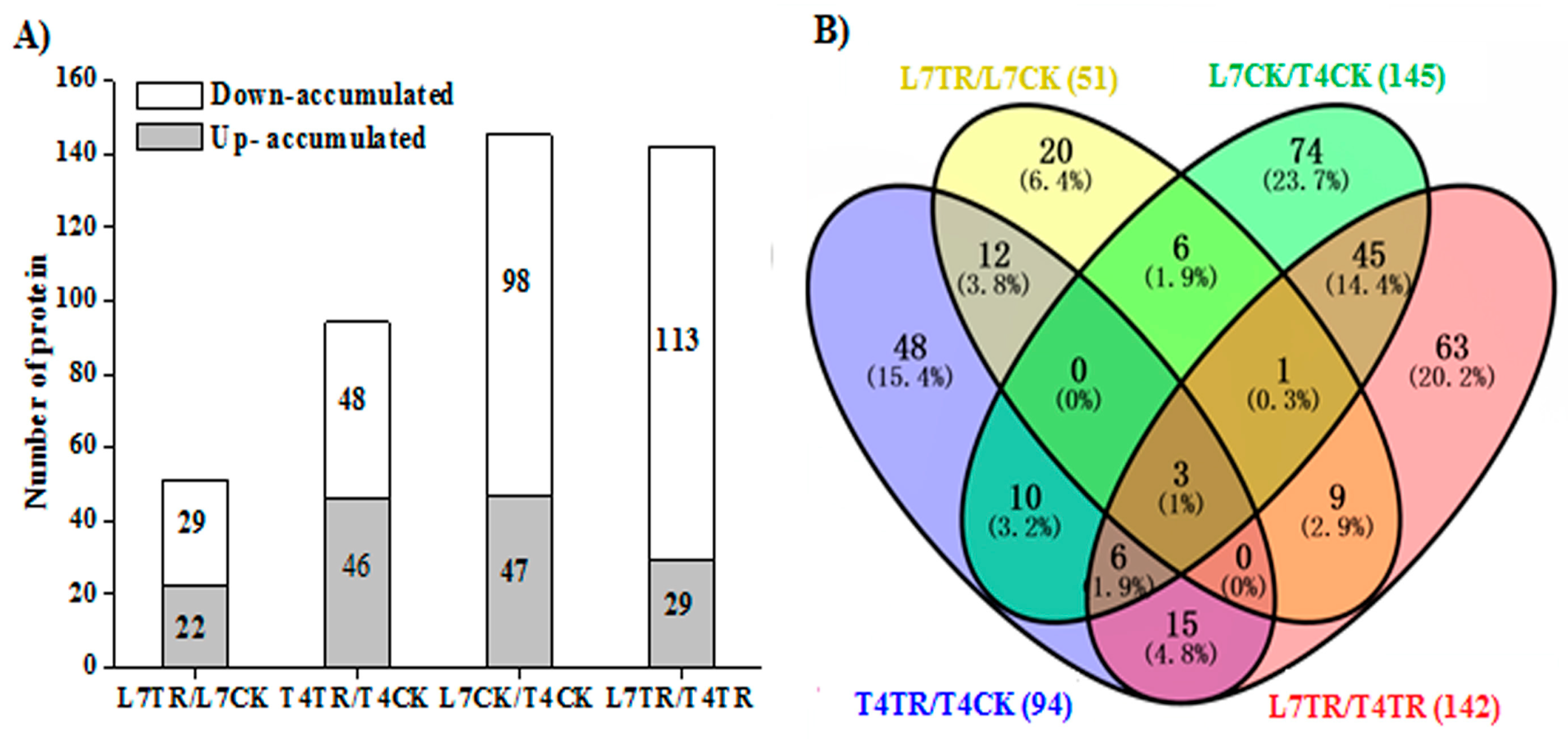
2.3. Identification and Analysis of Differently Accumulated Proteins (DAPs)
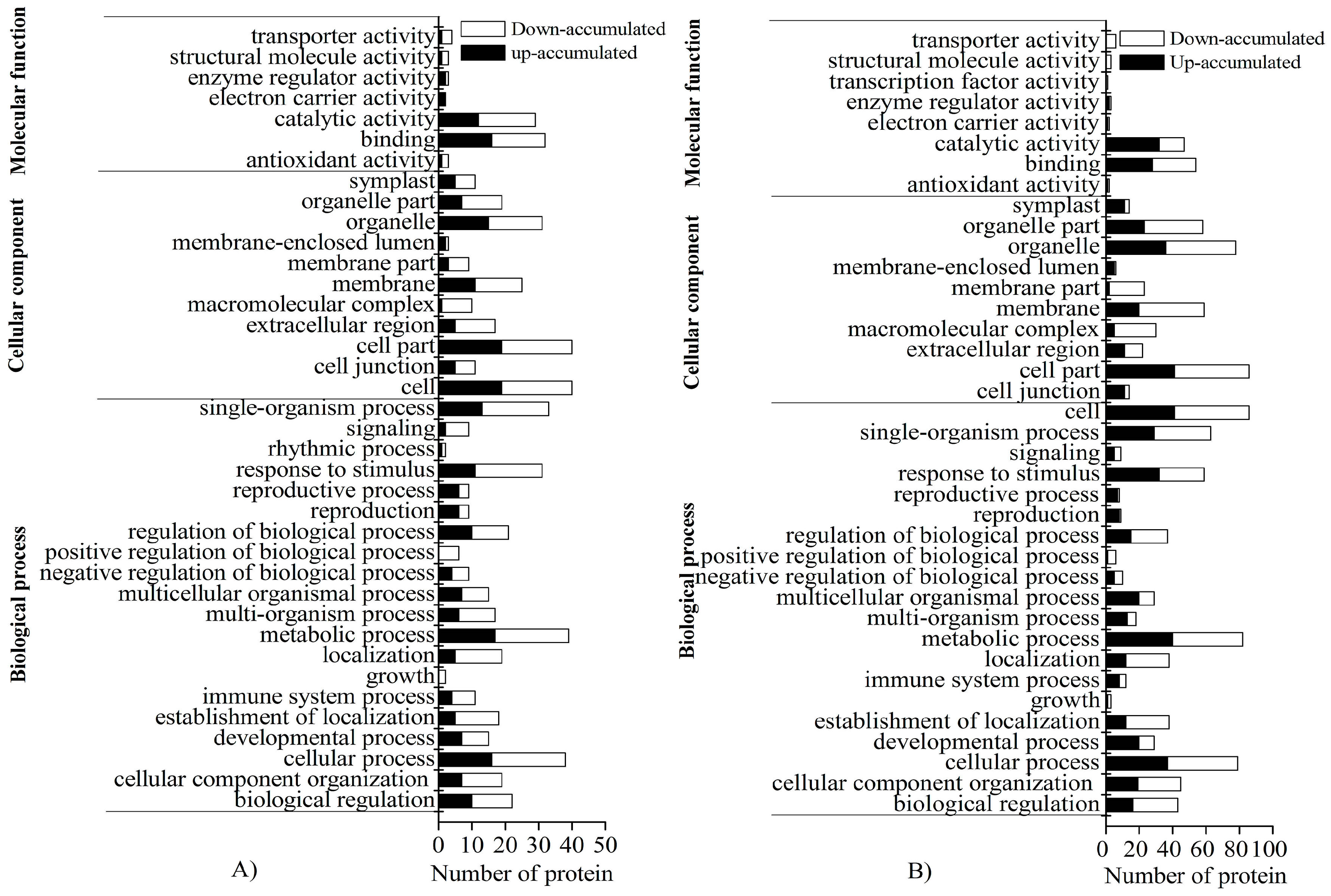
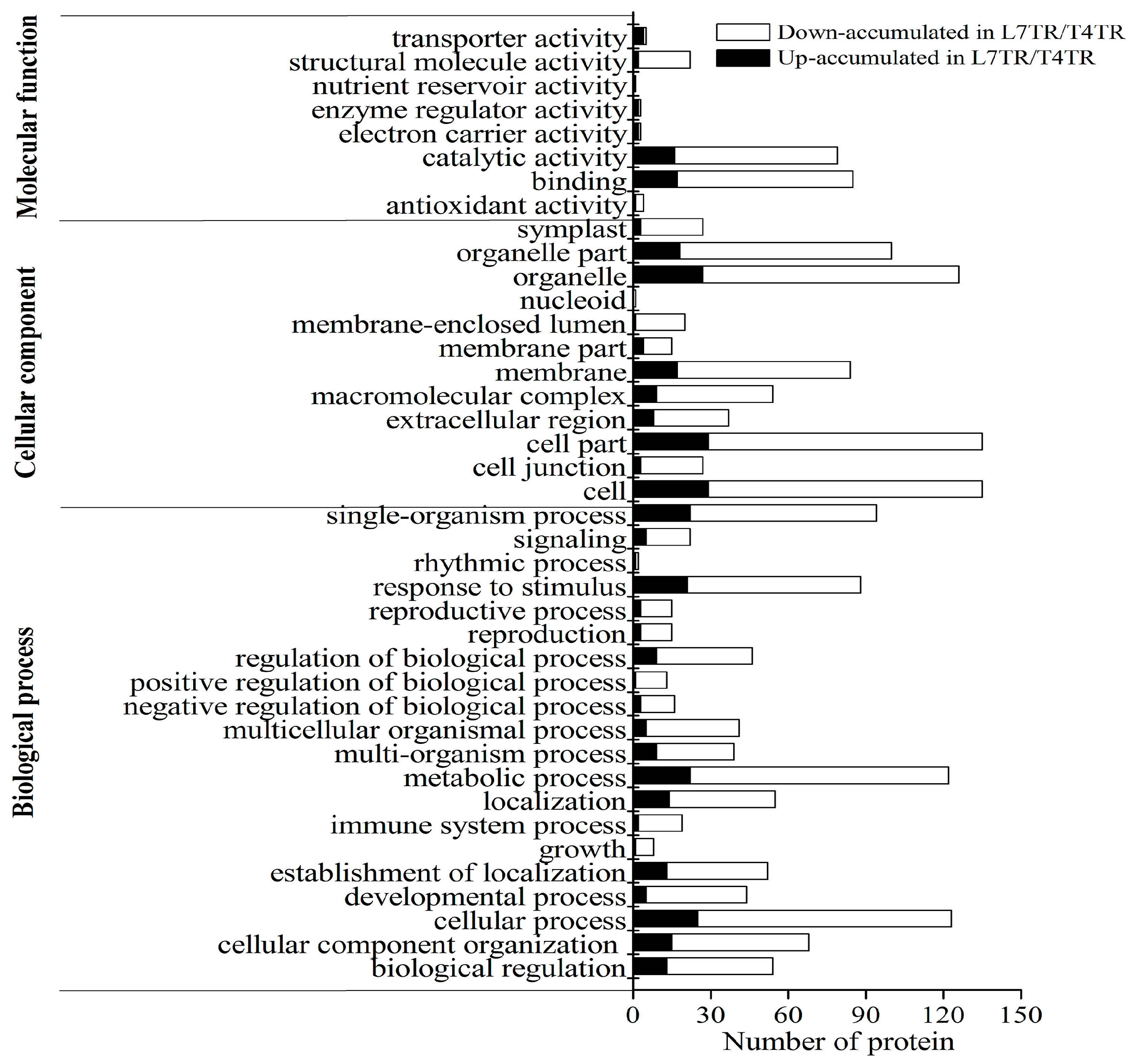
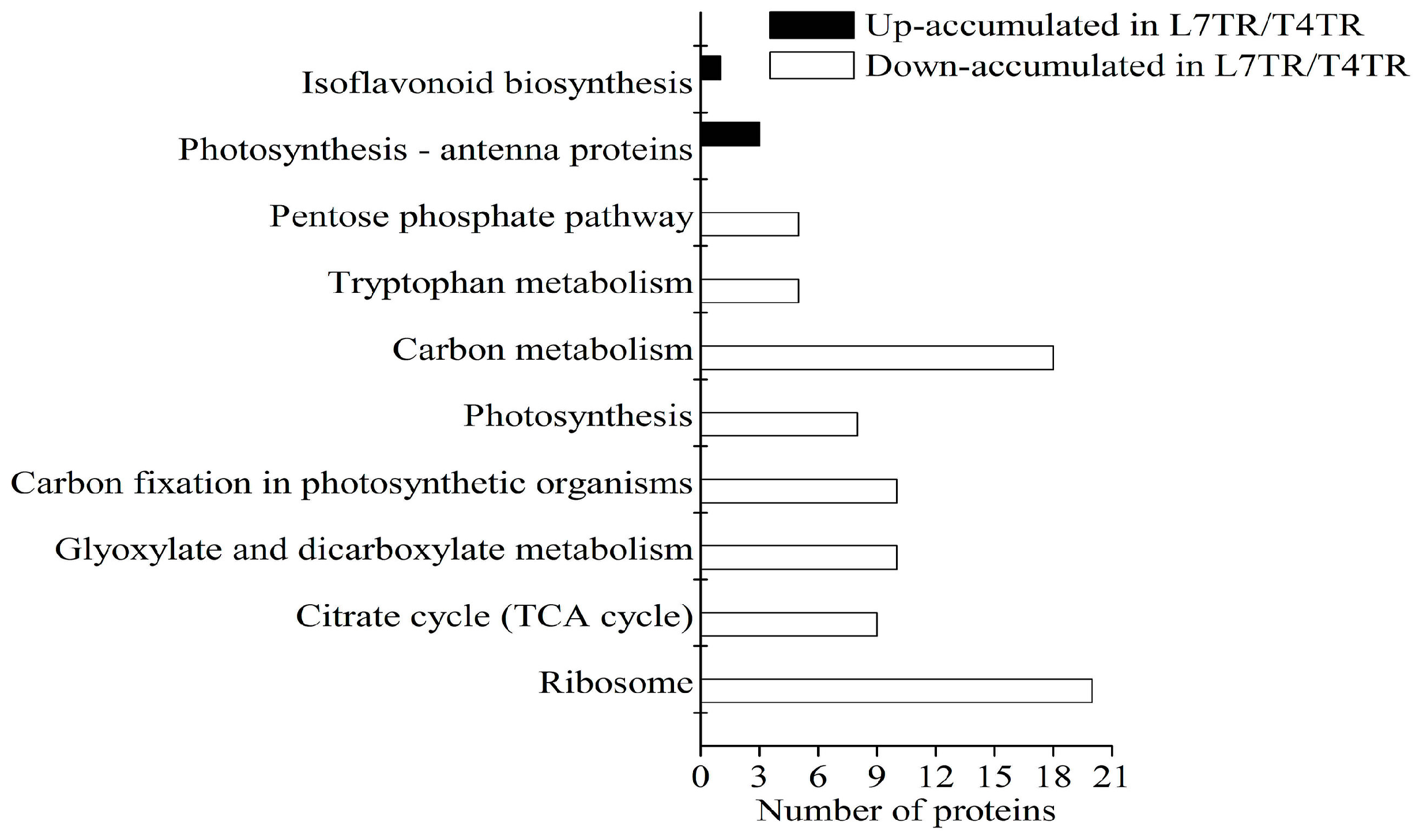
2.4. GO and KEGG Enrichment of DAPs under Cold Stress
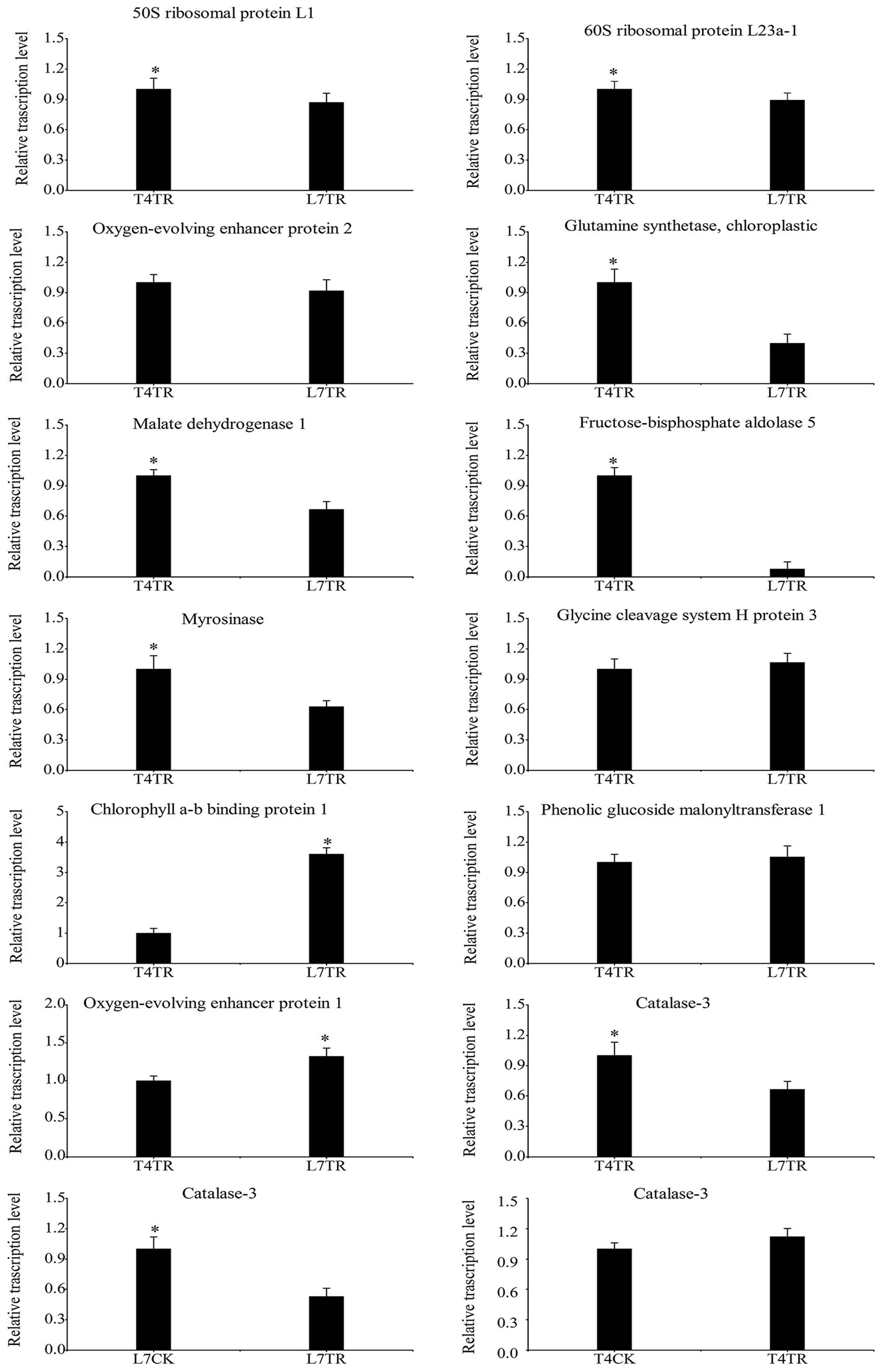
2.5. Correlation between Protein Abundance and Gene Expression by qRT-PCR
3. Discussion
3.1. Decreased Abundance of Photosynthesis-Related Proteins under Cold Stress
3.2. Decreased Abundance of Energy-Metabolism-Related Proteins under Cold Stress
3.3. Decreased Abundance of Carbon-Metabolism-Related Proteins under Cold Stress
3.4. Decreased Abundance of Ribosome-Related Proteins under Cold Stress
3.5. Other Metabolism-Related Proteins under Cold Stress
4. Materials and Methods
4.1. Plant Materials and Cold Stress Treatment
4.2. Analysis of Physiological and Biochemical Indices
4.3. Protein Extraction and iTRAQ Labeling
4.4. Separation of Peptides and LC–MS/MS Analysis
4.5. iTRAQ Protein Identification and Quantification
4.6. RNA Extraction and qPCR Analysis of Gene Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| iTRAQ | Isobaric tags for relative and absolute quantification |
| 2DE | Two-dimensional electrophoresis |
| CAT | Catalase |
| MDA | Malondialdehyde |
| PCA | Principal component analysis |
| TEAB | Triethylamonium bicarbonate |
| CK | Control |
| DAPs | Differently accumulated proteins |
| GO | Gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| ROS | Reactive oxygen species |
| OEE | Oxygen-evolving enhancer protein |
| PSII | Photosystem II |
| PSI | Photosystem I |
| TCA | Citrate cycle |
| PPP | Pentose phosphate pathway |
| NADP-ICDH | Accompanying cytosolic isocitrate dehydrogenase (NADP) |
| ACO | Aconitate hydratase, mitochondrial |
| SUCLA | Succinate-CoA ligase (ADP-forming)subunit beta |
| MDH | Malatedehydrogenase |
| TKT | Transketolase |
| FBPA | Fructose-bisphosphate aldolase |
| RPE | Ribulose-phosphate 3-epimerase |
| GS | Glutamine synthetase |
| GCSH | Glycine cleavage system H protein |
| GCS | Glycine cleavage system |
| CAT3 | Catalase3 |
| MDH | Malate dehydrogenase |
| OAA | Oxaloacetate |
| MYR | Myrosinase |
| PMAT | Phenolic glucosidemalonyl transferase |
References
- Wang, Q.W.; Guan, X.L.; Hu, Z.H.; Lu, C.F.; Leng, P.S. Relationship between cold tolerance and leaf structure of the three species of sedum. Chin. J. Appl. Environ. Biol. 2013, 19, 280–285. [Google Scholar] [CrossRef]
- Walker, D.J.; Romero, P.; Hoyos, A.D.; Correal, E. Seasonal changes in cold tolerance, water relations and accumulation of cations and compatible solutes in Atriplex halimus L. Environ. Exp. Bot. 2008, 64, 217–224. [Google Scholar] [CrossRef]
- An, D.; Yang, J.; Zhang, P. Transcriptome profiling of low temperature-treated cassava apical shoots showed dynamic responses of tropical plant to cold stress. BMC Genomics 2012, 13, 64. [Google Scholar] [CrossRef] [PubMed]
- Medina, J.; Catalá, R.; Salinas, J. Developmental and stress regulation of RCI2A and RCI2B, two cold-inducible genes of Arabidopsis encoding highly conserved hydrophobic proteins. Plant Physiol. 2001, 125, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Qin, W. Turf plant cell damage by low temperatures. Pratacult. Sci. 1993, 10, 57–61. [Google Scholar]
