Functional Identification of Salt-Stress-Related Genes Using the FOX Hunting System from Ipomoea pes-caprae

 ,
,
Abstract
:1. Introduction
2. Results
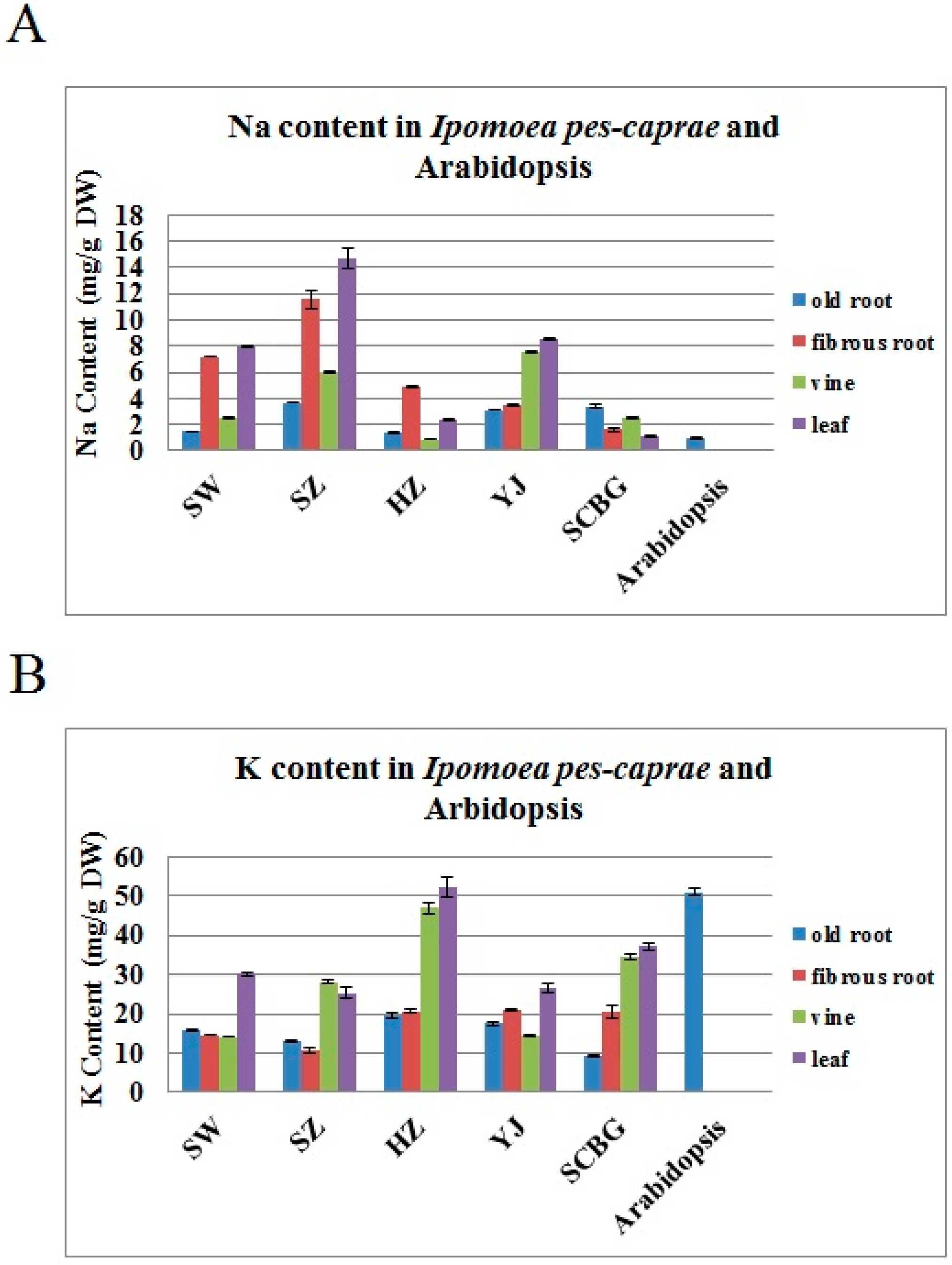
2.1. The Na+ and K+ Contents in I. pes-caprae
2.2. Construction of the cDNA Expression Library
2.3. Functional Screening of the cDNA Library for Mining the Candidate IpSR Genes
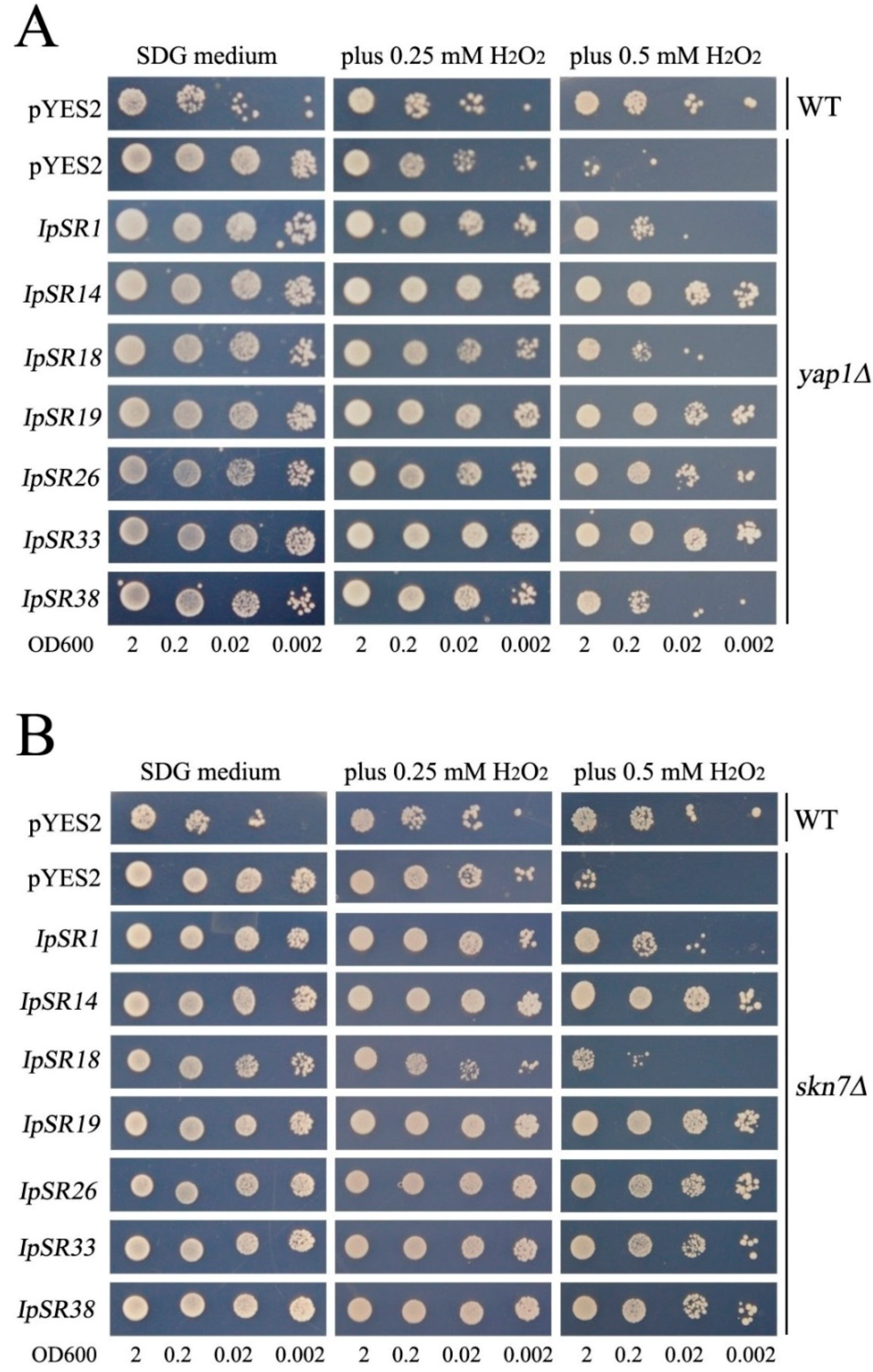
2.4. Retransformation and Salt Tolerance Confirmation in Yeast
2.5. H2O2 Sensitivity Assays in Yeast
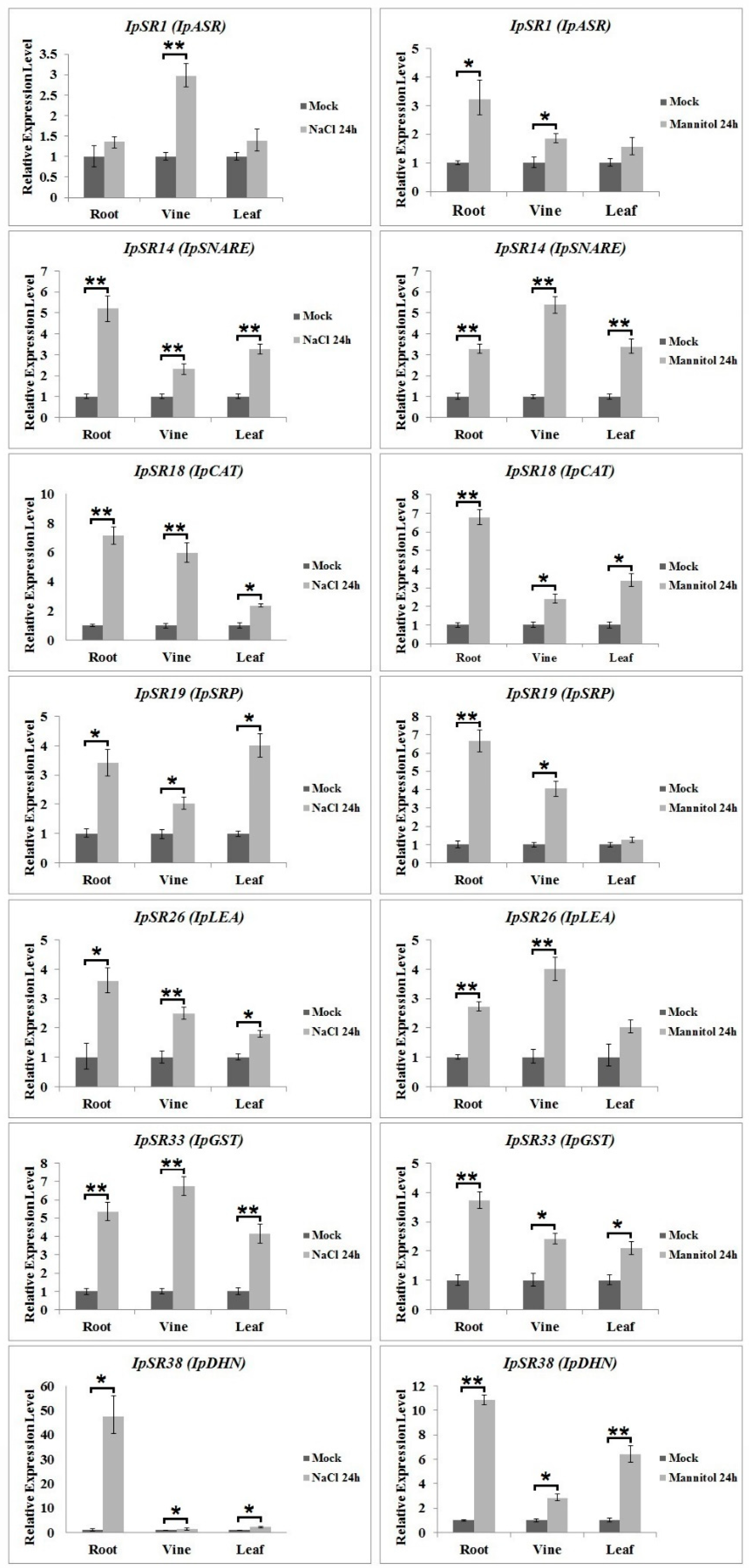
2.6. qRT-PCR Analysis of Candidate Genes under Salt and Osmotic Stress
3. Discussion
3.1. Plant Abscisic Acid, Stress, and Ripening-Induced (ASR) Proteins
3.2. LEA Proteins
3.3. SNARE Proteins
3.4. Catalase and Glutathione S-Transferase
3.5. Other Stress-Responsive Proteins
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Stress Treatments
4.2. Measurement of Na+ and K+ Contents in I. pes-caprae
4.3. cDNA Library Construction and Quality Examination Assays
4.4. Yeast Mutant Strains and Functional Screening
4.5. Salt and H2O2 Sensitivity Assays in Yeast Cells
4.6. Quantitative Reverse Transcription (qRT)-PCR Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FOX | Full-length cDNA Over-eXpressor |
| SR | Salt-stress Related |
| qRT-PCR | Quantitative Reverse Transcript-Polymerase Chain Reaction |
| ROS | Reactive Oxygen Species |
| SCBG | South China Botanical Garden |
| NCBI | National Center for Biotechnology Information |
| RIN | RNA Integrity Number |
| ORF | Open Reading Frame |
| LB | Luria-Bertani |
| MS | Murashige and Skoog |
| SDG | Synthetic Dropout/Galactose |
| OX | Over eXpression |
References
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2014, 65, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Das, P.; Parida, A.K.; Agarwal, P.K. Proteomics, metabolomics, and ionomics perspectives of salinity tolerance in halophytes. Front. Plant Sci. 2015, 6, 537. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Devall, M.S; Thien, L.B. Factors influencing the reproductive success of Ipomoea pes-caprae (Convolvulaceae) around the gulf of Mexico. Am. J. Bot. 1989, 76, 1821–1831. [Google Scholar] [CrossRef]
- Miryeganeh, M.; Takayama, K.; Tateishi, Y.; Kajita, T. Long-distance dispersal by sea-drifted seeds has maintained the global distribution of Ipomoea pes-caprae subsp. brasiliensis (Convolvulaceae). PLoS ONE 2014, 9, e91836. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, P.Y.; Liu, N.; Zhang, W.W.; Wang, J.; Jian, S.G. Biological and ecophysiological characteristics of a beach plant Ipomoea pes-caprae. J. Hunan Univ. Sci. Technol. (Nat. Sci. Ed.) 2011, 26, 117–121. [Google Scholar]
