Selenium-Binding Protein 1 in Human Health and Disease


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. SBP1 Discovery
3. The Role of Se in SBP1
4. SBP1 Levels Are Reduced in Cancer and Low Levels Are Predictive of Clinical Outcome
4.1. Is SBP1 a Tumor Suppressor?
4.2. Is SBP1 Downregulation an Early or Late Event in the Process of Tumorigenesis?
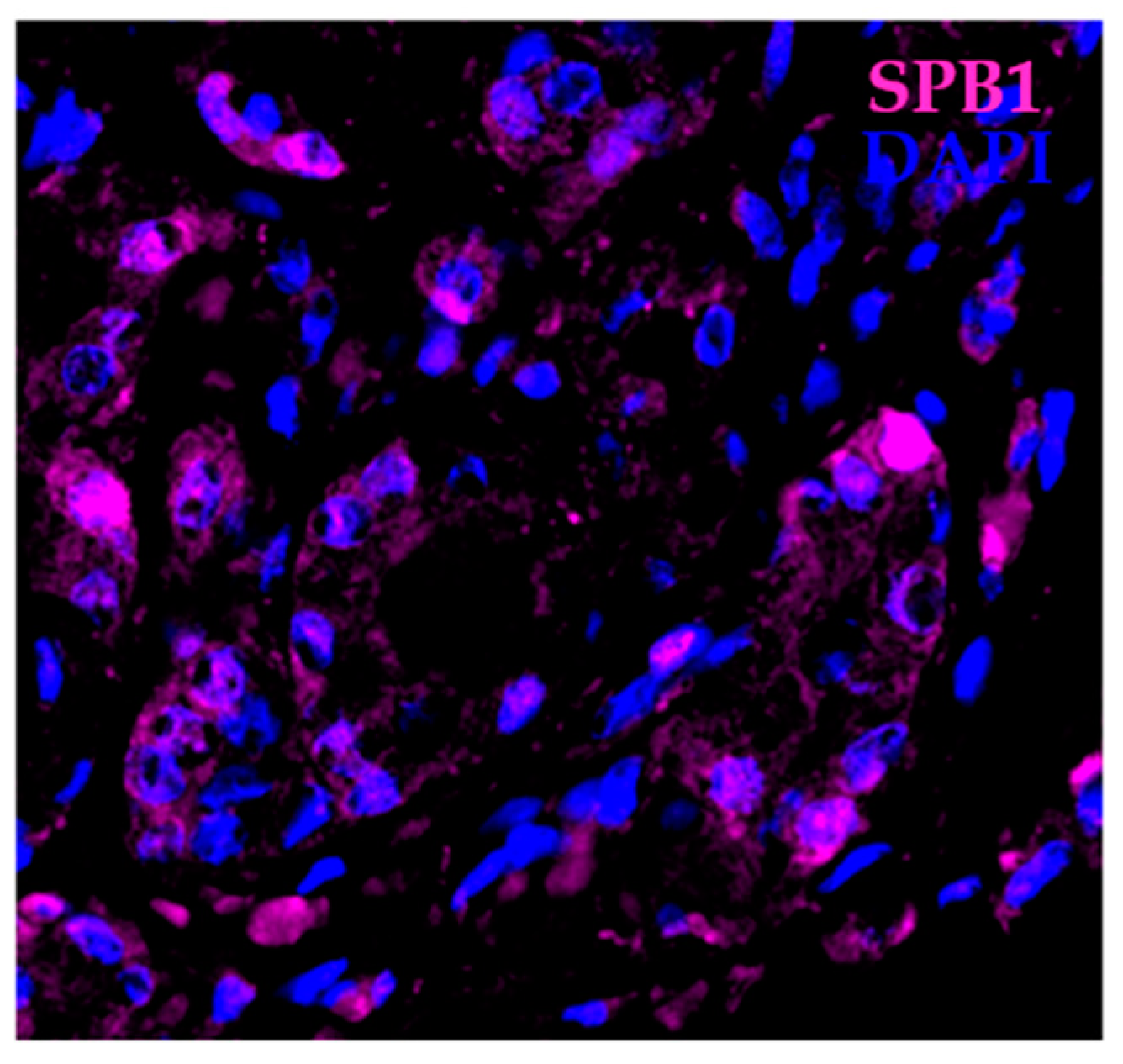
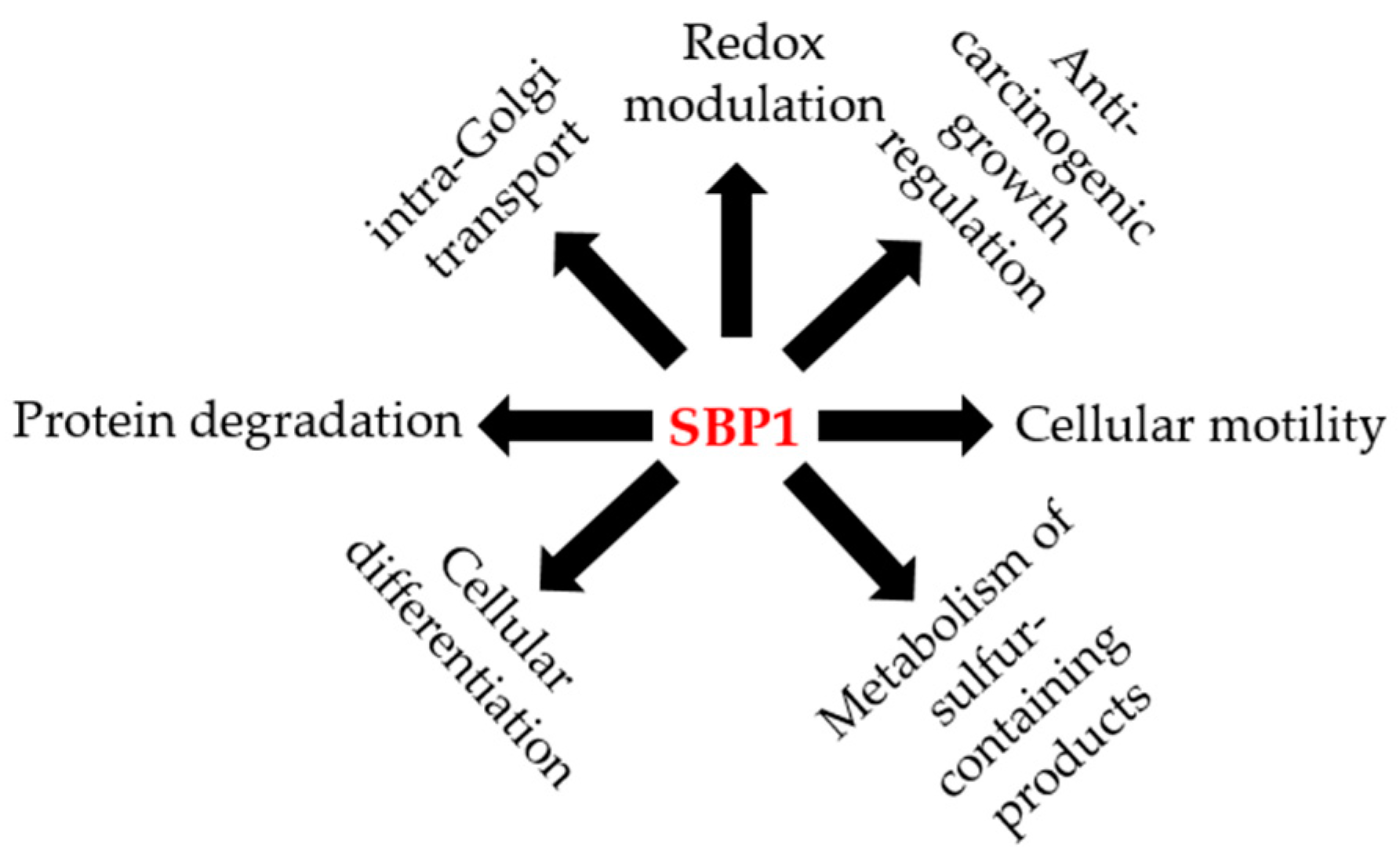
5. Physiological Roles for SBP1
Tissue-Specific Roles for SBP1
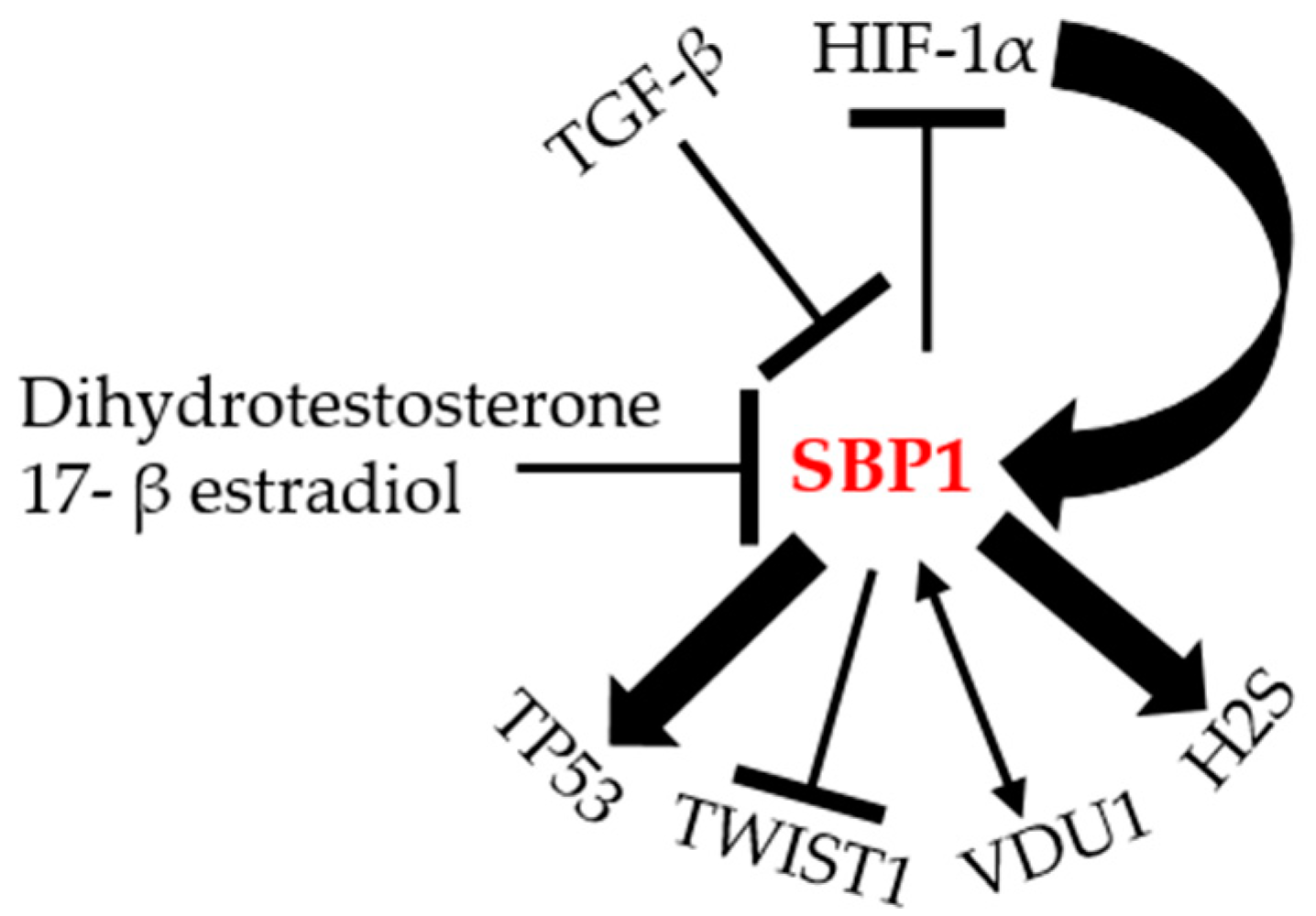
6. The Transcriptional Regulation of SBP1
7. SBP1 Is a Methanethiol Oxidase
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Clark, L.C.; Combs, G.F., Jr.; Turnbull, B.W.; Slate, E.H.; Chalker, D.K.; Chow, J.; Davis, L.S.; Glover, R.A.; Graham, G.F.; Gross, E.G.; et al. Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin. A randomized controlled trial. Nutritional prevention of cancer study group. JAMA 1996, 276, 1957–1963. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.C.; Dalkin, B.; Krongrad, A.; Combs, G.F., Jr.; Turnbull, B.W.; Slate, E.H.; Witherington, R.; Herlong, J.H.; Janosko, E.; Carpenter, D.; et al. Decreased incidence of prostate cancer with selenium supplementation: Results of a double-blind cancer prevention trial. Br. J. Urol. 1998, 81, 730–734. [Google Scholar] [CrossRef] [PubMed]
- Schrauzer, G.N.; White, D.A.; Schneider, C.J. Cancer mortality correlation studies. Iii. Statistical association with dietary selenium intakes. Bioinorg. Chem. 1977, 7, 23–34. [Google Scholar] [CrossRef]
- Shamberger, R.J.; Frost, D.V. Possible protective effect of selenium against human cancer. Can. Med. Assoc. J. 1969, 100, 682. [Google Scholar] [PubMed]
- Brinkman, M.; Reulen, R.C.; Kellen, E.; Buntinx, F.; Zeegers, M.P. Are men with low selenium levels at increased risk of prostate cancer? Eur. J. Cancer 2006, 42, 2463–2471. [Google Scholar] [CrossRef] [PubMed]
- Fortmann, S.P.; Burda, B.U.; Senger, C.A.; Lin, J.S.; Whitlock, E.P. Vitamin and mineral supplements in the primary prevention of cardiovascular disease and cancer: An updated systematic evidence review for the u.S. Preventive services task force. Ann. Intern. Med. 2013, 159, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, H.; Speckmann, B.; Sies, H. Toward understanding success and failures in the use of selenium for cancer prevention. Antioxid. Redox Signal. 2013, 19, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Vinceti, M.; Crespi, C.M.; Malagoli, C.; Del Giovane, C.; Krogh, V. Friend or foe? The current epidemiologic evidence on selenium and human cancer risk. J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2013, 31, 305–341. [Google Scholar] [CrossRef] [PubMed]
- Ip, C. Lessons from basic research in selenium and cancer prevention. J. Nutr. 1998, 128, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.A. Selenium: Epidemiology and basic science. J. Urol. 2004, 171, S50–S53. [Google Scholar] [CrossRef] [PubMed]
- Behne, D.; Kyriakopoulos, A. Mammalian selenium-containing proteins. Annu. Rev. Nutr. 2001, 21, 453–473. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.D.; Uthus, E.O. Dietary folate and selenium affect dimethylhydrazine-induced aberrant crypt formation, global DNA methylation and one-carbon metabolism in rats. J. Nutr. 2003, 133, 2907–2914. [Google Scholar] [CrossRef] [PubMed]