- Kreps, J.A.; Wu, Y.; Chang, H.S.; Zhu, T.; Wang, X.; Harper, J.F. Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress. Plant Physiol. 2002, 130, 2129–2141. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.; Thomashow, M.F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, M.A.; Maruyama, K.; Abe, H.; Khan, M.A.; Katsura, K.; Ito, Y.; Yoshiwara, K.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Monitoring expression profiles of rice genes under cold, drought, and high-salinity stresses and abscisic acid application using cDNA microarray and RNA gel-blot analyses. Plant Physiol. 2003, 133, 1755–1767. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Takeda, M.; Kidokoro, S.; Yamada, K.; Sakuma, Y.; Urano, K.; Fujita, M.; Yoshiwara, K.; Matsukura, S.; Morishita, Y. Metabolic pathways involved in cold acclimation identified by integrated analysis of metabolites and transcripts regulated by DREB1A and DREB2A. Plant Physiol. 2009, 150, 1972–1980. [Google Scholar] [CrossRef] [PubMed]
- Pradetbalade, B.; Boulmé, F.; Beug, H.; Müllner, E.W.; Garciasanz, J.A. Translation control: Bridging the gap between genomics and proteomics? Trends Biochem. Sci. 2001, 26, 225–229. [Google Scholar] [CrossRef]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.M.; Zhao, T.T.; Gan, S.H.; Ren, X.D.; Fang, L.C.; Karungo, S.K.; Wang, Y.; Chen, L.; Li, S.H.; Xin, H.P. Ethylene positively regulates cold tolerance in grapevine by modulating the expression of ETHYLENE RESPONSE FACTOR 057. Sci. Rep. 2016, 6, 24066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.K. Proteomics and a future generation of plant molecular biologists. Plant Mol. Biol. 2002, 48, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Casado-Vela, J.; Martinez-Esteso, M.J.; Rodríguez, E.; Borras, E.; Elortza, F.; Bru-Martinez, R. iTRAQ-based quantitative analysis of protein mixtures withlarge fold change and dynamic range. Proteomics 2010, 10, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Men, J.L.; Chang, M.C.; Feng, C.P.; Yuan, L.G. iTRAQ-based quantitative proteome revealed metabolic changes of Flammulina velutipes mycelia in response to cold stress. J. Proteom. 2017, 156, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.G.; Liu, A.; Bi, A.; Amombo, E.; Gitau, M.M.; Huang, X.B.; Chen, L.; Fu, J.M. Identification of differentially expressed proteins in bermudagrass response to cold stress in the presence of ethylene. Environ. Exp. Bot. 2017, 139, 67–78. [Google Scholar] [CrossRef]
- Wang, X.; Shan, X.; Wu, Y.; Su, S.; Li, S.; Liu, H.; Han, J.; Xue, C.; Yuan, Y. ITRAQ-based quantitative proteomic analysis reveals new metabolic pathways responding to chilling stress in maize seedlings. J. Proteom. 2016, 146, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Yin, X.; Xiang, Y.; Xin, H.; Li, S.; Yang, P. iTRAQ-based comparative proteomic analyses of two grapevine cultivars in response to cold Stress. Curr. Proteom. 2017, 14, 42–52. [Google Scholar] [CrossRef]