- Urano, K.; Kurihara, Y.; Seki, M.; Shinozaki, K. ‘Omics’ analyses of regulatory networks in plant abiotic stress responses. Curr. Opin. Plant. Biol. 2010, 3, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Higuchi-Takeuchi, M.; Kondou, Y.; Ichikawa, T.; Matsui, M. Full-length cDNA overexpressor gene hunting system (FOX hunting system). Methods Mol. Biol. 2011, 678, 77–89. [Google Scholar]
- Sakurai, T.; Kondou, Y.; Akiyama, K.; Kurotani, A.; Higuchi, M.; Ichikawa, T.; Kuroda, H.; Kusano, M.; Mori, M.; Saitou, T.; et al. RiceFOX: A database of Arabidopsis mutant lines overexpressing rice full-length cDNA that contains a wide range of trait information to facilitate analysis of gene function. Plant Cell Physiol. 2011, 52, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zong, J.; Tan, Z.; Li, L.; Hu, B.; Chen, C.; Chen, J.; Liu, J. Systematic mining of salt-tolerant genes in halophyte-Zoysia matrella through cDNA expression library screening. Plant Physiol. Biochem. 2015, 89, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, C.; Tan, Z.; Liu, J.; Zhuang, L.; Yang, Z.; Huang, B. Functional identification and characterization of genes cloned from halophyte seashore paspalum conferring salinity and cadmium tolerance. Front. Plant Sci. 2016, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, L.; Lu, J.; Cheng, Y.; You, Q.; Wang, L.; Song, X.; Zhou, X.; Jiao, Y. Large-scale investigation of soybean gene functions by overexpressing a full-Length soybean cDNA library in Arabidopsis. Front. Plant Sci. 2018, 9, 631. [Google Scholar] [CrossRef] [PubMed]
- Lippuner, V.; Cyert, M.S.; Gasser, C.S. Two classes of plant cDNA clones differentially complement yeast calcineurin mutants and increase salt tolerance of wild-type yeast. J. Biol. Chem. 1996, 271, 12859–12866. [Google Scholar] [CrossRef] [PubMed]
- Piao, H.L.; Pih, K.T.; Lim, J.H.; Kang, S.G.; Jin, J.B.; Kim, S.H.; Hwang, I. An Arabidopsis GSK3/shaggy-like gene that complements yeast salt stress-sensitive mutants is induced by NaCl and abscisic acid. Plant Physiol. 1999, 119, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.K.; Pardo, J.M.; Takeda, S.; Bressan, R.A.; Hasegawa, P.M. Tobacco and Arabidiopsis SLT1 mediate salt tolerance of yeast. Plant Mol. Biol. 2001, 45, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Obata, T.; Kitamoto, H.K.; Nakamura, A.; Fukuda, A.; Tanaka, Y. Rice shaker potassium channel OsKAT1 confers tolerance to salinity stress on yeast and rice cells. Plant Physiol. 2007, 144, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Li, J.; Sun, X.; Liu, Y.; Wang, X.; Zhang, H.; Pan, H. Exploration for the salinity tolerance-related genes from xero-halophyte Atriplex canescens exploiting yeast functional screening system. Int. J. Mol. Sci. 2017, 18, E2444. [Google Scholar] [CrossRef] [PubMed]