- Ip, C.; Dong, Y.; Ganther, H. New concepts in selenium chemoprevention. Cancer Metastasis Rev. 2002, 21, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, H.; Dong, Y.; Park, Y.M.; Ip, C. Endoplasmic reticulum stress signal mediators are targets of selenium action. Cancer Res. 2005, 65, 9073–9079. [Google Scholar] [CrossRef] [PubMed]
- Bera, S.; De Rosa, V.; Rachidi, W.; Diamond, A.M. Does a role for selenium in DNA damage repair explain apparent controversies in its use in chemoprevention? Mutagenesis 2013, 28, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Kohrle, J. The deiodinase family: Selenoenzymes regulating thyroid hormone availability and action. Cell. Mol. Life Sci. 2000, 57, 1853–1863. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Avery, J.C.; Hoffmann, P.R. Selenium, selenoproteins, and immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [PubMed]
- Turanov, A.A.M.M.; Gladyshev, V.N. Selenium and male reproduction. In Selenium: Its Molecular Biology and Role in Human Health, 3rd ed.; Hatfield, D.L.B.M., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2012; pp. 409–417. [Google Scholar]
- Handy, D.E.L.J. Selenoproteins in cardiovascular redox pathology. In Selenium: Its Molecular Biology and Role in Human Health, 3rd ed.; Hatfield, D.L.B.M., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2012; pp. 249–259. [Google Scholar]
- Kaushal, N.G.U.; Nelson, S.M.; Narayan, V.; Prabhu, K.S. Selenium and inflamation. In Selenium: Its Molecular Biology and Role in Human Health, 3rd ed.; Hatfield, D.L.B.M., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2012; pp. 443–456. [Google Scholar]
- Kudva, A.K.; Shay, A.E.; Prabhu, K.S. Selenium and inflammatory bowel disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G71–G77. [Google Scholar] [CrossRef] [PubMed]
- Moghadaszadeh, B.; Petit, N.; Jaillard, C.; Brockington, M.; Quijano Roy, S.; Merlini, L.; Romero, N.; Estournet, B.; Desguerre, I.; Chaigne, D.; et al. Mutations in sepn1 cause congenital muscular dystrophy with spinal rigidity and restrictive respiratory syndrome. Nat. Genet. 2001, 29, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigo, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, D.L.; Gladyshev, V.N. How selenium has altered our understanding of the genetic code. Mol. Cell. Biol. 2002, 22, 3565–3576. [Google Scholar] [CrossRef] [PubMed]
- Bansal, M.P.; Oborn, C.J.; Danielson, K.G.; Medina, D. Evidence for two selenium-binding proteins distinct from glutathione peroxidase in mouse liver. Carcinogenesis 1989, 10, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Goldberg, M.L.; Pohl, N.M.; Bi, X.; Tong, C.; Xiong, B.; Koh, T.J.; Diamond, A.M.; Yang, W. Functional and physical interaction between the selenium-binding protein 1 (sbp1) and the glutathione peroxidase 1 selenoprotein. Carcinogenesis 2010, 31, 1360–1366. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.W.; Tsui, S.K.; Liew, C.; Lee, C.C.; Waye, M.M.; Fung, K.P. Isolation, characterization, and chromosomal mapping of a novel cdna clone encoding human selenium binding protein. J. Cell. Biochem. 1997, 64, 217–224. [Google Scholar] [CrossRef]
- Yang, M.; Sytkowski, A.J. Differential expression and androgen regulation of the human selenium-binding protein gene hsp56 in prostate cancer cells. Cancer Res. 1998, 58, 3150–3153. [Google Scholar] [PubMed]