- Zheng, X.; Fan, S.; Wei, H.; Tao, C.; Ma, Q.; Ma, Q.; Zhang, S.; Li, H.; Pang, C.; Yu, S. iTRAQ-based quantitative proteomic analysis reveals cold responsive proteins involved in leaf senescence in upland cotton (Gossypium hirsutum L.). Int. J. Mol. Sci. 2017, 18, 1984. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ye, C.; Wang, M.; Chu, G. Identification of cold-stress responsive proteins in Anabasis aphylla seedlings via the iTRAQ proteomics technique. J. Plant Interact. 2017, 12, 505–519. [Google Scholar] [CrossRef]
- Sun, W.C.; Wu, J.Y.; Fang, Y.; Liu, Q.; Yang, R.Y.; Ma, W.G.; Li, X.C.; Zhang, J.J.; Zhang, P.F.; Lei, J.M. Growth and development characteristics of winter rapeseed northern-extended from the cold and arid regions in China. Acta Agron. Sin. 2010, 36, 2124–2134. [Google Scholar] [CrossRef]
- Sun, W.C.; Wu, J.Y.; Zeng, J.; Zhu, H.X.; Liu, Y.L.; Zhang, Y.H. Primary evaluation of cold tolerance among eight winter Brassica rapa. J. Hunan Agric. Univ. 2007, 33, 151–155. [Google Scholar]
- Xu, Y.Z.; Zeng, X.C.; Zhang, F.Q.; Sun, J.; Sun, W.C.; Wu, J.Y.; Fang, Y.; Liu, Z.G.; Sun, B.L. Response of leaf anatomical structure and photosynthetic characteristics of winter turnip rape (Brassica rapa L.) to low temperature before winter. Acta Agron. Sin. 2017, 43, 432–441. [Google Scholar] [CrossRef]
- Liu, Z.G.; Sun, W.C.; Yang, N.N.; Wang, Y.; Li, H.E.; Zhao, C.X.; Shi, P.F.; Yang, G.; Li, X.C.; Wu, J.Y. Morphology and physiological characteristics of cultivars with different levels of cold-resistance in winter rapeseed (Brassica campestris L.) during cold acclimation. Sci. Agric. Sin. 2013, 46, 4679–4687. [Google Scholar]
- Zeng, X.C.; Xu, Y.Z.; Jiang, J.J.; Zhang, F.Q.; Ma, L.; Wu, D.W.; Wang, Y.P.; Sun, W.C. Identification of cold stress responsive microRNAs in two winter turnip rape (Brassica rapa L.) by high throughput sequencing. BMC Plant Biol. 2018, 18, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, J.; Li, Z.F.; Fu, L.S.; Sun, Y.; Wang, X.; Chen, Y.; Chen, Z. Comparison of physiological metabolism of different cold resistance cultivars during cold acclimation period. J. Northeast Agric. Univ. 2009, 40, 14–17. [Google Scholar]
- Gan, C.S.; Chong, P.K.; Pham, T.K.; Wright, P.C. Technical experimental and biological variations in isobaric tags for relative and absolute quantitation (iTRAQ). J. Proteome Res. 2007, 6, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.X. Research progress on the mechanism of improving plant cold hardiness. Acta Ecol. Sin. 2012, 32, 7966–7980. [Google Scholar]
- Wei, G.S.; Deng, S. An overview for temperature perception and cold acclimation in plants. J. Shandong Agric. Univ. 2015, 46, 827–832. [Google Scholar]
- Kosová, K.; Vítámvás, P.; Planchon, S.; Renaut, J.; Vanková, R.; Prášil, I.T. Proteome analysis of cold response in spring and winter wheat (Triticum aestivum) crowns reveals similarities in stress adaptation and differences in regulatory processes between the growth habits. J. Proteome Res. 2013, 12, 4830–4845. [Google Scholar] [CrossRef] [PubMed]
- Tanino, K.K.; McKersie, B.D. Injury within the crown of winter wheat seedlings after freezing and icing stress. Can. J. Bot. 1984, 63, 432–436. [Google Scholar] [CrossRef]
