Role of the Complement System in the Response to Orthopedic Biomaterials



{kind=link}
{kind=link}
Abstract
:1. Introduction
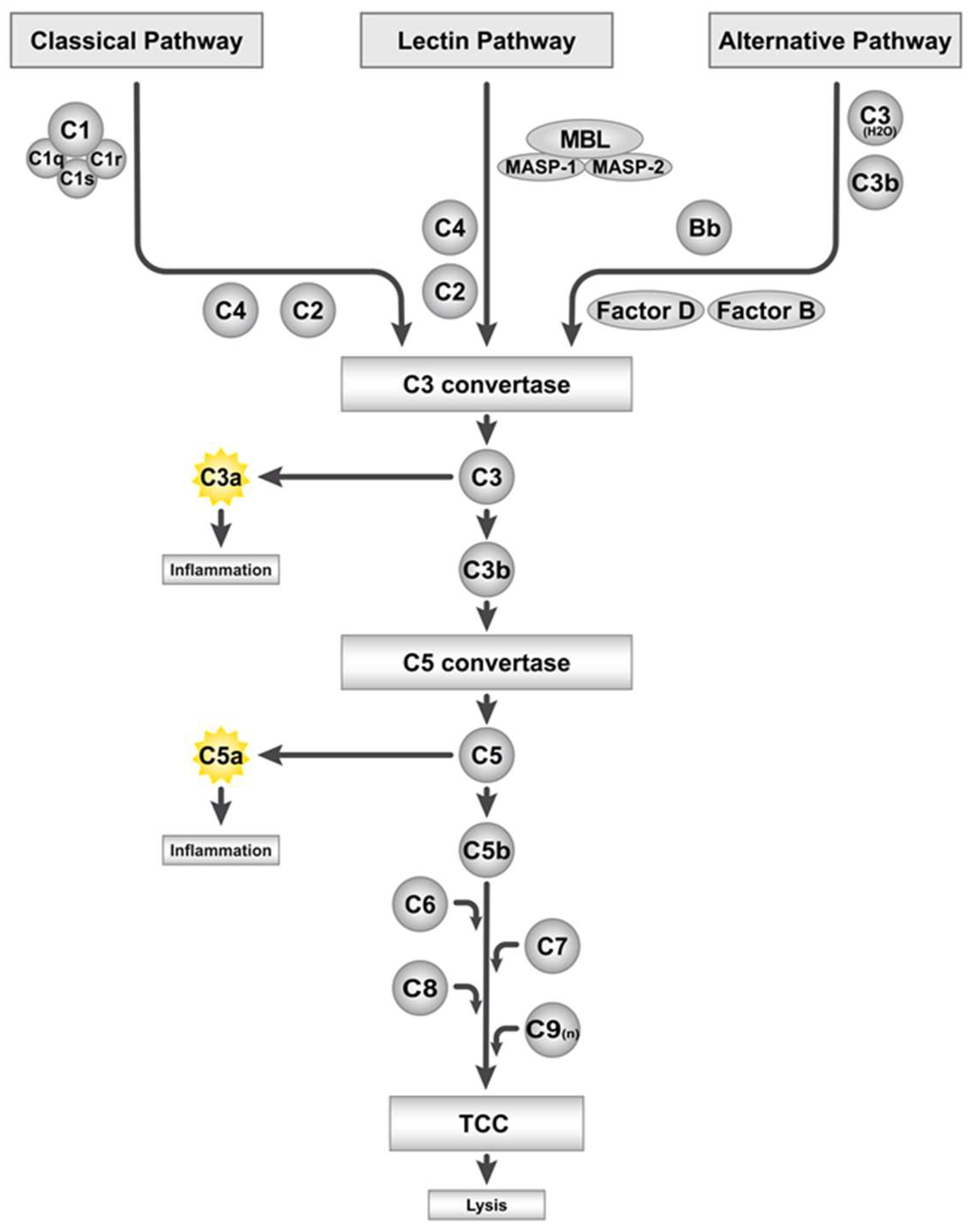
2. The Complement System and Its Activation by Artificial Surfaces
3. Role of the Complement System in Bone
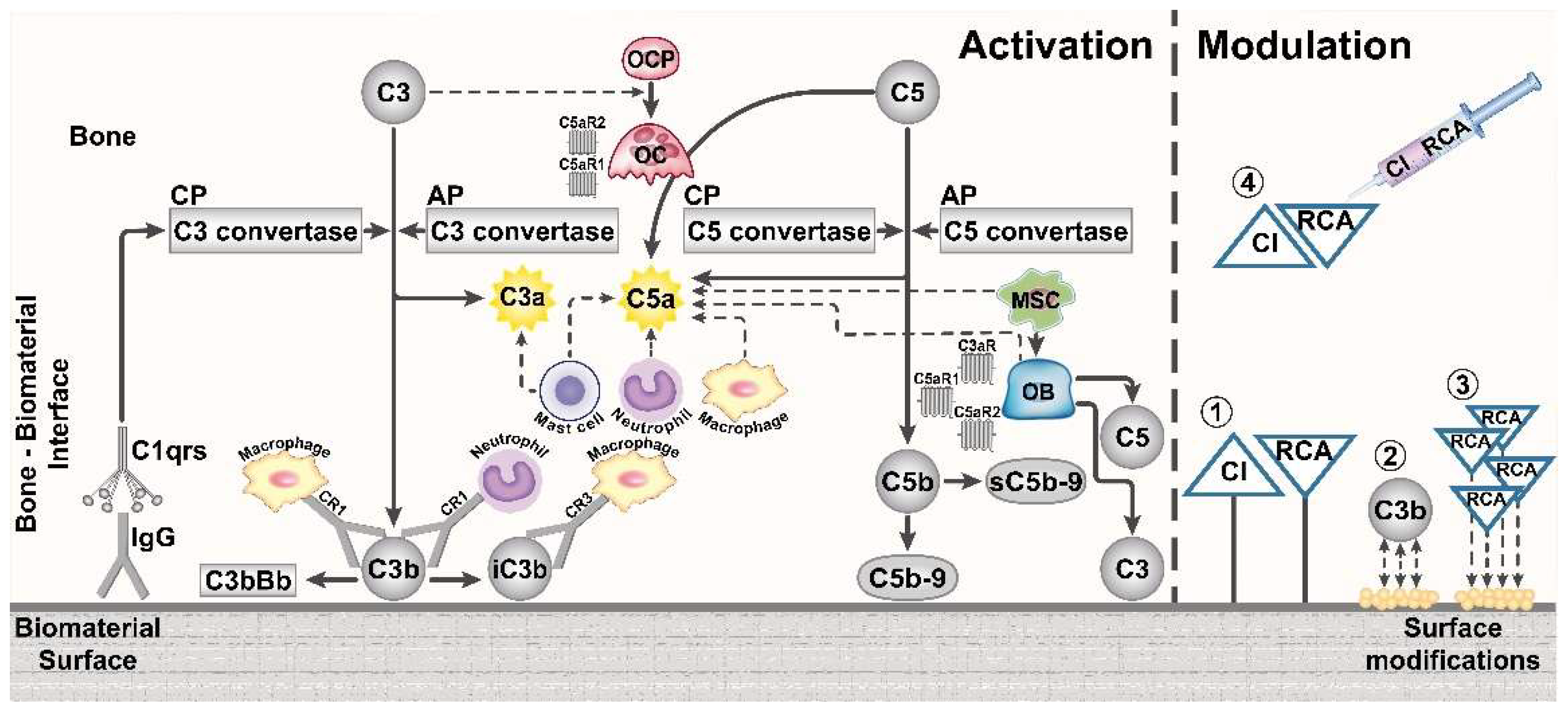
4. Potential Role of the Complement System in the Immune Response to Bone Biomaterials
4.1. Metals
4.2. Ceramics
4.3. Polymers
5. Modification of Biomaterial Surfaces to Influence Complement Activation
6. Conclusions and Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brydone, A.S.; Meek, D.; Maclaine, S. Bone grafting, orthopedic biomaterials, and the clinical need for bone engineering. Proc. Inst. Mech. Eng. H 2010, 224, 1329–1343. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, W.; Sendi, P. Pathogenesis of implant-associated infection: the role of the host. Semin. Immunopathol. 2011, 33, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Mödinger, Y.; Loffler, B.; Huber-Lang, M.; Ignatius, A. Complement involvement in bone homeostasis and bone disorders. Semin. Immunol. 2018, 37, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Schoengraf, P.; Lambris, J.D.; Recknagel, S.; Kreja, L.; Liedert, A.; Brenner, R.E.; Huber-Lang, M.; Ignatius, A. Does complement play a role in bone development and regeneration? Immunobiology 2013, 218, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ekdahl, K.N.; Lambris, J.D.; Elwing, H.; Ricklin, D.; Nilsson, P.H.; Teramura, Y.; Nicholls, I.A.; Nilsson, B. Innate immunity activation on biomaterial surfaces: A mechanistic model and coping strategies. Adv. Drug Deliv. Rev. 2011, 63, 1042–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engberg, A.E.; Rosengren-Holmberg, J.P.; Chen, H.; Nilsson, B.; Lambris, J.D.; Nicholls, I.A.; Ekdahl, K.N. Blood protein-polymer adsorption: Implications for understanding complement-mediated hemoincompatibility. J. Biomed. Mater. Res. A 2011, 97, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, B.; Ekdahl, K.N.; Mollnes, T.E.; Lambris, J.D. The role of complement in biomaterial-induced inflammation. Mol. Immunol. 2007, 44, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Lambris, J.D.; Reid, K.B.; Volanakis, J.E. The evolution, structure, biology and pathophysiology of complement. Immunol. Today 1999, 20, 207–211. [Google Scholar] [CrossRef]
- Noris, M.; Remuzzi, G. Overview of complement activation and regulation. Semin. Nephrol. 2013, 33, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Klos, A.; Tenner, A.J.; Johswich, K.O.; Ager, R.R.; Reis, E.S.; Kohl, J. The role of the anaphylatoxins in health and disease. Mol. Immunol. 2009, 46, 2753–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.F.; Ward, P.A. Role of C5a in inflammatory responses. Annu. Rev. Immunol. 2005, 23, 821–852. [Google Scholar] [CrossRef] [PubMed]
- Tegla, C.A.; Cudrici, C.; Patel, S.; Trippe, R., 3rd; Rus, V.; Niculescu, F.; Rus, H. Membrane attack by complement: the assembly and biology of terminal complement complexes. Immunol. Res. 2011, 51, 45–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serna, M.; Giles, J.L.; Morgan, B.P.; Bubeck, D. Structural basis of complement membrane attack complex formation. Nat. Commun. 2016, 7, 10587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubbers, R.; van Essen, M.F.; van Kooten, C.; Trouw, L.A. Production of complement components by cells of the immune system. Clin. Exp. Immunol. 2017, 188, 183–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Hong, M.H.; Jin, C.H.; Ishimi, Y.; Udagawa, N.; Shinki, T.; Abe, E.; Suda, T. The specific production of the third component of complement by osteoblastic cells treated with 1 alpha,25-dihydroxyvitamin D3. FEBS Lett. 1991, 285, 21–24. [Google Scholar] [CrossRef]
- Ignatius, A.; Schoengraf, P.; Kreja, L.; Liedert, A.; Recknagel, S.; Kandert, S.; Brenner, R.E.; Schneider, M.; Lambris, J.D.; Huber-Lang, M. Complement C3a and C5a modulate osteoclast formation and inflammatory response of osteoblasts in synergism with IL-1beta. J. Cell. Biochem. 2011, 112, 2594–2605. [Google Scholar] [CrossRef] [PubMed]
