What Is the Evidence that Parkinson’s Disease Is a Prion Disorder, Which Originates in the Gut?



Abstract
:
1. Introduction
2. The Hypothesis of Prion-Like Propagation of A-Synuclein within the Gut-Brain Axis
3. Evidence in PD Patients
4. Evidence from Experimental Models
5. Does Parkinson’s Disease Actually Originate in the Gut?
6. Concluding Remarks
Funding
Conflicts of Interest
Abbreviations
| CNS | Central nervous system |
| DMV | Dorsal motor nucleus of the vagus |
| EECs | Enteroendocrine cells |
| ENS | Enteric nervous system |
| GI | Gastrointestinal |
| IML | Intermediolateral cell column |
| KYN | Kynurenine |
| LAG3 | Lymphocyte-activation gene 3 |
| LBs | Lewy bodies |
| LNs | Lewy neuritis |
| LP | Lewy pathology |
| LPS | Lipopolysaccharide |
| PANS | Peripheral autonomic nervous system |
| PFFs | Preformed fibrillar α-synuclein assemblies |
| PD | Parkinson’s disease |
| SCFAs | Short chain fatty acids |
| SIBO | Small intestinal overgrowth |
| SNpc | Substantia nigra pars compacta |
| TNTs | Tunneling nanotubes |
| VM | Ventral mesencephalic |
References
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef]
- Poirier, A.A.; Aubé, B.; Côté, M.; Morin, N.; Di Paolo, T.; Soulet, D. Gastrointestinal Dysfunctions in Parkinson’s Disease: Symptoms and Treatments. Parkinsons Dis. 2016, 2016, 6762528. [Google Scholar] [CrossRef] [PubMed]
- Stirpe, P.; Hoffman, M.; Badiali, D.; Colosimo, C. Constipation: An emerging risk factor for Parkinson’s disease? Eur. J. Neurol. 2016, 23, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Cersosimo, M.G.; Benarroch, E.E. Pathological correlates of gastrointestinal dysfunction in Parkinson’s disease. Neurobiol. Dis. 2012, 46, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; del Tredici, K.; Rüb, U.; de Vos, R.A.; Jansen Steur, E.N.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Saito, Y.; Shioya, A.; Sano, T.; Sumikura, H.; Murata, M.; Murayama, S. Lewy body pathology involves the olfactory cells in Parkinson’s disease and related disorders. Mov. Disord. 2016, 31, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, K.; Takahashi, H.; Takeda, S.; Ohama, E.; Ikuta, F. Parkinson’s disease: The presence of Lewy bodies in Auerbach’s and Meissner’s plexuses. Acta Neuropathol. 1988, 76, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Del Tredici, K.; Hawkes, C.H.; Ghebremedhin, E.; Braak, H. Lewy pathology in the submandibular gland of individuals with incidental Lewy body disease and sporadic Parkinson’s disease. Acta Neuropathol. 2010, 119, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; del Tredici, K. Invited Article: Nervous system pathology in sporadic Parkinson disease. Neurology 2008, 70, 1916–1925. [Google Scholar] [CrossRef] [PubMed]
- Brundin, P.; Ma, J.; Kordower, J.H. How strong is the evidence that Parkinson’s disease is a prion disorder? Curr. Opin. Neurol. 2016, 29, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Kordower, J.H.; Chu, Y.; Hauser, R.A.; Freeman, T.B.; Olanow, C.W. Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson’s disease. Nat. Med. 2008, 14, 504–506. [Google Scholar] [CrossRef] [PubMed]
- Kordower, J.H.; Chu, Y.; Hauser, R.A.; Olanow, C.W.; Freeman, T.B. Transplanted dopaminergic neurons develop PD pathologic changes: A second case report. Mov. Disord. 2008, 23, 2303–2306. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Englund, E.; Holton, J.L.; Soulet, D.; Hagell, P.; Lees, A.J.; Lashley, T.; Quinn, N.P.; Rehncrona, S.; Björklund, A.; et al. Lewy bodies in grafted neurons in subjects with Parkinson’s disease suggest host-to-graft disease propagation. Nat. Med. 2008, 14, 501–503. [Google Scholar] [CrossRef] [PubMed]