- Gołębiowska-Pikania, G.; Kopeć, P.; Surówka, E.; Krzewska, M.; Dubas, E.; Nowicka, A.; Rapacz, M.; Wójcik-Jagła, M.; Malaga, S.; Żur, I. Changes in protein abundance and activity involved in freezing tolerance acquisition in winter barley (Hordeum vulgare L.). J. Proteom. 2017, 169, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Mikula, P.; Zezulka, S.; Jancula, D.; Marsalek, B. Metabolic activity and membrane integrity changes in–new findings on hydrogen peroxide toxicity in cyanobacteria. Eur. J. Phycol. 2012, 47, 195–206. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Khan, M.H.; Panda, S.K. Hydrogen peroxide induces oxidative stress in detached leaves of Oryza sativa L. Plant Physiol. 2007, 33, 83–95. [Google Scholar]
- Thounaojam, T.C.; Panda, P.; Mazumdar, P.; Kumar, D.; Sharma, G.D.; Sahoo, L.; Panda, S.K. Excess copper induced oxidative stress and response of antioxidants in rice. Plant Physiol. Biochem. 2012, 53, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Misra, N.; Gupta, A.K. Effect of salinity and different nitrogen sources on the activity of antioxidant enzymes and indole alkaloid content in Catharanthus roseus seedlings. J. Plant Physiol. 2006, 163, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yi, H. Effect of sulfur dioxide on ROS production, gene expression and antioxidant enzyme activity in Arabidopsis plants. Plant Physiol. Biochem. 2012, 58, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hu, C.; Tan, Q.; Nie, Z.; Sun, X. Effects of molybdenum on water utilization, antioxidative defense system and osmotic-adjustment ability in winter wheat (Triticum aestivum) under drought stress. Plant Physiol. Biochem. 2014, 83, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, M.; Thomashow, M. Plant cold acclimation identifying gene regulons involved in freezing tolerance. In Vitro Cell. Dev. Biol.-Anim. 2010, 46, S78–S79. [Google Scholar]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C (3) plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, K.; Hanagata, N.; Dubinsky, Z.; Baba, S.; Karube, I. Molecular characterization of cDNA encoding oxygen evolving enhancer protein 1 increased by salt treatment in the mangrove Bruguiera gymnorrhiza. Plant Cell Physiol. 2000, 41, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Mizobuchi, A.; Yamamoto, Y. Assembly of photosystem II polypeptides and expression of oxygen evolution activity in the chloroplasts of Euglena gracilis Z during the dark-light transition. Biochim. Biophys. Acta Bioenerg. 1989, 977, 26–32. [Google Scholar] [CrossRef]
- Rocco, M.; Arena, S.; Renzone, G.; Scippa, G.S.; Lomaglio, T.; Verrillo, F.; Scaloni, A.; Marra, M. Proteomic analysis of temperature stress-responsive proteins in Arabidopsis thaliana rosette leaves. Mol. Biosyst. 2013, 9, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Gharechahi, J.; Alizadeh, H.; Naghavi, M.R.; Sharifi, G. A proteomic analysis to identify cold acclimation associated proteins in wild wheat (Triticum urartu L.). Mol. Biol. Rep. 2014, 41, 3897–3905. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Han, G.; Shang, C.; Li, J.; Zhang, H.; Liu, F.; Zhang, Y. Proteomic analyses reveal differences in cold acclimation mechanisms in freezing-tolerant and freezingsensitive cultivars of alfalfa. Front. Plant Sci. 2015, 6, 105. [Google Scholar] [CrossRef] [PubMed]