- Bajic, G.; Degn, S.E.; Thiel, S.; Andersen, G.R. Complement activation, regulation, and molecular basis for complement-related diseases. EMBO J. 2015, 34, 2735–2757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastellos, D.C.; Ricklin, D.; Hajishengallis, E.; Hajishengallis, G.; Lambris, J.D. Complement therapeutics in inflammatory diseases: promising drug candidates for C3-targeted intervention. Mol. Oral. Microbiol. 2016, 31, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Lambris, J.D. Complement-targeted therapeutics in periodontitis. Adv. Exp. Med. Biol. 2013, 735, 197–206. [Google Scholar] [PubMed]
- Ricklin, D.; Lambris, J.D. Complement therapeutics. Semin. Immunol. 2016, 28, 205–207. [Google Scholar] [CrossRef] [PubMed]
- Gorbet, M.B.; Sefton, M.V. Biomaterial-associated thrombosis: roles of coagulation factors, complement, platelets and leukocytes. Biomaterials 2004, 25, 5681–5703. [Google Scholar] [CrossRef] [PubMed]
- Sperling, C.; Schweiss, R.B.; Streller, U.; Werner, C. In vitro hemocompatibility of self-assembled monolayers displaying various functional groups. Biomaterials 2005, 26, 6547–6557. [Google Scholar] [CrossRef] [PubMed]
- Sperling, C.; Maitz, M.F.; Talkenberger, S.; Gouzy, M.F.; Groth, T.; Werner, C. In vitro blood reactivity to hydroxylated and non-hydroxylated polymer surfaces. Biomaterials 2007, 28, 3617–3625. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Liu, L.; Elwing, H.B. Complement activation and inflammation triggered by model biomaterial surfaces. J. Biomed. Mater. Res. 1998, 41, 333–340. [Google Scholar] [CrossRef]
- Mollnes, T.E. Complement and biocompatibility. Vox Sang 1998, 74 (Suppl 2), 303–307. [Google Scholar] [CrossRef]
- Franz, S.; Rammelt, S.; Scharnweber, D.; Simon, J.C. Immune responses to implants—a review of the implications for the design of immunomodulatory biomaterials. Biomaterials 2011, 32, 6692–6709. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.J.; Clegg, R.E.; Leavesley, D.I.; Pearcy, M.J. Mediation of biomaterial-cell interactions by adsorbed proteins: a review. Tissue Eng. 2005, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.; Ekdahl, K.N.; Lambris, J.D.; Nilsson, B. Binding of C3 fragments on top of adsorbed plasma proteins during complement activation on a model biomaterial surface. Biomaterials 2005, 26, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Andersson, J.; Ekdahl, K.N.; Larsson, R.; Nilsson, U.R.; Nilsson, B. C3 adsorbed to a polymer surface can form an initiating alternative pathway convertase. J. Immunol. 2002, 168, 5786–5791. [Google Scholar] [CrossRef] [PubMed]
- Tengvall, P.; Askendal, A.; Lundstrom, I.I. Ellipsometric in vitro studies on the activation of complement by human immunoglobulins M and G after adsorption to methylated silicon. Colloids Surf. B Biointerfaces 2001, 20, 51–62. [Google Scholar] [CrossRef]
- Fearon, D.T. Identification of the membrane glycoprotein that is the C3b receptor of the human erythrocyte, polymorphonuclear leukocyte, B lymphocyte, and monocyte. J. Exp. Med. 1980, 152, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnaout, M.A. Structure and function of the leukocyte adhesion molecules CD11/CD18. Blood 1990, 75, 1037–1050. [Google Scholar] [PubMed]
- Love, R.J.; Jones, K.S. The recognition of biomaterials: pattern recognition of medical polymers and their adsorbed biomolecules. J. Biomed. Mater. Res. A 2013, 101, 2740–2752. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Engberg, A.E.; Jonsson, N.; Sandholm, K.; Nicholls, I.A.; Mollnes, T.E.; Fromell, K.; Nilsson, B.; Ekdahl, K.N. Reciprocal relationship between contact and complement system activation on artificial polymers exposed to whole human blood. Biomaterials 2016, 77, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Marosok, R.; Washburn, R.; Indorf, A.; Solomon, D.; Sherertz, R. Contribution of vascular catheter material to the pathogenesis of infection: depletion of complement by silicone elastomer in vitro. J. Biomed. Mater. Res. 1996, 30, 245–250. [Google Scholar] [CrossRef]