- Kamakura, M.; Furukawa, A. Compensatory function for water transport by adventitious roots of Ipomoea pes-caprae. J. Plant Res. 2009, 122, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Quintero, F.J.; Blatt, M.R.; Pardo, J.M. Functional conservation between yeast and plant endosomal Na+/H+ antiporters. FEBS Lett. 2000, 471, 224–228. [Google Scholar] [CrossRef]
- Zhou, Y.; Yin, X.; Duan, R.; Hao, G.; Guo, J.; Jiang, X. SpAHA1 and SpSOS1 coordinate in transgenic yeast to improve salt tolerance. PLoS ONE 2015, 10, e0137447. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Møller, A.L.; Kristiansen, K.A.; Schulz, A.; Møller, I.M.; Schjoerring, J.K.; Jahn, T.P. Specific aquaporins facilitate the diffusion of hydrogen peroxide across membranes. J. Biol. Chem. 2007, 282, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Ventura, Y.; Eshel, A.; Pasternak, D.; Sagi, M. The development of halophyte-based agriculture: Past and present. Ann. Bot. 2015, 115, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Reimann, C.; Breckle, S.W. Salt tolerance and ion relations of Salsola kali L.: Differences between ssp. tragus (L.) Nyman and ssp. ruthenica (Iljin) Soó. New Phytol. 1995, 130, 37–45. [Google Scholar] [CrossRef]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt tolerance and crop potential of halophytes. CRC Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Mishra, A.; Tanna, B. Halophytes: Potential resources for salt stress tolerance genes and promoters. Front. Plant Sci. 2017, 8, 829. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K.; Hasegawa, P.M.; Bressan, R.A. Molecular aspects of osmotic stress in plants. CRC Crit. Rev. Plant Sci. 1997, 16, 253–277. [Google Scholar] [CrossRef]
- Xiong, L.; Zhu, J.K. Molecular and genetic aspects of plant responses to osmotic stress. Plant Cell Environ. 2002, 25, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.X.; Yang, X.; Li, X.L.; Yu, X.D.; Li, Q.L. The SlASR gene cloned from the extreme halophyte Suaeda liaotungensis K. enhances abiotic stress tolerance in transgenic Arabidopsis thaliana. Gene 2014, 549, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Jha, B.; Lal, S.; Tiwari, V.; Yadav, S.K.; Agarwal, P.K. The SbASR-1 gene cloned from an extreme halophyte Salicornia brachiata enhances salt tolerance in transgenic tobacco. Mar. Biotechnol. (NY) 2012, 14, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, P.G.; Carrari, F. ASR1 transcription factor and its role in metabolism. Plant Signal. Behav. 2015, 10, e992751. [Google Scholar] [CrossRef] [PubMed]