- Kirchgeßner, H.D.; Reichert, K.; Hauff, K.; Steinbrecher, R.; Schnitzler, J.P.; Pfündel, E.E. Light and temperature, but not UV radiation, affect chlorophylls and carotenoids in Norway spruce needles (Piceaabies (L.) Karst.). Plant Cell Environ. 2003, 26, 1169–1179. [Google Scholar] [CrossRef]
- Milne-Redhead, E. Plant respiration. Kew Bull. 1965, 19, 198. [Google Scholar] [CrossRef]
- Wan, Q.L.; Liu, M.Y. A preliminary study on cold resistant physiology of Ruta Graveolens. Bull. Bot. Res. 1997, 17, 190–194. [Google Scholar]
- Wan, A.; Liu, M.Y. Study on annual growth rhythm and physiological character of mature plants of Qdonis amurensis. Bull. Bot. Res. 1996, 16, 351–355. [Google Scholar]
- Wang, D.L.; Ye, W.W.; Wang, J.J.; Song, L.Y.; Fan, W.L.; Cui, Y.P. Construction of SSH library and its analyses of cotton drought associated genes under drought stress. Acta Agron. Sin. 2010, 36, 2035–2044. [Google Scholar] [CrossRef]
- Hoshida, H.; Tanaka, Y.; Hibino, T.; Hayashi, Y.; Tanaka, A.; Takabe, T.; Takabe, T. Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast glutamine synthetase. Plant Mol. Biol. 2000, 43, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Hasse, D.; Mikkat, S.; Hagemann, M.; Bauwe, H. Alternative splicing produces an H-protein with better substrate properties for the P-protein of glycine decarboxylase. FEBS J. 2010, 276, 6985–6991. [Google Scholar] [CrossRef] [PubMed]
- Häusler, R.E.; Lea, P.J.; Leegood, R.C. Control of photosynthesis in barley leaves with reduced activities of glutamine synthetase or glutamate synthase. Planta 1994, 200, 388–396. [Google Scholar]
- Wallsgrove, R.M.; Turner, J.C.; Hall, N.P.; Kendall, A.C.; Bright, S.W.J. Barley mutants lacking chloroplast glutamine synthetase-biochemical and genetic analysis. Plant Physiol. 1987, 83, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Kozaki, A.; Takeba, G. Photorespiration protects C3 plants from photooxidation. Nature 1996, 384, 557–560. [Google Scholar] [CrossRef]
- Timm, S.; Florian, A.; Arrivault, S.; Stitt, M.; Fernie, A.R.; Bauwe, H. Glycine decarboxylase controls photosynthesis and plant growth. FEBS Lett. 2012, 586, 3692–3697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Die, J.V.; Arora, R.; Rowland, L.J. Proteome dynamics of cold-acclimating Rhododendron species contrasting in their freezing tolerance and thermonasty behavior. PLoS ONE 2017, 12, e0177389. [Google Scholar] [CrossRef] [PubMed]
- Imin, N.; Kerim, T.; Weinman, J.J.; Rolfe, B.G. Low temperature treatment at the young microspore stage induces protein changes in rice anthers. Mol. Cell. Proteom. 2006, 5, 274–292. [Google Scholar] [CrossRef] [PubMed]
- Lv, G.Y.; Guo, X.G.; Xie, L.P.; Xie, C.G.; Zhang, X.H.; Yang, Y.; Xiao, L.; Tang, Y.Y.; Pan, X.L.; Guo, A.G. Molecular characterization, gene evolution, and expression analysis of the fructose-1,6-bisphosphate aldolase (FBA) gene family in wheat (Triticum aestivum L.). Front. Plant Sci. 2017, 8, 1030. [Google Scholar] [CrossRef] [PubMed]