- Kristian, S.A.; Birkenstock, T.A.; Sauder, U.; Mack, D.; Gotz, F.; Landmann, R. Biofilm formation induces C3a release and protects Staphylococcus epidermidis from IgG and complement deposition and from neutrophil-dependent killing. J. Infect. Dis. 2008, 197, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Orapiriyakul, W.; Young, P.S.; Damiati, L.; Tsimbouri, P.M. Antibacterial surface modification of titanium implants in orthopedics. J. Tissue. Eng. 2018, 9, 2041731418789838. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Steinle, H.; Golombek, S.; Hann, L.; Schlensak, C.; Wendel, H.P.; Avci-Adali, M. Blood-Contacting Biomaterials: In vitro evaluation of the hemocompatibility. Front. Bioeng. Biotechnol. 2018, 6, 99. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H. Osteoimmunology: Shared mechanisms and crosstalk between the immune and bone systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Greenblatt, M.B.; Shim, J.H. Osteoimmunology: a brief introduction. Immune. Netw. 2013, 13, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Abe, E.; Jin, C.H.; Hong, M.H.; Katagiri, T.; Kinoshita, T.; Amizuka, N.; Ozawa, H.; Suda, T. The biological roles of the third component of complement in osteoclast formation. Endocrinology 1993, 133, 397–404. [Google Scholar] [CrossRef]
- Jin, C.H.; Shinki, T.; Hong, M.H.; Sato, T.; Yamaguchi, A.; Ikeda, T.; Yoshiki, S.; Abe, E.; Suda, T. 1 alpha,25-dihydroxyvitamin D3 regulates in vivo production of the third component of complement (C3) in bone. Endocrinology 1992, 131, 2468–2475. [Google Scholar] [CrossRef] [PubMed]
- Ignatius, A.; Ehrnthaller, C.; Brenner, R.E.; Kreja, L.; Schoengraf, P.; Lisson, P.; Blakytny, R.; Recknagel, S.; Claes, L.; Gebhard, F.; et al. The anaphylatoxin receptor C5aR is present during fracture healing in rats and mediates osteoblast migration in vitro. J. Trauma. 2011, 71, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Schraufstatter, I.U.; Discipio, R.G.; Zhao, M.; Khaldoyanidi, S.K. C3a and C5a are chemotactic factors for human mesenchymal stem cells, which cause prolonged ERK1/2 phosphorylation. J. Immunol. 2009, 182, 3827–3836. [Google Scholar] [CrossRef] [PubMed]
- Bergdolt, S.; Kovtun, A.; Hägele, Y.; Liedert, A.; Schinke, T.; Amling, M.; Huber-Lang, M.; Ignatius, A. Osteoblast-specific overexpression of complement receptor C5aR1 impairs fracture healing. PLoS ONE 2017, 12, e0179512. [Google Scholar] [CrossRef] [PubMed]
- Pobanz, J.M.; Reinhardt, R.A.; Koka, S.; Sanderson, S.D. C5a modulation of interleukin-1 beta-induced interleukin-6 production by human osteoblast-like cells. J. Periodontal. Res. 2000, 35, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Mödinger, Y.; Rapp, A.E.; Pazmandi, J.; Vikman, A.; Holzmann, K.; Haffner-Luntzer, M.; Huber-Lang, M.; Ignatius, A. C5aR1 interacts with TLR2 in osteoblasts and stimulates the osteoclast-inducing chemokine CXCL10. J. Cell. Mol. Med. 2018. In press. [Google Scholar]
- Tu, Z.; Bu, H.; Dennis, J.E.; Lin, F. Efficient osteoclast differentiation requires local complement activation. Blood 2010, 116, 4456–4463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovtun, A.; Bergdolt, S.; Hägele, Y.; Matthes, R.; Lambris, J.D.; Huber-Lang, M.; Ignatius, A. Complement receptors C5aR1 and C5aR2 act differentially during the early immune response after bone fracture but are similarly involved in bone repair. Sci. Rep. 2017, 7, 14061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber-Lang, M.; Kovtun, A.; Ignatius, A. The role of complement in trauma and fracture healing. Semin. Immunol. 2013, 25, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Mödinger, Y.; Rapp, A.E.; Vikman, A.; Ren, Z.; Fischer, V.; Bergdolt, S.; Haffner-Luntzer, M.; Song, W.C.; Lambris, J.D.; Huber-Lang, M.; et al. Reduced terminal complement complex formation in mice manifests in low bone mass and impaired fracture healing. Am. J. Pathol. 2018. In press. [Google Scholar]
- Ehrnthaller, C.; Huber-Lang, M.; Nilsson, P.; Bindl, R.; Redeker, S.; Recknagel, S.; Rapp, A.; Mollnes, T.; Amling, M.; Gebhard, F.; et al. Complement C3 and C5 deficiency affects fracture healing. PLoS ONE 2013, 8, e81341. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.E. Mechanisms of endosseous integration. Int. J. Prosthodont. 1998, 11, 391–401. [Google Scholar] [PubMed]