- Bae, E.J.; Lee, H.J.; Lee, S.J. Cell Models to Study Cell-to-Cell Transmission of α-Synuclein. Methods Mol. Biol. 2016, 1345, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Emmanouilidou, E.; Melachroinou, K.; Roumeliotis, T.; Garbis, S.D.; Ntzouni, M.; Margaritis, L.H.; Stefanis, L.; Vekrellis, K. Cell-produced alpha-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival. J. Neurosci. 2010, 30, 6838–6851. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Patel, S.; Lee, S.J. Intravesicular localization and exocytosis of alpha-synuclein and its aggregates. J. Neurosci. 2005, 25, 6016–6024. [Google Scholar] [CrossRef] [PubMed]
- Visanji, N.P.; Brooks, P.L.; Hazrati, L.N.; Lang, A.E. The prion hypothesis in Parkinson’s disease: Braak to the future. Acta Neuropathol. Commun. 2013, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Prymaczok, N.C.; Riek, R.; Gerez, J. More than a Rumor Spreads in Parkinson’s Disease. Front. Hum. Neurosci. 2016, 10, 608. [Google Scholar] [CrossRef] [PubMed]
- Jucker, M.; Walker, L.C. Propagation and spread of pathogenic protein assemblies in neurodegenerative diseases. Nat. Neurosci. 2018, 21, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shao, Q.; Yuan, Y.H.; Chen, N.H. Prion-like propagation of α-synuclein in the gut-brain axis. Brain Res. Bull. 2018, 140, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.; Debs, L.H.; Patel, A.P.; Nguyen, D.; Patel, K.; O’Connor, G.; Grati, M.; Mittal, J.; Yan, D.; Eshraghi, A.A.; et al. Neurotransmitters: The Critical Modulators Regulating Gut-Brain Axis. J. Cell. Physiol. 2017, 232, 2359–2372. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kasper, L.H. The role of microbiome in central nervous system disorders. Brain Behav. Immun. 2014, 38, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felice, V.D.; Quigley, E.M.; Sullivan, A.M.; O’Keeffe, G.W.; O’Mahony, S.M. Microbiota-gut-brain signalling in Parkinson’s disease: Implications for non-motor symptoms. Parkinsonism Relat. Disord. 2016, 27, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Calvani, R.; Picca, A.; Lo Monaco, M.R.; Landi, F.; Bernabei, R.; Marzetti, E. Of Microbes and Minds: A Narrative Review on the Second Brain Aging. Front. Med. 2018, 5, 53. [Google Scholar] [CrossRef] [PubMed]
- Pan-Montojo, F.; Anichtchik, O.; Dening, Y.; Knels, L.; Pursche, S.; Jung, R.; Jackson, S.; Gille, G.; Spillantini, M.G.; Reichmann, H.; et al. Progression of Parkinson’s disease pathology is reproduced by intragastric administration of rotenone in mice. PLoS ONE 2010, 5, e8762. [Google Scholar] [CrossRef] [PubMed]
- Pan-Montojo, F.; Schwarz, M.; Winkler, C.; Arnhold, M.; O’Sullivan, G.A.; Pal, A.; Said, J.; Marsico, G.; Verbavatz, J.M.; Rodrigo-Angulo, M.; et al. Environmental toxins trigger PD-like progression via increased alpha-synuclein release from enteric neurons in mice. Sci. Rep. 2012, 2, 898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noorian, A.R.; Rha, J.; Annerino, D.M.; Bernhard, D.; Taylor, G.M.; Greene, J.G. α-Synuclein transgenic mice display age-related slowing of gastrointestinal motility associated with transgene expression in the vagal system. Neurobiol. Dis. 2012, 48, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Svensson, E.; Horváth-Puhó, E.; Thomsen, R.W.; Djurhuus, J.C.; Pedersen, L.; Borghammer, P.; Sørensen, H.T. Vagotomy and subsequent risk of Parkinson’s disease. Ann. Neurol. 2015, 78, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Unger, M.M.; Spiegel, J.; Dillmann, K.U.; Grundmann, D.; Philippeit, H.; Bürmann, J.; Faßbender, K.; Schwiertz, A.; Schäfer, K.H. Short chain fatty acids and gut microbiota differ between patients with Parkinson’s disease and age-matched controls. Parkinsonism Relat. Disord. 2016, 32, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased intestinal permeability correlates with sigmoid mucosa α-synuclein staining and endotoxin exposure markers in early Parkinson’s disease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef] [PubMed]