- Heerden, P.D.R.V.; Krüger, G.H.J.; Loveland, J.E.; Parry, M.A.J.; Foyer, C.H. Dark chilling imposes metabolic restrictions on photosynthesis in soybean. Plant Cell Environ. 2010, 26, 323–337. [Google Scholar] [CrossRef]
- Zeng, Y.; Yu, J.; Cang, J.; Liu, L.; Mu, Y.; Wang, J.; Zhang, D. Detection of sugar accumulation and expression levels of correlative key enzymes in winter wheat (Triticum aestivum) at low temperatures. Biosci. Biotechnol. Biochem. 2011, 75, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Prasad, T.K. Role of catalase in inducing chilling tolerance in pre-emergent maize seedlings. Plant Physiol. 1997, 114, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Prasad, T.K.; Anderson, M.D.; Stewart, C.R. Acclimation, hydrogen peroxide, and abscisicacid protect mitochondria against irreversible chilling injury in maize seedlings. Plant Physiol. 1994, 105, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Prasad, T.K.; Anderson, M.D.; Stewart, C.R. Localization and characterization of peroxidases in the mitochondria of chilling-acclimated maize seedlings. Plant Physiol. 1995, 108, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.M.; Zhang, H.Y.; Tan, Z.B.; Huang, Y.B. Molecular cloning and chilling expression of maize Catalase-3 Gene. J. Sichuan Agric. Univ. 2006, 24, 272–275. [Google Scholar]
- Roupakias, D.G.; Mcmillin, D.E.; Scandalios, J.G. Chromosomal location of the catalase structural genes in Zea mays using B-A translocations. Theor. Appl. Genet. 1980, 58, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Redinbaugh, M.G.; Sabre, M.; Scandalios, J.G. The distribution of catalase activity, isozyme protein, and transcript in the tissues of the developing maize seedling. Plant Physiol. 1990, 92, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Carrari, F.; Lytovchenko, A.; Smith, A.; Loureiro, M.; Ratcliffe, R.; Sweetlove, L.; Fernie, A. Enhanced photosynthetic performance and growth as a consequence of decreasing mitochondrial malate dehydrogenase activity in transgenic tomato plants. Plant Physiol. 2005, 137, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Hao, Y.; Ming, L.I.; Pang, M.; Liu, Z.; Zhai, H. Gene clone, expression and enzyme activity assay of a cytosolic malate dehydrogenase from apple fruits. Acta Hortic. Sin. 2008, 2, 307–313. [Google Scholar] [CrossRef]
- Wang, Q.J.; Sun, H.; Dong, Q.L.; Sun, T.Y.; Jin, Z.X.; Hao, Y.J.; Yao, Y.X. The enhancement of tolerance to salt and cold stresses by modifying the redox state and salicylic acid content via the cytosolic malate dehydrogenase gene in transgenic apple plants. Plant Biotechnol. J. 2016, 14, 1986–1997. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Zang, N.; Zhang, J.; Zhang, H.; Li, Y.; Liu, Y.; Feng, W.; Liang, X. New insights into the mechanisms of acetic acid resistance in Acetobacter pasteurianus using iTRAQ-dependent quantitative proteomic analysis. Int. J. Food Microbiol. 2016, 238, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Deng, G.; Su, Y.; Liu, J.J.; Yang, Y.; Du, G.H.; Chen, Z.Y.; Liu, F.H. Protein mechanisms in response to NaCl-stress of salt-tolerant and salt-sensitive industrial hemp based on iTRAQ technology. Ind. Crop. Prod. 2016, 83, 444–452. [Google Scholar] [CrossRef]