- Flemetakis, E.; Agalou, A.; Kavroulakis, N.; Dimou, M.; Martsikovskaya, A.; Slater, A.; Spaink, H.P.; Roussis, A.; Katinakis, P. Lotus japonicus gene ljsbp is highly conserved among plants and animals and encodes a homologue to the mammalian selenium-binding proteins. Mol. Plant Microbe Interact. 2002, 15, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, J.; Fleming, J.; Walker, M.; Harrison, P. Different patterns of regulation of the genes encoding the closely related 56 kda selenium- and acetaminophen-binding proteins in normal tissues and during carcinogenesis. Carcinogenesis 1993, 14, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.Y.; Wang, Y.; Sytkowski, A.J. Human selenium binding protein-1 (hsp56) interacts with vdu1 in a selenium-dependent manner. Biochem. Biophys. Res. Commun. 2009, 379, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Raucci, R.; Colonna, G.; Guerriero, E.; Capone, F.; Accardo, M.; Castello, G.; Costantini, S. Structural and functional studies of the human selenium binding protein-1 and its involvement in hepatocellular carcinoma. Biochim. Biophys. Acta 2011, 1814, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Ying, Q.; Ansong, E.; Diamond, A.M.; Yang, W. A critical role for cysteine 57 in the biological functions of selenium binding protein-1. Int. J. Mol. Sci. 2015, 16, 27599–27608. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Na, X.; Wang, D.; Schoen, S.R.; Messing, E.M.; Wu, G. Ubiquitination of a novel deubiquitinating enzyme requires direct binding to von hippel-lindau tumor suppressor protein. J. Biol. Chem. 2002, 277, 4656–4662. [Google Scholar] [CrossRef] [PubMed]
- Pol, A.; Renkema, G.H.; Tangerman, A.; Winkel, E.G.; Engelke, U.F.; de Brouwer, A.P.M.; Lloyd, K.C.; Araiza, R.S.; van den Heuvel, L.; Omran, H.; et al. Mutations in SELENBP1, encoding a novel human methanethiol oxidase, cause extraoral halitosis. Nat. Genet. 2018, 50, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, D.M.; Copeland, P.R. Mechanism and regulation of selenoprotein synthesis. Annu. Rev. Nutr. 2003, 23, 17–40. [Google Scholar] [CrossRef] [PubMed]
- Bansal, M.P.; Ip, C.; Medina, D. Levels and 75se-labeling of specific proteins as a consequence of dietary selenium concentration in mice and rats. Proc. Soc. Exp. Biol. Med. 1991, 196, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Ding, G.; Gu, C.; Zhou, J.; Kuang, M.; Ji, Y.; He, Y.; Kondo, T.; Fan, J. Decreased selenium-binding protein 1 enhances glutathione peroxidase 1 activity and downregulates hif-1alpha to promote hepatocellular carcinoma invasiveness. Clin. Cancer Res. 2012, 18, 3042–3053. [Google Scholar] [CrossRef] [PubMed]
- Jerome-Morais, A.; Wright, M.E.; Liu, R.; Yang, W.; Jackson, M.I.; Combs, G.F., Jr.; Diamond, A.M. Inverse association between glutathione peroxidase activity and both selenium-binding protein 1 levels and gleason score in human prostate tissue. Prostate 2012, 72, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Ansong, E.; Yang, W.; Diamond, A.M. Molecular cross-talk between members of distinct families of selenium containing proteins. Mol. Nutr. Food Res. 2014, 58, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.M.; Helmke, S.M.; Hunsucker, S.W.; Netea-Maier, R.T.; Chiang, S.A.; Heinz, D.E.; Shroyer, K.R.; Duncan, M.W.; Haugen, B.R. Quantitative and qualitative differences in protein expression between papillary thyroid carcinoma and normal thyroid tissue. Mol. Carcinog. 