- Claes, L.; Recknagel, S.; Ignatius, A. Fracture healing under healthy and inflammatory conditions. Nat. Rev. Rheumatol. 2012, 8, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Trindade, R.; Albrektsson, T.; Tengvall, P.; Wennerberg, A. Foreign Body Reaction to Biomaterials: On Mechanisms for Buildup and Breakdown of Osseointegration. Clin. Implant. Dent. Relat. Res. 2016, 18, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Albrektsson, T.; Dahlin, C.; Jemt, T.; Sennerby, L.; Turri, A.; Wennerberg, A. Is marginal bone loss around oral implants the result of a provoked foreign body reaction? Clin. Implant. Dent. Relat. Res. 2014, 16, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Donath, K.; Laass, M.; Gunzl, H.J. The histopathology of different foreign-body reactions in oral soft tissue and bone tissue. Virchows Arch. A Pathol. Anat. Histopathol. 1992, 420, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, Z.; Brooks, P.J.; Barzilay, O.; Fine, N.; Glogauer, M. Macrophages, Foreign Body Giant Cells and Their Response to Implantable Biomaterials. Materials 2015, 8, 5671–5701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milde, R.; Ritter, J.; Tennent, G.A.; Loesch, A.; Martinez, F.O.; Gordon, S.; Pepys, M.B.; Verschoor, A.; Helming, L. Multinucleated Giant Cells Are Specialized for Complement-Mediated Phagocytosis and Large Target Destruction. Cell Rep. 2015, 13, 1937–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Z.; Triffitt, J.T. A review on macrophage responses to biomaterials. Biomed. Mater. 2006, 1, R1–R9. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamaki, Y.; Sasaki, K.; Sasaki, A.; Takakubo, Y.; Hasegawa, H.; Ogino, T.; Konttinen, Y.T.; Salo, J.; Takagi, M. Enhanced osteolytic potential of monocytes/macrophages derived from bone marrow after particle stimulation. J. Biomed. Mater. Res. B Appl. Biomater. 2008, 84, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Athanasou, N.A. The pathobiology and pathology of aseptic implant failure. Bone Joint Res. 2016, 5, 162–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trindade, R.; Albrektsson, T.; Wennerberg, A. Current concepts for the biological basis of dental implants: foreign body equilibrium and osseointegration dynamics. Oral Maxillofac. Surg. Clin. North Am. 2015, 27, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.; Michiardi, A.; Castano, O.; Planell, J.A. Biomaterials in orthopedics. J. R Soc. Interface 2008, 5, 1137–1158. [Google Scholar] [CrossRef] [PubMed]
- Markatos, K.; Tsoucalas, G.; Sgantzos, M. Hallmarks in the history of orthopedic implants for trauma and joint replacement. Acta. Med. Hist. Adriat. 2016, 14, 161–176. [Google Scholar] [PubMed]
- Tamaddon, M.; Samizadeh, S.; Wang, L.; Blunn, G.; Liu, C. Intrinsic Osteoinductivity of Porous Titanium Scaffold for Bone Tissue Engineering. Int. J. Biomater. 2017, 2017, 5093063. [Google Scholar] [CrossRef] [PubMed]
- Dabrowski, B.; Swieszkowski, W.; Godlinski, D.; Kurzydlowski, K.J. Highly porous titanium scaffolds for orthopedic applications. J. Biomed. Mater. Res. B Appl. Biomater. 2010, 95, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Bijukumar, D.R.; Segu, A.; Souza, J.C.M.; Li, X.; Barba, M.; Mercuri, L.G.; J, J.J.; Mathew, M.T. Systemic and local toxicity of metal debris released from hip prostheses: A review of experimental approaches. Nanomedicine 2018, 14, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, P.F.; Lichstein, P.M.; Shen, C.; Tokarski, A.T.; Parvizi, J. Why are total knee arthroplasties failing today--has anything changed after 10 years? J. Arthroplasty 2014, 29, 1774–1778. [Google Scholar] [CrossRef] [PubMed]
- Wawrzynski, J.; Gil, J.A.; Goodman, A.D.; Waryasz, G.R. Hypersensitivity to Orthopedic Implants: A Review of the Literature. Rheumatol. Ther. 2017, 4, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, L.; Flanagan, B.F.; Hunt, J.A. Flow cytometric measurement of phagocytosis reveals a role for C3b in metal particle uptake by phagocytes. J. Biomed. Mater. Res. A 2005, 73, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Perala, D.; Chapman, R.; Gelfand, J. Complement activation by dental implants. Int. J. Oral Maxillofac. Implants 1991, 6, 136–141. [Google Scholar] [PubMed]