- Xu, W.J.; Chen, Y.Z.; Yan, X.F. The role of glucosinolate-myrosinase system in plant defence, growth and development. Plant Physiol. Commun. 2008, 44, 1189–1196. [Google Scholar]
- Wojakowska, A.; Piasecka, A.; García-López, P.M.; Zamora-Natera, F.; Krajewski, P.; Marczak, Ł.; Kachlicki, P.; Stobiecki, M. Structural analysis and profiling of phenolic secondary metabolites of Mexican lupine species using LC–MS techniques. Phytochemistry 2013, 92, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Posmyk, M.M.; Bailly, C.; Szafrańska, K.; Janas, K.M.; Corbineau, F. Antioxidant enzymes and isoflavonoids in chilled soybean (Glycine max (L.) Merr.) seedlings. J. Plant Physiol. 2005, 162, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Dhaubhadel, S. Regulation of isoflavonoid biosynthesis in soybean seeds. Soybean-Biochem. Chem. Physiol. 2008, 50, 660–661. [Google Scholar]
- Chen, X.Q. Key enzymes in soybean isoflavones biosynthesis and its metabolic engineering. China Biotechnol. 2012, 32, 133–138. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein using the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Campos, P.S.; Quartin, V.N.; Ramalho, J.C.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.T.; Qi, Y.P.; Lu, Y.B.; Guo, P.; Sang, W.; Feng, H.; Zhang, H.X.; Chen, L.S. iTRAQ protein profile analysis of Citrus sinensis roots in response to long-term boron-deficiency. J. Proteom. 2013, 93, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Patterson, J.; Ford, K.; Cassin, A.; Natera, S.; Bacic, A. Increased abundance of proteins involved in phytosiderophore production in boron-tolerant barley. Plant Physiol. 2007, 144, 1612–1631. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pang, C.; Wei, H.L.; Song, M.Z.; Meng, Y.Y.; Ma, J.H.; Fan, S.L.; Yu, S.X. Data for proteomic profiling of anthers from a photosensitive male sterile mutant and wild-type cotton (Gossypium hirsutum L.). Data Brief 2015, 4, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.; Yan, G.X.; Yang, Q.; Zhai, L.N.; Zhang, C.; Zhang, F.Q.; Guan, R.Z. ITRAQ-based quantitative proteomics analysis of Brassica napus leaves reveals pathways associated with chlorophyll deficiency. J. Proteom. 2015, 113, 244–259. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C.; Yao, L.R.; Li, B.C.; Meng, Y.X.; Ma, X.L.; Yong, L.; Si, E.; Ren, P.R.; Yang, K.; Shang, X.W.; et al. Comparative proteomic analysis of cultured suspension cells of the Halophyte halogeton glomeratus by iTRAQ provides insights into response mechanisms to salt stress. Front. Plant Sci. 2016, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Xing, S.; Tian, Z.; Gao, S.; Xie, J.; Chang, H.; Liu, G.; Luo, J.; Yin, H. Proteomic analysis of sheep primary testicular cells infected with bluetongue virus. Proteomics 2016, 16, 1499–1514. [Google Scholar] [CrossRef] [PubMed]
- The Brassica Database (BRAD). Available online: http://brassicadb.org/brad/datasets/pub/BrassicaceaeGenome/Brassica_rapa/V2.0/Scaffold2.0/BrapaV2.0_evm.out.pep.fa.gz (accessed on 8 October 2016).