- Shkolnik, D.; Bar-Zvi, D. Tomato ASR1 abrogates the response to abscisic acid and glucose in Arabidopsis by competing with ABI4 for DNA binding. Plant Biotechnol. J. 2008, 6, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Candat, A.; Paszkiewicz, G.; Neveu, M.; Gautier, R.; Logan, D.C.; Avelange-Macherel, M.H.; Macherel, D. The ubiquitous distribution of late embryogenesis abundant proteins across cell compartments in Arabidopsis offers tailored protection against abiotic stress. Plant Cell 2014, 26, 3148–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Song, Q.; Li, D.; Yang, X.; Li, D. Multifunctional roles of plant dehydrins in response to environmental stresses. Front. Plant Sci. 2017, 8, 1018. [Google Scholar] [CrossRef] [PubMed]
- Hincha, D.K.; Thalhammer, A. LEA proteins: IDPs with versatile functions in cellular dehydration tolerance. Biochem. Soc. Trans. 2012, 40, 1000–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosová, K.; Vítámvás, P.; Prášil, I.T. Wheat and barley dehydrins under cold, drought, and salinity—What can LEA-II proteins tell us about plant stress response? Front. Plant Sci. 2014, 5, 343. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, R.; Sutter, J.U.; Blatt, M.R. A new catch in the SNARE. Trends Plant Sci. 2004, 9, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Grefen, C.; Blatt, M.R. SNAREs--molecular governors in signalling and development. Curr. Opin. Plant Biol. 2008, 11, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Tarte, V.N.; Seok, H.Y.; Woo, D.H.; Le, D.H.; Tran, H.T.; Baik, J.W.; Kang, I.S.; Lee, S.Y.; Chung, T.; Moon, Y.H. Arabidopsis Qc-SNARE gene AtSFT12 is involved in salt and osmotic stress responses and Na+ accumulation in vacuoles. Plant Cell Rep. 2015, 34, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Yadav, N.S.; Tiwari, V.; Agarwal, P.K.; Jha, B. A SNARE-Like superfamily protein SbSLSP from the halophyte Salicornia brachiata confers salt and drought tolerance by maintaining membrane stability, K+/Na+ ratio, and antioxidant machinery. Front. Plant Sci. 2016, 7, 737. [Google Scholar] [CrossRef] [PubMed]