- Arvidsson, S.; Askendal, A.; Tengvall, P. Blood plasma contact activation on silicon, titanium and aluminium. Biomaterials 2007, 28, 1346–1354. [Google Scholar] [CrossRef] [PubMed]
- Nygren, H.; Tengvall, P.; Lundstrom, I. The initial reactions of TiO2 with blood. J. Biomed. Mater. Res. 1997, 34, 487–492. [Google Scholar] [CrossRef]
- Walivaara, B.; Askendal, A.; Lundstrom, I.; Tengvall, P. Blood protein interactions with titanium surfaces. J. Biomater. Sci. Polym. Ed. 1996, 8, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Romero-Gavilan, F.; Gomes, N.C.; Rodenas, J.; Sanchez, A.; Azkargorta, M.; Iloro, I.; Elortza, F.; Garcia Arnaez, I.; Gurruchaga, M.; Goni, I.; et al. Proteome analysis of human serum proteins adsorbed onto different titanium surfaces used in dental implants. Biofouling 2017, 33, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Linderback, P.; Harmankaya, N.; Askendal, A.; Areva, S.; Lausmaa, J.; Tengvall, P. The effect of heat- or ultra violet ozone-treatment of titanium on complement deposition from human blood plasma. Biomaterials 2010, 31, 4795–4801. [Google Scholar] [CrossRef] [PubMed]
- Sawase, T.; Jimbo, R.; Baba, K.; Shibata, Y.; Ikeda, T.; Atsuta, M. Photo-induced hydrophilicity enhances initial cell behavior and early bone apposition. Clin. Oral. Implants Res. 2008, 19, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Harmankaya, N.; Igawa, K.; Stenlund, P.; Palmquist, A.; Tengvall, P. Healing of complement activating Ti implants compared with non-activating Ti in rat tibia. Acta Biomater. 2012, 8, 3532–3540. [Google Scholar] [CrossRef] [PubMed]
- Trindade, R.; Albrektsson, T.; Galli, S.; Prgomet, Z.; Tengvall, P.; Wennerberg, A. Osseointegration and foreign body reaction: Titanium implants activate the immune system and suppress bone resorption during the first 4 weeks after implantation. Clin. Implant Dent. Relat. Res. 2018, 20, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Romero-Gavilan, F.; Araujo-Gomes, N.; Sanchez-Perez, A.M.; Garcia-Arnaez, I.; Elortza, F.; Azkargorta, M.; de Llano, J.J.M.; Carda, C.; Gurruchaga, M.; Suay, J.; et al. Bioactive potential of silica coatings and its effect on the adhesion of proteins to titanium implants. Colloids Surf. B Biointerfaces 2018, 162, 316–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo-Gomes, N.; Romero-Gavilan, F.; Sanchez-Perez, A.M.; Gurruchaga, M.; Azkargorta, M.; Elortza, F.; Martinez-Ibanez, M.; Iloro, I.; Suay, J.; Goni, I. Characterization of serum proteins attached to distinct sol-gel hybrid surfaces. J. Biomed. Mater. Res. B Appl. Biomater. 2018, 106, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Araujo-Gomes, N.; Romero-Gavilan, F.; Lara-Saez, I.; Elortza, F.; Azkargorta, M.; Iloro, I.; Martinez-Ibanez, M.; Martin de Llano, J.J.; Gurruchaga, M.; Goni, I.; et al. Silica-gelatin hybrid sol-gel coatings: A proteomic study with biocompatibility implications. J. Tissue. Eng. Regen. Med. 2018, 12, 1769–1779. [Google Scholar] [CrossRef] [PubMed]
- Hulsart-Billstrom, G.; Dawson, J.I.; Hofmann, S.; Muller, R.; Stoddart, M.J.; Alini, M.; Redl, H.; El Haj, A.; Brown, R.; Salih, V.; et al. A surprisingly poor correlation between in vitro and in vivo testing of biomaterials for bone regeneration: results of a multicentre analysis. Eur. Cells. Mater. 2016, 31, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Thamaraiselvi, T.V.; Rajeswari, S. Biological Evaluation of Bioceramic Materials. Trends Biomater. Aertif. Organs 2004, 18, 9–17. [Google Scholar]
- Ghalme, S.G.; Mankar, A.; Bhalerao, Y. Biomaterials in Hip Joint Replacement. Int. J. Mater. Sci. Eng. 2016, 4, 113–125. [Google Scholar]
- Remes, A.; Williams, D.F. Relationship between chemotaxis and complement activation by ceramic biomaterials. Biomaterials 1991, 12, 661–667. [Google Scholar] [CrossRef]
- Klein, C.P.; de Groot, K.; van Kamp, G. Activation of complement C3 by different calcium phosphate powders. Biomaterials 1983, 4, 181–184. [Google Scholar] [CrossRef]
- Tengvall, P.; Askendal, A. Ellipsometric in vitro studies on blood plasma and serum adsorption to zirconium. J. Biomed. Mater. Res. 2001, 57, 285–290. [Google Scholar] [CrossRef]