2006, 45, 613–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Liao, L.; Wan, Y.P.; Li, M.X.; Chen, S.H.; Mo, W.J.; Zhao, Q.L.; Huang, L.F.; Zeng, G.Q. Downregulation of selenium-binding protein 1 is associated with poor prognosis in lung squamous cell carcinoma. World J. Surg. Oncol. 2016, 14, 70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xu, W.; Pan, W.; Wang, N.; Li, G.; Fan, X.; Xu, X.; Shen, S.; Das, U.N. Selenium-binding protein 1 may decrease gastric cellular proliferation and migration. Int. J. Oncol. 2013, 42, 1620–1629. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.J.; Ma, Y.Y.; He, X.J.; Wang, H.J.; Ye, Z.Y.; Tao, H.Q. Suppression of selenium-binding protein 1 in gastric cancer is associated with poor survival. Hum. Pathol. 2011, 42, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.S.; Lee, G.T.; Kim, Y.H.; Kwon, S.Y.; Choi, S.H.; Kim, T.H.; Kwon, T.G.; Yun, S.J.; Kim, I.Y.; Kim, W.J. Decreased selenium-binding protein 1 mrna expression is associated with poor prognosis in renal cell carcinoma. World J. Surg. Oncol. 2014, 12, 288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, Y.E.; Zhang, P.; Liu, F.; Sung, C.J.; Steinhoff, M.M.; Quddus, M.R.; Lawrence, W.D. Progressive loss of selenium-binding protein 1 expression correlates with increasing epithelial proliferation and papillary complexity in ovarian serous borderline tumor and low-grade serous carcinoma. Hum. Pathol. 2010, 41, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.C.; Park, D.C.; Ng, S.K.; Lee, J.Y.; Ni, X.; Ng, W.C.; Bandera, C.A.; Welch, W.R.; Berkowitz, R.S.; Mok, S.C.; et al. Selenium binding protein 1 in ovarian cancer. Int. J. Cancer 2006, 118, 2433–2440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stammer, K.; Edassery, S.L.; Barua, A.; Bitterman, P.; Bahr, J.M.; Hales, D.B.; Luborsky, J.L. Selenium-binding protein 1 expression in ovaries and ovarian tumors in the laying hen, a spontaneous model of human ovarian cancer. Gynecol. Oncol. 2008, 109, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, F.; Younes, M.; Liu, H.; Chen, C.; Yao, Q. Reduced selenium-binding protein 1 in breast cancer correlates with poor survival and resistance to the anti-proliferative effects of selenium. PLoS ONE 2013, 8, e63702. [Google Scholar] [CrossRef] [PubMed]
- Ansong, E.; Ying, Q.; Ekoue, D.N.; Deaton, R.; Hall, A.R.; Kajdacsy-Balla, A.; Yang, W.; Gann, P.H.; Diamond, A.M. Evidence that selenium binding protein 1 is a tumor suppressor in prostate cancer. PLoS ONE 2015, 10, e0127295. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.Y.; Zhou, J.R.; Gao, C.; Feldman, L.; Sytkowski, A.J. Human selenium binding protein-1 (hsp56) is a negative regulator of hif-1alpha and suppresses the malignant characteristics of prostate cancer cells. BMB Rep. 2014, 47, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chen, Y.; Yang, X.; Jiang, Y. Selenium-binding protein 1 is associated with the degree of colorectal cancer differentiation and is regulated by histone modification. Oncol. Rep. 2014, 31, 2506–2514. [Google Scholar] [CrossRef] [PubMed]