- Nisa, Z.U.; Mallano, A.I.; Yu, Y.; Chen, C.; Duan, X.; Amanullah, S.; Kousar, A.; Baloch, A.W.; Sun, X.; Tabys, D.; et al. GsSNAP33, a novel glycine soja SNAP25-type protein gene: Improvement of plant salt and drought tolerances in transgenic Arabidopsis thaliana. Plant Physiol. Biochem. 2017, 119, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, D.; Banerjee, M.; Bihani, S.C.; Ballal, A. A salt-inducible Mn-catalase (KatB) protects cyanobacterium from oxidative stress. Plant Physiol. 2016, 170, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Li, C.; Zhang, Y.; He, Q.; Daud, M.K.; Chen, J.; Zhu, S. Glutathione S-transferase gene family in Gossypium raimondii and G. arboreum: Comparative genomic study and their expression under salt stress. Front. Plant Sci. 2016, 7, 139. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Cummins, I.; Cole, D.J.; Edwards, R. Glutathione-mediated detoxification systems in plants. Curr. Opin. Plant Biol. 1998, 1, 258–266. [Google Scholar] [CrossRef]
- Jha, B.; Sharma, A.; Mishra, A. Expression of SbGSTU (tau class glutathione S-transferase) gene isolated from Salicornia brachiata in tobacco for salt tolerance. Mol. Biol. Rep. 2011, 38, 4823–4832. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Asif, M.H.; Chakrabarty, D.; Tripathi, R.D.; Dubey, R.S.; Trivedi, P.K. Expression of a rice Lambda class of glutathione S-transferase, OsGSTL2, in Arabidopsis provides tolerance to heavy metal and other abiotic stresses. J. Hazard Mater. 2013, 248–249, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.; Lam, H.M. A putative lambda class glutathione S-transferase enhances plant survival under salinity stress. Plant Cell Physiol. 2014, 55, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xing, X.J.; Tian, Y.S.; Peng, R.H.; Xue, Y.; Zhao, W.; Yao, Q.H. Transgenic Arabidopsis plants expressing tomato glutathione S-transferase showed enhanced resistance to salt and drought stress. PLoS ONE 2015, 10, e0136960. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Muramoto, K.; Takada, R.; Nakagawa, S.; Shigeoka, S.; Yoshimura, K. Modulation of NADH levels by Arabidopsis nudix hydrolases, AtNUDX6 and 7, and the respective proteins themselves play distinct roles in the regulation of various cellular responses involved in biotic/abiotic stresses. Plant Cell Physiol. 2016, 57, 1295–1308. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Chang, S.H.; Cho, K.C.; Kim, S.; Park, S.; Lee, A.Y.; Jiang, H.L.; Kim, H.J.; Lee, S.; Yu, K.N.; et al. Endoplasmic reticulum-Golgi intermediate compartment protein 3 knockdown suppresses lung cancer through endoplasmic reticulum stress-induced autophagy. Oncotarget 2016, 7, 65335–65347. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.J.; Han, X.X.; Yin, L.L.; Xing, M.Q.; Xu, Z.H.; Xue, H.W. Arabidopsis PROTEASOME REGULATOR1 is required for auxin-mediated suppression of proteasome activity and regulates auxin signalling. Nat. Commun. 2016, 7, 11388. [Google Scholar] [CrossRef] [PubMed]