- Rosengren, A.; Pavlovic, E.; Oscarsson, S.; Krajewski, A.; Ravaglioli, A.; Piancastelli, A. Plasma protein adsorption pattern on characterized ceramic biomaterials. Biomaterials 2002, 23, 1237–1247. [Google Scholar] [CrossRef]
- Ferraz, N.; Nilsson, B.; Hong, J.; Karlsson Ott, M. Nanoporesize affects complement activation. J. Biomed. Mater. Res. A 2008, 87, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheikh, Z.; Najeeb, S.; Khurshid, Z.; Verma, V.; Rashid, H.; Glogauer, M. Biodegradable Materials for Bone Repair and Tissue Engineering Applications. Materials 2015, 8, 5744–5794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeHeer, D.H.; Engels, J.A.; DeVries, A.S.; Knapp, R.H.; Beebe, J.D. In situ complement activation by polyethylene wear debris. J. Biomed. Mater. Res. 2001, 54, 12–19. [Google Scholar] [CrossRef]
- Wooley, P.H.; Fitzgerald, R.H., Jr.; Song, Z.; Davis, P.; Whalen, J.D.; Trumble, S.; Nasser, S. Proteins bound to polyethylene components in patients who have aseptic loosening after total joint arthroplasty. A preliminary report. J. Bone Joint Surg. Am. 1999, 81, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Pascual, M.; Plastre, O.; Montdargent, B.; Labarre, D.; Schifferli, J.A. Specific interactions of polystyrene biomaterials with factor D of human complement. Biomaterials 1993, 14, 665–670. [Google Scholar] [CrossRef]
- Rakshit, D.S.; Lim, J.T.; Ly, K.; Ivashkiv, L.B.; Nestor, B.J.; Sculco, T.P.; Purdue, P.E. Involvement of complement receptor 3 (CR3) and scavenger receptor in macrophage responses to wear debris. J. Orthop. Res. 2006, 24, 2036–2044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noordin, S.; Shortkroff, S.; Sledge, C.B.; Spector, M. Investigation of the activation of a human serum complement protein, C3, by orthopedic prosthetic particulates. Biomaterials 2004, 25, 5347–5352. [Google Scholar] [CrossRef] [PubMed]
- Rokstad, A.M.; Brekke, O.L.; Steinkjer, B.; Ryan, L.; Kollarikova, G.; Strand, B.L.; Skjak-Braek, G.; Lambris, J.D.; Lacik, I.; Mollnes, T.E.; et al. The induction of cytokines by polycation containing microspheres by a complement dependent mechanism. Biomaterials 2013, 34, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Gravastrand, C.; Hamad, S.; Fure, H.; Steinkjer, B.; Ryan, L.; Oberholzer, J.; Lambris, J.D.; Lacik, I.; Mollnes, T.E.; Espevik, T.; et al. Alginate microbeads are coagulation compatible, while alginate microcapsules activate coagulation secondary to complement or directly through FXII. Acta Biomater. 2017, 58, 158–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakasam, M.; Locs, J.; Salma-Ancane, K.; Loca, D.; Largeteau, A.; Berzina-Cimdina, L. Biodegradable Materials and Metallic Implants-A Review. J. Funct. Biomater. 2017, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Tegnander, A.; Engebretsen, L.; Bergh, K.; Eide, E.; Holen, K.J.; Iversen, O.J. Activation of the complement system and adverse effects of biodegradable pins of polylactic acid (Biofix) in osteochondritis dissecans. Acta Orthop. Scand. 1994, 65, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Ceonzo, K.; Gaynor, A.; Shaffer, L.; Kojima, K.; Vacanti, C.A.; Stahl, G.L. Polyglycolic acid-induced inflammation: role of hydrolysis and resulting complement activation. Tissue Eng. 2006, 12, 301–308. [Google Scholar] [CrossRef] [PubMed]
- James, K.; Levene, H.; Parsons, J.R.; Kohn, J. Small changes in polymer chemistry have a large effect on the bone-implant interface: evaluation of a series of degradable tyrosine-derived polycarbonates in bone defects. Biomaterials 1999, 20, 2203–2212. [Google Scholar] [CrossRef]
- Magno, M.H.R.; Kim, J.; Srinivasan, A.; McBride, S.; Bolikal, D.; Darr, A.; Hollinger, J.O.; Kohn, J. Synthesis, degradation and biocompatibility of tyrosine-derived polycarbonate scaffolds. J. Mater. Chem. 2010, 20, 8885–8893. [Google Scholar] [CrossRef]