- Pohl, N.M.; Tong, C.; Fang, W.; Bi, X.; Li, T.; Yang, W. Transcriptional regulation and biological functions of selenium-binding protein 1 in colorectal cancer in vitro and in nude mouse xenografts. PLoS ONE 2009, 4, e7774. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Chen, C.; Qu, Y.; Xiang, H.; Ai, Q.; Yang, F.; Tan, X.; Zhou, Y.; Jiang, G.; Zhang, Z. Selenium-binding protein 1 in head and neck cancer is low-expression and associates with the prognosis of nasopharyngeal carcinoma. Medicine 2016, 95, e4592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schott, M.; de Jel, M.M.; Engelmann, J.C.; Renner, P.; Geissler, E.K.; Bosserhoff, A.K.; Kuphal, S. Selenium-binding protein 1 is down-regulated in malignant melanoma. Oncotarget 2018, 9, 10445–10456. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kang, H.J.; You, K.T.; Kim, S.H.; Lee, K.Y.; Kim, T.I.; Kim, C.; Song, S.Y.; Kim, H.J.; Lee, C.; et al. Suppression of human selenium-binding protein 1 is a late event in colorectal carcinogenesis and is associated with poor survival. Proteomics 2006, 6, 3466–3476. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Yang, M.; Chan, J.; Sun, T.; Mucci, L.A.; Penney, K.L.; Lee, G.S.; Kantoff, P.W. Association of genetic variations of selenoprotein genes, plasma selenium levels, and prostate cancer aggressiveness at diagnosis. Prostate 2016, 76, 691–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, Q.; Ansong, E.; Diamond, A.M.; Lu, Z.; Yang, W.; Bie, X. Quantitative proteomic analysis reveals that anti-cancer effects of selenium-binding protein 1 in vivo are associated with metabolic pathways. PLoS ONE 2015, 10, e0126285. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.T.; Ding, G.Y.; Yang, X.; Dong, R.Z.; Hu, B.; Zhu, X.D.; Cai, J.B.; Ji, Y.; Shi, G.M.; Shen, Y.H.; et al. Invasive potential of hepatocellular carcinoma is enhanced by loss of selenium-binding protein 1 and subsequent upregulation of cxcr4. Am. J. Cancer Res. 2018, 8, 1040–1049. [Google Scholar] [PubMed]
- Caswell, D.R.; Chuang, C.H.; Ma, R.K.; Winters, I.P.; Snyder, E.L.; Winslow, M.M. Tumor suppressor activity of SELENBP1, a direct nkx2-1 target, in lung adenocarcinoma. Mol. Cancer Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, S.; Ishida, T.; Takeda, T.; Ishii, Y.; Onomura, Y.; Tsukimori, K.; Takechi, S.; Yamaguchi, T.; Uchi, H.; Suzuki, S.O.; et al. Selenium-binding protein 1: Its physiological function, dependence on aryl hydrocarbon receptors, and role in wasting syndrome by 2,3,7,8-tetrachlorodibenzo-p-dioxin. Biochim. Biophys. Acta 2013, 1830, 3616–3624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhan, N.; Dong, W.G. Altered expression of selenium-binding protein 1 in gastric carcinoma and precursor lesions. Med. Oncol. 2011, 28, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.Q.; Yi, H.; Zhang, P.F.; Li, X.H.; Hu, R.; Li, M.Y.; Li, C.; Qu, J.Q.; Deng, X.; Xiao, Z.Q. The function and significance of SELENBP1 downregulation in human bronchial epithelial carcinogenic process. PLoS ONE 2013, 8, e71865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, C.; Wang, X.; Liu, F.; Sung, C.J.; Quddus, M.R.; Lawrence, W.D. The expression of selenium-binding protein 1 is decreased in uterine leiomyoma. Diagn. Pathol. 2010, 5, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroshi, I.; Masayuki, I.; Shosuke, K. Early decreases in pulmonary, hepatic and renal glutathione levels in response to cadmium instillation into rat trachea. J. Appl. Toxicol. 1991, 11, 211–217. [Google Scholar] [CrossRef]