- Salas-Muñoz, S.; Rodríguez-Hernández, A.A.; Ortega-Amaro, M.A.; Salazar-Badillo, F.B.; Jiménez-Bremont, J.F. Arabidopsis AtDjA3 null mutant shows increased sensitivity to abscisic acid, salt, and osmotic stress in germination and post-germination stages. Front. Plant Sci. 2016, 7, 220. [Google Scholar] [CrossRef] [PubMed]
- Engqvist, M.K.; Schmitz, J.; Gertzmann, A.; Florian, A.; Jaspert, N.; Arif, M.; Balazadeh, S.; Mueller-Roeber, B.; Fernie, A.R.; Maurino, V.G. GLYCOLATE OXIDASE3, a glycolate oxidase homolog of yeast l-Lactate cytochrome c oxidoreductase, supports l-Lactate oxidation in roots of Arabidopsis. Plant Physiol. 2015, 169, 1042–1061. [Google Scholar] [CrossRef] [PubMed]
- Quan, S.; Yang, P.; Cassin-Ross, G.; Kaur, N.; Switzenberg, R.; Aung, K.; Li, J.; Hu, J. Proteome analysis of peroxisomes from etiolated Arabidopsis seedlings identifies a peroxisomal protease involved in β-oxidation and development. Plant Physiol. 2013, 163, 1518–1538. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Nakazawa, M.; Kawashima, M.; Iizumi, H.; Kuroda, H.; Kondou, Y.; Tsuhara, Y.; Suzuki, K.; Ishikawa, A.; Seki, M.; et al. The FOX hunting system: An alternative gain-of-function gene hunting technique. Plant J. 2006, 48, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Hakata, M.; Amano, K.; Miyao, A.; Toki, N.; Kajikawa, M.; Pang, J.; Higashi, N.; Ando, S.; Toki, S.; et al. A genome-wide gain-of function analysis of rice genes using the FOX-hunting system. Plant Mol. Biol. 2007, 65, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Higashi, Y.; Ohama, N.; Ishikawa, T.; Katori, T.; Shimura, A.; Kusakabe, K.; Yamaguchi-Shinozaki, K.; Ishida, J.; Tanaka, M.; Seki, M.; et al. HsfA1d, a protein identified via FOX hunting using Thellungiella salsuginea cDNAs improves heat tolerance by regulating heat-stress-responsive gene expression. Mol. Plant 2013, 6, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Ariga, H.; Tanaka, T.; Ono, H.; Sakata, Y.; Hayashi, T.; Taji, T. CSP41b, a protein identified via FOX hunting using Eutrema salsugineum cDNAs, improves heat and salinity stress tolerance in transgenic Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 464, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Rigó, G.; Valkai, I.; Faragó, D.; Kiss, E.; Van Houdt, S.; Van de Steene, N.; Hannah, M.A.; Szabados, L. Gene mining in halophytes: Functional identification of stress tolerance genes in Lepidium crassifolium. Plant Cell Environ. 2016, 39, 2074–2084. [Google Scholar] [CrossRef] [PubMed]