- Keshavarzian, A.; Green, S.J.; Engen, P.A.; Voigt, R.M.; Naqib, A.; Forsyth, C.B.; Mutlu, E.; Shannon, K.M. Colonic bacterial composition in Parkinson’s disease. Mov. Disord. 2015, 30, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Caputi, V.; Giron, M.C. Microbiome-Gut-Brain Axis and Toll-Like Receptors in Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 1689. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, S.; Goto, S.; Tsuji, H.; Okuno, T.; Asahara, T.; Nomoto, K.; Shibata, A.; Fujisawa, Y.; Minato, T.; Okamoto, A.; et al. Intestinal Dysbiosis and Lowered Serum Lipopolysaccharide-Binding Protein in Parkinson’s Disease. PLoS ONE 2015, 10, e0142164. [Google Scholar] [CrossRef] [PubMed]
- Scheperjans, F.; Aho, V.; Pereira, P.A.; Koskinen, K.; Paulin, L.; Pekkonen, E.; Haapaniemi, E.; Kaakkola, S.; Eerola-Rautio, J.; Pohja, M.; et al. Gut microbiota are related to Parkinson’s disease and clinical phenotype. Mov. Disord. 2015, 30, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.A.; Saltykova, I.V.; Zhukova, I.A.; Alifirova, V.M.; Zhukova, N.G.; Dorofeeva, Y.B.; Tyakht, A.V.; Kovarsky, B.A.; Alekseev, D.G.; Kostryukova, E.S.; et al. Analysis of Gut Microbiota in Patients with Parkinson’s Disease. Bull. Exp. Biol. Med. 2017, 162, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Bedarf, J.R.; Hildebrand, F.; Coelho, L.P.; Sunagawa, S.; Bahram, M.; Goeser, F.; Bork, P.; Wüllner, U. Functional implications of microbial and viral gut metagenome changes in early stage L-DOPA-naïve Parkinson’s disease patients. Genome Med. 2017, 9, 39. [Google Scholar] [CrossRef] [PubMed]
- Hill-Burns, E.M.; Debelius, J.W.; Morton, J.T.; Wissemann, W.T.; Lewis, M.R.; Wallen, Z.D.; Peddada, S.D.; Factor, S.A.; Molho, E.; Zabetian, C.P.; et al. Parkinson’s disease and Parkinson’s disease medications have distinct signatures of the gut microbiome. Mov. Disord. 2017, 32, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.; Brown, S.M.; Hao, Y.; Tetz, V. Parkinson’s disease and bacteriophages as its overlooked contributors. Sci. Rep. 2018, 8, 10812. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, M.; Bonazzi, P.; Scarpellini, E.; Bendia, E.; Lauritano, E.C.; Fasano, A.; Ceravolo, M.G.; Capecci, M.; Rita Bentivoglio, A.; Provinciali, L.; et al. Prevalence of small intestinal bacterial overgrowth in Parkinson’s disease. Mov. Disord. 2011, 26, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Bove, F.; Gabrielli, M.; Petracca, M.; Zocco, M.A.; Ragazzoni, E.; Barbaro, F.; Piano, C.; Fortuna, S.; Tortora, A.; et al. The role of small intestinal bacterial overgrowth in Parkinson’s disease. Mov. Disord. 2013, 28, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.H.; Mahadeva, S.; Thalha, A.M.; Gibson, P.R.; Kiew, C.K.; Yeat, C.M.; Ng, S.W.; Ang, S.P.; Chow, S.K.; Tan, C.T.; et al. Small intestinal bacterial overgrowth in Parkinson’s disease. Parkinsonism Relat. Disord. 2014, 20, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Cassani, E.; Barichella, M.; Cancello, R.; Cavanna, F.; Iorio, L.; Cereda, E.; Bolliri, C.; Zampella Maria, P.; Bianchi, F.; Cestaro, B.; et al. Increased urinary indoxyl sulfate (indican): New insights into gut dysbiosis in Parkinson’s disease. Parkinsonism Relat. Disord. 