- Wang, Y.; Fang, W.; Huang, Y.; Hu, F.; Ying, Q.; Yang, W.; Xiong, B. Reduction of selenium-binding protein 1 sensitizes cancer cells to selenite via elevating extracellular glutathione: A novel mechanism of cancer-specific cytotoxicity of selenite. Free Radic. Biol. Med. 2015, 79, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Porat, A.; Sagiv, Y.; Elazar, Z. A 56-kda selenium-binding protein participates in intra-golgi protein transport. J. Biol. Chem. 2000, 275, 14457–14465. [Google Scholar] [CrossRef] [PubMed]
- Miyaguchi, K. Localization of selenium-binding protein at the tips of rapidly extending protrusions. Histochem. Cell Biol. 2004, 121, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Glatt, S.J.; Everall, I.P.; Kremen, W.S.; Corbeil, J.; Sasik, R.; Khanlou, N.; Han, M.; Liew, C.C.; Tsuang, M.T. Comparative gene expression analysis of blood and brain provides concurrent validation of SELENBP1 up-regulation in schizophrenia. Proc. Natl. Acad. Sci. USA 2005, 102, 15533–15538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanazawa, T.; Chana, G.; Glatt, S.J.; Mizuno, H.; Masliah, E.; Yoneda, H.; Tsuang, M.T.; Everall, I.P. The utility of SELENBP1 gene expression as a biomarker for major psychotic disorders: Replication in schizophrenia and extension to bipolar disorder with psychosis. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2008, 147B, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Udawela, M.; Money, T.T.; Neo, J.; Seo, M.S.; Scarr, E.; Dean, B.; Everall, I.P. SELENBP1 expression in the prefrontal cortex of subjects with schizophrenia. Transl. Psychiatry 2015, 5, e615. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, T.; Glatt, S.J.; Faraone, S.V.; Hwu, H.G.; Yoneda, H.; Tsuang, M.T. Family-based association study of SELENBP1 in schizophrenia. Schizophr. Res. 2009, 113, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Chau, E.J.; Mostaid, M.S.; Cropley, V.; McGorry, P.; Pantelis, C.; Bousman, C.A.; Everall, I.P. Downregulation of plasma SELENBP1 protein in patients with recent-onset schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 85, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Brauer, A.U.; Savaskan, N.E. Molecular actions of selenium in the brain: Neuroprotective mechanisms of an essential trace element. Rev. Neurosci. 2004, 15, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Du, P.; Lin, S.; Dudley, V.J.; Hernandez, M.R.; Sarthy, V.P. Gene expression changes in retinal muller (glial) cells exposed to elevated pressure. Curr. Eye Res. 2011, 36, 754–767. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qu, D.; An, J.; Yuan, G.; Liu, Y. Integrated microarray analysis provided novel insights to the pathogenesis of glaucoma. Mol. Med. Rep. 2017, 16, 8735–8746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okunuki, Y.; Usui, Y.; Takeuchi, M.; Kezuka, T.; Hattori, T.; Masuko, K.; Nakamura, H.; Yudoh, K.; Usui, M.; Nishioka, K.; et al. Proteomic surveillance of autoimmunity in behcet’s disease with uveitis: Selenium binding protein is a novel autoantigen in behcet’s disease. Exp. Eye Res. 2007, 84, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Krause, I.; Weinberger, A. Behcet’s disease. Curr. Opin. Rheumatol. 