- Higuchi-Takeuchi, M.; Matsui, M. Screening for gene function using the FOX (full-length cDNA overexpressor gene) hunting system. Methods Mol. Biol. 2014, 1056, 201–210. [Google Scholar] [PubMed]
- Abe, K.; Ichikawa, H. Gene overexpression resources in cereals for functional genomics and discovery of useful genes. Front. Plant Sci. 2016, 7, 1359. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Woods, R.A. Transformation of yeast by the LiAc/SS carrier DNA/PEG method. Methods Mol. Biol. 2006, 313, 107–120. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone | GenBank Accession No. | Functional Annotation | Length of cDNAs and Proteins Encoded by the Longest ORFs |
|---|---|---|---|
| IpSR1 (IpASR) | MF680587 | putative ripening protein, abscisic acid, stress, and ripening-induced protein (ASR) | 962 bp, 215 aa |
| IpSR2 (IpATPD) | MF680588 | ATP synthase delta chain, chloroplastic | 921 bp, 250 aa |
| IpSR3 (IpNUD) | MF680589 | nudix hydrolase, chloroplastic | 769 bp, 168 aa |
| IpSR4 (IpTSJT1-1) | MF680590 | stem-specific protein TSJT1-like | 1260 bp, 251 aa |
| IpSR5 (IpCDI) | MF680591 | contact-dependent growth inhibition (CDI)-like protein | 1262 bp, 277 aa |
| IpSR6 (IpFBP) | MF680592 | F-box protein At5g46170-like | 1538 bp, 388 aa |
| IpSR7 (IpCAB21) | MF680593 | chlorophyll a-b binding protein 21, chloroplastic | 1013 bp, 267 aa |
| IpSR8 (IpERVT) | MF680594 | endoplasmic reticulum vesicle transporter, C-terminal | 1428 bp, 386 aa |
| IpSR9 (IpTSJT1-2) | MF680595 | stem-specific protein TSJT1-like | 1196 bp, 237 aa |
| IpSR10 (IpPI1) | MF680596 | probable proteasome inhibitor | 1115 bp, 343 aa |
| IpSR11 (IpFRK) | MF680597 | fructokinase | 1312 bp, 324 aa |
| IpSR12 (IpPABP) | MF680598 | polyadenylate-binding protein RBP47B′ isoform X1 | 1598 bp, 423 aa |
| IpSR13 (IpTC) | MF680599 | probable tocopherol cyclase, chloroplastic | 2007 bp, 487 aa |
| IpSR14 (IpSNARE) | MF680600 | hypothetical protein, Syntaxin/t-SNARE family protein | 1743 bp, 346 aa |
| IpSR15 (IpPT) | MF680601 | probable inorganic phosphate transporter 1-3 | 1918 bp, 540 aa |
| IpSR16 (IpAHCY) | MF680602 | adenosylhomocysteinase 1 | 1844 bp, 485 aa |
| IpSR17 (IpPUP) | MF680603 | peptide upstream protein | 2123 bp, 453 aa |
| IpSR18 (IpCAT) | MF680604 | catalase | 1758 bp, 492 aa |
| IpSR19 (IpSRP) | MF680605 | stress responsive alpha-beta barrel domain protein | 847 bp, 221 aa |
| IpSR20 (IpRPB7) | MF680606 | DNA-directed RNA polymerase II subunit RPB7 | 976 bp, 177 aa |
| IpSR21 (IpUSP3) | MF680607 | ubiquitin carboxyl-terminal hydrolase 3 | 1536 bp, 368 aa |