- Sokolov, A.; Hellerud, B.C.; Pharo, A.; Johannessen, E.A.; Mollnes, T.E. Complement activation by candidate biomaterials of an implantable microfabricated medical device. J. Biomed. Mater. Res. B Appl. Biomater. 2011, 98, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Toskas, G.; Cherif, C.; Hund, R.D.; Laourine, E.; Mahltig, B.; Fahmi, A.; Heinemann, C.; Hanke, T. Chitosan (PEO)/silica hybrid nanofibers as a potential biomaterial for bone regeneration. Carbohydr. Polym. 2013, 94, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Dong, Y.; Xiao, J.; Gu, R.; Ding, M.; Huang, T.; Li, J.; Zhao, N.; Liao, H. In vivo immuno-reactivity analysis of the porous three-dimensional chitosan/SiO2 and chitosan/SiO2 /hydroxyapatite hybrids. J. Biomed. Mater. Res. A 2018, 106, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, S.; Fujihara, Y.; Sakamoto, T.; Asawa, Y.; Komura, M.; Nagata, S.; Takato, T.; Hoshi, K. Tissue responses against tissue-engineered cartilage consisting of chondrocytes encapsulated within non-absorbable hydrogel. J. Tissue. Eng. Regen. Med. 2013, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Arima, Y.; Kawagoe, M.; Toda, M.; Iwata, H. Complement activation by polymers carrying hydroxyl groups. ACS Appl. Mater. Interfaces 2009, 1, 2400–2407. [Google Scholar] [CrossRef] [PubMed]
- Rupp, F.; Liang, L.; Geis-Gerstorfer, J.; Scheideler, L.; Huttig, F. Surface characteristics of dental implants: A review. Dent. Mater. 2018, 34, 40–57. [Google Scholar] [CrossRef] [PubMed]
- Rungsiyakull, C.; Li, Q.; Sun, G.; Li, W.; Swain, M.V. Surface morphology optimization for osseointegration of coated implants. Biomaterials 2010, 31, 7196–7204. [Google Scholar] [CrossRef] [PubMed]
- Milleret, V.; Tugulu, S.; Schlottig, F.; Hall, H. Alkali treatment of microrough titanium surfaces affects macrophage/monocyte adhesion, platelet activation and architecture of blood clot formation. Eur. Cell Mater. 2011, 21, 430–444. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Klein, T.; Murray, R.Z.; Crwford, R.; Chang, J.; Chengtie, W.; Xiao, Y. Osteoimmunomodulation for the development of advanced bone biomaterials. Mater. Today 2016, 19, 304–321. [Google Scholar] [CrossRef] [Green Version]
- Thevenot, P.; Hu, W.; Tang, L. Surface chemistry influences implant biocompatibility. Curr. Top. Med. Chem. 2008, 8, 270–280. [Google Scholar] [PubMed]
- Hulander, M.; Lundgren, A.; Berglin, M.; Ohrlander, M.; Lausmaa, J.; Elwing, H. Immune complement activation is attenuated by surface nanotopography. Int. J. Nanomedicine 2011, 6, 2653–2666. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, B.; Korsgren, O.; Lambris, J.D.; Ekdahl, K.N. Can cells and biomaterials in therapeutic medicine be shielded from innate immune recognition? Trends Immunol. 2010, 31, 32–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeAngelis, R.A.; Reis, E.S.; Ricklin, D.; Lambris, J.D. Targeted complement inhibition as a promising strategy for preventing inflammatory complications in hemodialysis. Immunobiology 2012, 217, 1097–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pekna, M.; Larsson, R.; Formgren, B.; Nilsson, U.R.; Nilsson, B. Complement activation by polymethyl methacrylate minimized by end-point heparin attachment. Biomaterials 1993, 14, 189–192. [Google Scholar] [CrossRef]
- Videm, V.; Svennevig, J.L.; Fosse, E.; Semb, G.; Osterud, A.; Mollnes, T.E. Reduced complement activation with heparin-coated oxygenator and tubings in coronary bypass operations. J. Thorac. Cardiovasc. Surg. 1992, 103, 806–813. [Google Scholar] [PubMed]
- Videm, V.; Mollnes, T.E.; Bergh, K.; Fosse, E.; Mohr, B.; Hagve, T.A.; Aasen, A.O.; Svennevig, J.L. Heparin-coated cardiopulmonary bypass equipment. II. Mechanisms for reduced complement activation in vivo. J. Thorac. Cardiovasc. Surg. 1999, 117, 803–809. [Google Scholar] [PubMed]
- Watkins, N.J.; Braidley, P.; Bray, C.J.; Savill, C.M.; White, D.J. Coating of human decay accelerating factor (hDAF) onto medical devices to improve biocompatibility. Immunopharmacology 1997, 38, 111–118. [Google Scholar] [CrossRef]
- Gillinov, A.M.; DeValeria, P.A.; Winkelstein, J.A.; Wilson, I.; Curtis, W.E.; Shaw, D.; Yeh, C.G.; Rudolph, A.R.; Baumgartner, W.A.; Herskowitz, A.; et al. Complement inhibition with soluble complement receptor type 1 in cardiopulmonary bypass. Ann. Thorac. Surg. 1993, 55, 619–624. [Google Scholar] [CrossRef]