2015, 21, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, S.; Mohajeri, M.H. Changes of Colonic Bacterial Composition in Parkinson’s Disease and Other Neurodegenerative Diseases. Nutrients 2018, 10, 708. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.J.; Chiu, I.M. Bacterial Signaling to the Nervous System through Toxins and Metabolites. J. Mol. Biol. 2017, 429, 587–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Clairembault, T.; Leclair-Visonneau, L.; Coron, E.; Bourreille, A.; Le Dily, S.; Vavasseur, F.; Heymann, M.F.; Neunlist, M.; Derkinderen, P. Structural alterations of the intestinal epithelial barrier in Parkinson’s disease. Acta Neuropathol. Commun. 2015, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Lionnet, A.; Leclair-Visonneau, L.; Neunlist, M.; Murayama, S.; Takao, M.; Adler, C.H.; Derkinderen, P.; Beach, T.G. Does Parkinson’s disease start in the gut? Acta Neuropathol. 2018, 135, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pituch, A.; Walkowiak, J.; Banaszkiewicz, A. Butyric acid in functional constipation. Prz. Gastroenterol. 2013, 8, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Ohira, H.; Tsutsui, W.; Fujioka, Y. Are Short Chain Fatty Acids in Gut Microbiota Defensive Players for Inflammation and Atherosclerosis? J. Atheroscler. Thromb. 2017, 24, 660–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engen, P.A.; Dodiya, H.B.; Naqib, A.; Forsyth, C.B.; Green, S.J.; Voigt, R.M.; Kordower, J.H.; Mutlu, E.A.; Shannon, K.M.; Keshavarzian, A. The Potential Role of Gut-Derived Inflammation in Multiple System Atrophy. J. Parkinsons Dis. 2017, 7, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.M.; Kamel, F.; Ross, G.W.; Jewell, S.A.; Marras, C.; Hoppin, J.A.; Umbach, D.M.; Bhudhikanok, G.S.; Meng, C.; Korell, M.; et al. Peptidoglycan recognition protein genes and risk of Parkinson’s disease. Mov. Disord. 2014, 29, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.; Liu, L.F.; Meng, N.; Tang, Z.; Chua, K.K.; Chen, L.L.; Song, J.X.; Mok, V.C.; Xie, L.X.; Li, M.; et al. LC-MS-based urinary metabolite signatures in idiopathic Parkinson’s disease. J. Proteome Res. 2015, 14, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.; Berk, M.; Carvalho, A.; Caso, J.R.; Sanz, Y.; Walder, K.; Maes, M. The Role of the Microbial Metabolites Including Tryptophan Catabolites and Short Chain Fatty Acids in the Pathophysiology of Immune-Inflammatory and Neuroimmune Disease. Mol. Neurobiol. 2017, 54, 4432–4451. [Google Scholar] [CrossRef] [PubMed]
- Stolzenberg, E.; Berry, D.; Yang, D.; Lee, E.Y.; Kroemer, A.; Kaufman, S.; Wong, G.C.L.; Oppenheim, J.J.; Sen, S.; Fishbein, T.; et al. A Role for Neuronal Alpha-Synuclein in Gastrointestinal Immunity. J. Innate Immun. 2017, 9, 456–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braak, H.; de Vos, R.A.; Bohl, J.; del Tredici, K. Gastric alpha-synuclein immunoreactive inclusions in Meissner’s and Auerbach’s plexuses in cases staged for Parkinson’s disease-related brain pathology. Neurosci. Lett. 2006, 396, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Shannon, K.M.; Keshavarzian, A.; Dodiya, H.B.; Jakate, S.; Kordower, J.H. Is alpha-synuclein in the colon a biomarker for premotor Parkinson’s disease? Evidence from 3 cases. Mov. Disord. 2012, 27, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Shannon, K.M.; Keshavarzian, A.; Mutlu, E.; Dodiya, H.B.; Daian, D.; Jaglin, J.A.; Kordower, J.H. α-Synuclein in colonic submucosa in early untreated Parkinson’s disease. Mov. Disord. 2012, 27, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Gold, A.; Turkalp, Z.T.; Munoz, D.G. Enteric alpha-synuclein expression is increased in Parkinson’s disease but not Alzheimer’s disease. Mov. Disord. 