| IpSR22 (IpRPS25) | MF680608 | 40S ribosomal protein S25 | 557 bp, 108 aa |
| IpSR23 (IpPI2) | MF680609 | probable proteasome inhibitor | 1116 bp, 300 aa |
| IpSR24 (IpHP1) | MF680610 | hypothetical protein AT3G52710 | 997 bp, 225 aa |
| IpSR25 (IpPMM) | MF680611 | phosphomannomutase | 1048 bp, 246 aa |
| IpSR26 (IpLEA) | MF680612 | desiccation-related protein At2g46140 | 1392 bp, 313 aa |
| IpSR27 (IpLRRK) | MF680613 | protein kinase superfamily protein | 1732 bp, 385 aa |
| IpSR28 (IpDNAJ) | MF765747 | dnaj protein-like protein | 1595 bp, 428 aa |
| IpSR29 (IpPSK) | MF680614 | phytosulfokines-like | 721 bp, 81 aa |
| IpSR30 (IpPSCXI) | MF680615 | photosystem I reaction center subunit XI, chloroplastic | 803 bp, 218 aa |
| IpSR31 (IpSCP) | MF680616 | sugar carrier protein C | 1877 bp, 528 aa |
| IpSR32 (IpHRGP) | MF680617 | hydroxyproline-rich glycoprotein family protein | 930 bp, 215 aa |
| IpSR33 (IpGST) | MF680618 | glutathione S-transferase L3-like isoform X1 | 1022 bp, 234 aa |
| IpSR34 (IpABAH) | MF680619 | abscisic acid 8′-hydroxylase 4 | 1748 bp, 465 aa |
| IpSR35 (IpSR45a) | MF680620 | serine/arginine-rich splicing factor SR45a isoform X2 | 1106 bp, 243 aa |
| IpSR36 (IpFMT) | MF680621 | quercetin 3-O-methyltransferase 1 | 1277 bp, 356 aa |
| IpSR37 (IpGBP) | MF680622 | guanine nucleotide-binding protein subunit beta-like protein | 1253 bp, 326 aa |
| IpSR38 (IpDHN) | KX426069 | dehydrin | 983 bp, 217 aa |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Zhang, H.; Zheng, J.-X.; Mo, H.; Xia, K.-F.; Jian, S.-G. Functional Identification of Salt-Stress-Related Genes Using the FOX Hunting System from Ipomoea pes-caprae. Int. J. Mol. Sci. 2018, 19, 3446. https://doi.org/10.3390/ijms19113446
Zhang M, Zhang H, Zheng J-X, Mo H, Xia K-F, Jian S-G. Functional Identification of Salt-Stress-Related Genes Using the FOX Hunting System from Ipomoea pes-caprae. International Journal of Molecular Sciences. 2018; 19(11):3446. https://doi.org/10.3390/ijms19113446
Chicago/Turabian StyleZhang, Mei, Hui Zhang, Jie-Xuan Zheng, Hui Mo, Kuai-Fei Xia, and Shu-Guang Jian. 2018. "Functional Identification of Salt-Stress-Related Genes Using the FOX Hunting System from Ipomoea pes-caprae" International Journal of Molecular Sciences 19, no. 11: 3446. https://doi.org/10.3390/ijms19113446
APA StyleZhang, M., Zhang, H., Zheng, J. -X., Mo, H., Xia, K. -F., & Jian, S. -G. (2018). Functional Identification of Salt-Stress-Related Genes Using the FOX Hunting System from Ipomoea pes-caprae. International Journal of Molecular Sciences, 19(11), 3446. https://doi.org/10.3390/ijms19113446