- Lappegard, K.T.; Fung, M.; Bergseth, G.; Riesenfeld, J.; Lambris, J.D.; Videm, V.; Mollnes, T.E. Effect of complement inhibition and heparin coating on artificial surface-induced leukocyte and platelet activation. Ann. Thorac. Surg. 2004, 77, 932–941. [Google Scholar] [CrossRef]
- Nilsson, B.; Larsson, R.; Hong, J.; Elgue, G.; Ekdahl, K.N.; Sahu, A.; Lambris, J.D. Compstatin inhibits complement and cellular activation in whole blood in two models of extracorporeal circulation. Blood 1998, 92, 1661–1667. [Google Scholar] [CrossRef]
- Schmidt, S.; Haase, G.; Csomor, E.; Lutticken, R.; Peltroche-Llacsahuanga, H. Inhibitor of complement, Compstatin, prevents polymer-mediated Mac-1 up-regulation of human neutrophils independent of biomaterial type tested. J. Biomed. Mater. Res. A 2003, 66, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Kourtzelis, I.; Markiewski, M.M.; Doumas, M.; Rafail, S.; Kambas, K.; Mitroulis, I.; Panagoutsos, S.; Passadakis, P.; Vargemezis, V.; Magotti, P.; et al. Complement anaphylatoxin C5a contributes to hemodialysis-associated thrombosis. Blood 2010, 116, 631–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, E.S.; DeAngelis, R.A.; Chen, H.; Resuello, R.R.; Ricklin, D.; Lambris, J.D. Therapeutic C3 inhibitor Cp40 abrogates complement activation induced by modern hemodialysis filters. Immunobiology 2015, 220, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, J.; Larsson, R.; Richter, R.; Ekdahl, K.N.; Nilsson, B. Binding of a model regulator of complement activation (RCA) to a biomaterial surface: surface-bound factor H inhibits complement activation. Biomaterials 2001, 22, 2435–2443. [Google Scholar] [CrossRef]
- Andersson, J.; Bexborn, F.; Klinth, J.; Nilsson, B.; Ekdahl, K.N. Surface-attached PEO in the form of activated Pluronic with immobilized factor H reduces both coagulation and complement activation in a whole-blood model. J. Biomed. Mater. Res. A 2006, 76, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Q.; Qu, H.; Sfyroera, G.; Tzekou, A.; Kay, B.K.; Nilsson, B.; Nilsson Ekdahl, K.; Ricklin, D.; Lambris, J.D. Protection of nonself surfaces from complement attack by factor H-binding peptides: implications for therapeutic medicine. J. Immunol. 2011, 186, 4269–4277. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.C.; Prosser, B.E.; Caesar, J.J.; Kugelberg, E.; Li, S.; Zhang, Q.; Quoraishi, S.; Lovett, J.E.; Deane, J.E.; Sim, R.B.; et al. Neisseria meningitidis recruits factor H using protein mimicry of host carbohydrates. Nature 2009, 458, 890–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambris, J.D.; Ricklin, D.; Geisbrecht, B.V. Complement evasion by human pathogens. Nat. Rev. Microbiol. 2008, 6, 132–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serruto, D.; Rappuoli, R.; Scarselli, M.; Gros, P.; van Strijp, J.A. Molecular mechanisms of complement evasion: learning from staphylococci and meningococci. Nat. Rev. Microbiol. 2010, 8, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Engberg, A.E.; Sandholm, K.; Bexborn, F.; Persson, J.; Nilsson, B.; Lindahl, G.; Ekdahl, K.N. Inhibition of complement activation on a model biomaterial surface by streptococcal M protein-derived peptides. Biomaterials 2009, 30, 2653–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mödinger, Y.; Teixeira, G.Q.; Neidlinger-Wilke, C.; Ignatius, A. Role of the Complement System in the Response to Orthopedic Biomaterials. Int. J. Mol. Sci. 2018, 19, 3367. https://doi.org/10.3390/ijms19113367
Mödinger Y, Teixeira GQ, Neidlinger-Wilke C, Ignatius A. Role of the Complement System in the Response to Orthopedic Biomaterials. International Journal of Molecular Sciences. 2018; 19(11):3367. https://doi.org/10.3390/ijms19113367
Chicago/Turabian StyleMödinger, Yvonne, Graciosa Q. Teixeira, Cornelia Neidlinger-Wilke, and Anita Ignatius. 2018. "Role of the Complement System in the Response to Orthopedic Biomaterials" International Journal of Molecular Sciences 19, no. 11: 3367. https://doi.org/10.3390/ijms19113367
APA StyleMödinger, Y., Teixeira, G. Q., Neidlinger-Wilke, C., & Ignatius, A. (2018). Role of the Complement System in the Response to Orthopedic Biomaterials. International Journal of Molecular Sciences, 19(11), 3367. https://doi.org/10.3390/ijms19113367