2008, 20, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Torrealba, J.R.; Colburn, M.; Golner, S.; Chang, Z.; Scheunemann, T.; Fechner, J.H.; Roenneburg, D.; Hu, H.; Alam, T.; Kim, H.T.; et al. Selenium-binding protein-1 in smooth muscle cells is downregulated in a rhesus monkey model of chronic allograft nephropathy. Am. J. Transplant. 2005, 5, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Scortegagna, M.; Martin, R.J.; Kladney, R.D.; Neumann, R.G.; Arbeit, J.M. Hypoxia-inducible factor-1alpha suppresses squamous carcinogenic progression and epithelial-mesenchymal transition. Cancer Res. 2009, 69, 2638–2646. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Targeting hif-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; De Marzo, A.M.; Laughner, E.; Lim, M.; Hilton, D.A.; Zagzag, D.; Buechler, P.; Isaacs, W.B.; Semenza, G.L.; Simons, J.W. Overexpression of hypoxia-inducible factor 1alpha in common human cancers and their metastases. Cancer Res. 1999, 59, 5830–5835. [Google Scholar] [PubMed]
- Feldser, D.; Agani, F.; Iyer, N.V.; Pak, B.; Ferreira, G.; Semenza, G.L. Reciprocal positive regulation of hypoxia-inducible factor 1alpha and insulin-like growth factor 2. Cancer Res. 1999, 59, 3915–3918. [Google Scholar] [PubMed]
- Talks, K.L.; Turley, H.; Gatter, K.C.; Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J.; Harris, A.L. The expression and distribution of the hypoxia-inducible factors hif-1alpha and hif-2alpha in normal human tissues, cancers, and tumor-associated macrophages. Am. J. Pathol. 2000, 157, 411–421. [Google Scholar] [CrossRef]
- Birner, P.; Schindl, M.; Obermair, A.; Plank, C.; Breitenecker, G.; Oberhuber, G. Overexpression of hypoxia-inducible factor 1alpha is a marker for an unfavorable prognosis in early-stage invasive cervical cancer. Cancer Res. 2000, 60, 4693–4696. [Google Scholar] [PubMed]
- Burri, P.; Djonov, V.; Aebersold, D.M.; Lindel, K.; Studer, U.; Altermatt, H.J.; Mazzucchelli, L.; Greiner, R.H.; Gruber, G. Significant correlation of hypoxia-inducible factor-1alpha with treatment outcome in cervical cancer treated with radical radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2003, 56, 494–501. [Google Scholar] [CrossRef]
- Bos, R.; van der Groep, P.; Greijer, A.E.; Shvarts, A.; Meijer, S.; Pinedo, H.M.; Semenza, G.L.; van Diest, P.J.; van der Wall, E. Levels of hypoxia-inducible factor-1alpha independently predict prognosis in patients with lymph node negative breast carcinoma. Cancer 2003, 97, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Schindl, M.; Schoppmann, S.F.; Samonigg, H.; Hausmaninger, H.; Kwasny, W.; Gnant, M.; Jakesz, R.; Kubista, E.; Birner, P.; Oberhuber, G.; et al. Overexpression of hypoxia-inducible factor 1alpha is associated with an unfavorable prognosis in lymph node-positive breast cancer. Clin. Cancer Res. 2002, 8, 1831–1837. [Google Scholar] [PubMed]
- Birner, P.; Schindl, M.; Obermair, A.; Breitenecker, G.; Oberhuber, G. Expression of hypoxia-inducible factor 1alpha in epithelial ovarian tumors: Its impact on prognosis and on response to chemotherapy. Clin. Cancer Res. 2001, 7, 1661–1668. [Google Scholar] [PubMed]