- Chen, Q.Z.; Guo, W.S.; Feng, L.Z.; Ye, X.Z.; Xie, W.F.; Huang, X.P.; Liu, J.Y. Transcriptome and proteome analysis of Eucalyptus infected with Calonectria pseudoreteaudii. J. Ptoreom. 2015, 115, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, Y.Q.; Chen, Z.; Liu, Z.J.; Ren, Q.Y.; Yang, J.F.; Zhu, X.Q.; Guan, G.Q.; Liu, A.H.; Luo, J.X. Comparative proteomic and bioinformatic analysis of Theileria luwenshuni and Theileria uilenbergi. Exp. Parasitol. 2016, 166, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta DeltaC (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession (Uniprot) | Uniprot_Swissprot Description | Fold Changes a | ||
|---|---|---|---|---|
| L7TR/L7CK | T4TR/T4CK | L7TR/T4TR | ||
| Ribosome | ||||
| Q9LY66 | 50S ribosomal protein L1 | - | - | 0.348519904 |
| Q9SKX4 | 50S ribosomal protein L3-1 | - | 0.636134306 | 0.365801714 |
| Q9SLF7 | 60S acidic ribosomal protein P2-2 | - | - | 0.483258248 |
| Q8LD46 | 60S ribosomal protein L23a-1 | - | - | 0.352601546 |
| P36210 | 50S ribosomal protein L12-1 | - | - | 0.364051915 |
| P92959 | 50S ribosomal protein L24 | - | - | 0.366241266 |
| Citrate cycle (TCA cycle) | ||||
| Q9SRZ6 | Cytosolic isocitrate dehydrogenase [NADP] | - | 1.757858283 | 0.611169392 |
| Q9SIB9 | Aconitate hydratase 3 | - | - | 0.659753955 |
| O82662 | Succinate-CoA ligase [ADP-forming] | - | - | 0.633851199 |
| Glyoxylate and dicarboxylate metabolism | ||||
| Q42624 | Glutamine synthetase | - | - | 0.36059385 |
| Q9LQL0 | Glycine cleavage system H protein 3 | - | - | 0.388785817 |
| Carbon fixation in photosynthetic organisms | ||||
| Q8RWV0 | Transketolase-1 | - | 1.696959938 | 0.611169392 |
| Q9ZTP5 | Ribulose-phosphate 3-epimerase | - | - | 0.597820343 |
| Photosynthesis | ||||
| Q96334 | Oxygen-evolving enhancer protein 2 | - | - | 0.449814881 |
| P83504 | Oxygen-evolving enhancer protein 1 | - | - | 0.375329761 |
| Q8W0Y8 | Photosystem II reaction center PSB28 | - | - | 0.405032816 |
| Carbon metabolism | ||||
| Q42547 | Catalase-3 | 0.568893536 | - | 0.550541856 |
| P93819 | Malate dehydrogenase 1, cytoplasmic | - | - | 0.595674747 |
| Q9LF98 | Fructose-bisphosphate aldolase 8 | - | - | 0.405152525 |
| P25697 | Phosphoribulokinase, chloroplastic | - | - | 0.552468889 |
| Q43743 | Malate dehydrogenase 1, glyoxysomal | - | 1.672743344 | 0.579876978 |
| O65581 | Fructose-bisphosphate aldolase 5 | - | - | 0.348710843 |
| Tryptophan metabolism | ||||
| Q00326 | Myrosinase | 0.518096445 | - | 0.495250588 |
| Pentose phosphate pathway | ||||
| Q8RWV0 | Transketolase-1, chloroplastic | - | - | 0.611169392 |
| Q43157 | Ribulose-phosphate 3-epimerase | - | - | 0.553787078 |
| Photosynthesis—antenna proteins | ||||
| P27525 | Chlorophyll a-b binding protein CP24 | - | 0.574349177 | 1.601593552 |
| P04778 | Chlorophyll a-b binding protein 1 | - | - | 1.818756629 |
| Isoflavonoid biosynthesis | ||||
| Q940Z5 | Phenolic glucosidemalonyl transferase 1 | - | - | 1.519359178 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Zeng, X.; Wu, J.; Zhang, F.; Li, C.; Jiang, J.; Wang, Y.; Sun, W. iTRAQ-Based Quantitative Proteome Revealed Metabolic Changes in Winter Turnip Rape (Brassica rapa L.) under Cold Stress. Int. J. Mol. Sci. 2018, 19, 3346. https://doi.org/10.3390/ijms19113346
Xu Y, Zeng X, Wu J, Zhang F, Li C, Jiang J, Wang Y, Sun W. iTRAQ-Based Quantitative Proteome Revealed Metabolic Changes in Winter Turnip Rape (Brassica rapa L.) under Cold Stress. International Journal of Molecular Sciences. 2018; 19(11):3346. https://doi.org/10.3390/ijms19113346
Chicago/Turabian StyleXu, Yaozhao, Xiucun Zeng, Jian Wu, Fenqin Zhang, Caixia Li, Jinjin Jiang, Youping Wang, and Wancang Sun. 2018. "iTRAQ-Based Quantitative Proteome Revealed Metabolic Changes in Winter Turnip Rape (Brassica rapa L.) under Cold Stress" International Journal of Molecular Sciences 19, no. 11: 3346. https://doi.org/10.3390/ijms19113346
APA StyleXu, Y., Zeng, X., Wu, J., Zhang, F., Li, C., Jiang, J., Wang, Y., & Sun, W. (2018). iTRAQ-Based Quantitative Proteome Revealed Metabolic Changes in Winter Turnip Rape (Brassica rapa L.) under Cold Stress. International Journal of Molecular Sciences, 19(11), 3346. https://doi.org/10.3390/ijms19113346