- Sivridis, E.; Giatromanolaki, A.; Gatter, K.C.; Harris, A.L.; Koukourakis, M.I.; Tumor and Angiogenesis Research Group. Association of hypoxia-inducible factors 1alpha and 2alpha with activated angiogenic pathways and prognosis in patients with endometrial carcinoma. Cancer 2002, 95, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Tanaka, S.; Hiyama, T.; Ito, M.; Kitadai, Y.; Sumii, M.; Haruma, K.; Chayama, K. Hypoxia-inducible factor-1alpha expression and angiogenesis in gastrointestinal stromal tumor of the stomach. Oncol. Rep. 2003, 10, 797–802. [Google Scholar] [PubMed]
- Birner, P.; Gatterbauer, B.; Oberhuber, G.; Schindl, M.; Rossler, K.; Prodinger, A.; Budka, H.; Hainfellner, J.A. Expression of hypoxia-inducible factor-1 alpha in oligodendrogliomas: Its impact on prognosis and on neoangiogenesis. Cancer 2001, 92, 165–171. [Google Scholar] [CrossRef]
- Beasley, N.J.; Leek, R.; Alam, M.; Turley, H.; Cox, G.J.; Gatter, K.; Millard, P.; Fuggle, S.; Harris, A.L. Hypoxia-inducible factors hif-1alpha and hif-2alpha in head and neck cancer: Relationship to tumor biology and treatment outcome in surgically resected patients. Cancer Res. 2002, 62, 2493–2497. [Google Scholar] [PubMed]
- Volm, M.; Koomagi, R. Hypoxia-inducible factor (hif-1) and its relationship to apoptosis and proliferation in lung cancer. Anticancer Res. 2000, 20, 1527–1533. [Google Scholar] [PubMed]
- Chen, G.; Wang, H.; Miller, C.T.; Thomas, D.G.; Gharib, T.G.; Misek, D.E.; Giordano, T.J.; Orringer, M.B.; Hanash, S.M.; Beer, D.G. Reduced selenium-binding protein 1 expression is associated with poor outcome in lung adenocarcinomas. J. Pathol. 2004, 202, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Barr, L.A.; Calvert, J.W. Discoveries of hydrogen sulfide as a novel cardiovascular therapeutic. Circ. J. 2014, 78, 2111–2118. [Google Scholar] [CrossRef] [PubMed]
- Kohl, J.B.; Mellis, A.T.; Schwarz, G. Homeostatic impact of sulfite and hydrogen sulfide on cysteine catabolism. Br. J. Pharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C.; Ransy, C.; Modis, K.; Andriamihaja, M.; Murghes, B.; Coletta, C.; Olah, G.; Yanagi, K.; Bouillaud, F. Regulation of mitochondrial bioenergetic function by hydrogen sulfide. Part i. Biochemical and physiological mechanisms. Br. J. Pharmacol. 2014, 171, 2099–2122. [Google Scholar] [CrossRef] [PubMed]
- Bos, E.M.; van Goor, H.; Joles, J.A.; Whiteman, M.; Leuvenink, H.G. Hydrogen sulfide: Physiological properties and therapeutic potential in ischaemia. Br. J. Pharmacol. 2015, 172, 1479–1493. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Wang, R. Hydrogen sulfide-based therapeutics: Exploiting a unique but ubiquitous gasotransmitter. Nat. Rev. Drug Discov. 2015, 14, 329–345. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elhodaky, M.; Diamond, A.M. Selenium-Binding Protein 1 in Human Health and Disease. Int. J. Mol. Sci. 2018, 19, 3437. https://doi.org/10.3390/ijms19113437
Elhodaky M, Diamond AM. Selenium-Binding Protein 1 in Human Health and Disease. International Journal of Molecular Sciences. 2018; 19(11):3437. https://doi.org/10.3390/ijms19113437
Chicago/Turabian StyleElhodaky, Mostafa, and Alan M. Diamond. 2018. "Selenium-Binding Protein 1 in Human Health and Disease" International Journal of Molecular Sciences 19, no. 11: 3437. https://doi.org/10.3390/ijms19113437
APA StyleElhodaky, M., & Diamond, A. M. (2018). Selenium-Binding Protein 1 in Human Health and Disease. International Journal of Molecular Sciences, 19(11), 3437. https://doi.org/10.3390/ijms19113437