2013, 28, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Hilton, D.; Stephens, M.; Kirk, L.; Edwards, P.; Potter, R.; Zajicek, J.; Broughton, E.; Hagan, H.; Carroll, C. Accumulation of α-synuclein in the bowel of patients in the pre-clinical phase of Parkinson’s disease. Acta Neuropathol. 2014, 127, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Visanji, N.P.; Marras, C.; Kern, D.S.; Al Dakheel, A.; Gao, A.; Liu, L.W.; Lang, A.E.; Hazrati, L.N. Colonic mucosal a-synuclein lacks specificity as a biomarker for Parkinson disease. Neurology 2015, 84, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Lebouvier, T.; Neunlist, M.; Bruley des Varannes, S.; Coron, E.; Drouard, A.; N’Guyen, J.M.; Chaumette, T.; Tasselli, M.; Paillusson, S.; Flamand, M.; et al. Colonic biopsies to assess the neuropathology of Parkinson’s disease and its relationship with symptoms. PLoS ONE 2010, 5, e12728. [Google Scholar] [CrossRef] [PubMed]
- Pouclet, H.; Lebouvier, T.; Coron, E.; des Varannes, S.B.; Rouaud, T.; Roy, M.; Neunlist, M.; Derkinderen, P. A comparison between rectal and colonic biopsies to detect Lewy pathology in Parkinson’s disease. Neurobiol. Dis. 2012, 45, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Corbillé, A.G.; Letournel, F.; Kordower, J.H.; Lee, J.; Shanes, E.; Neunlist, M.; Munoz, D.G.; Derkinderen, P.; Beach, T.G. Evaluation of alpha-synuclein immunohistochemical methods for the detection of Lewy-type synucleinopathy in gastrointestinal biopsies. Acta Neuropathol. Commun. 2016, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebouvier, T.; Chaumette, T.; Damier, P.; Coron, E.; Touchefeu, Y.; Vrignaud, S.; Naveilhan, P.; Galmiche, J.P.; Bruley des Varannes, S.; Derkinderen, P.; et al. Pathological lesions in colonic biopsies during Parkinson’s disease. Gut 2008, 57, 1741–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beach, T.G.; Adler, C.H.; Sue, L.I.; Vedders, L.; Lue, L.; White Iii, C.L.; Akiyama, H.; Caviness, J.N.; Shill, H.A.; Sabbagh, M.N.; et al. Multi-organ distribution of phosphorylated alpha-synuclein histopathology in subjects with Lewy body disorders. Acta Neuropathol. 2010, 119, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Gelpi, E.; Navarro-Otano, J.; Tolosa, E.; Gaig, C.; Compta, Y.; Rey, M.J.; Martí, M.J.; Hernández, I.; Valldeoriola, F.; Reñé, R.; et al. Multiple organ involvement by alpha-synuclein pathology in Lewy body disorders. Mov. Disord. 2014, 29, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Stokholm, M.G.; Danielsen, E.H.; Hamilton-Dutoit, S.J.; Borghammer, P. Pathological α-synuclein in gastrointestinal tissues from prodromal Parkinson disease patients. Ann. Neurol. 2016, 79, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ferro, Á.; Rábano, A.; Catalán, M.J.; Rodríguez-Valcárcel, F.C.; Fernández Díez, S.; Herreros-Rodríguez, J.; García-Cobos, E.; Álvarez-Santullano, M.M.; López-Manzanares, L.; Mosqueira, A.J.; et al. In vivo gastric detection of α-synuclein inclusions in Parkinson’s disease. Mov. Disord. 2015, 30, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Bloch, A.; Probst, A.; Bissig, H.; Adams, H.; Tolnay, M. α-synuclein pathology of the spinal and peripheral autonomic nervous system in neurologically unimpaired elderly subjects. Neuropathol. Appl. Neurobiol. 2006, 32, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, K.; Takahashi, H.; Ohama, E.; Ikuta, F. Parkinson’s disease: An immunohistochemical study of Lewy body-containing neurons in the enteric nervous system. Acta Neuropathol. 1990, 79, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, D.A.; Bieger, D.; deVente, J.; Steinbusch, W.M. Vagal efferent projections: Viscerotopy, neurochemistry and effects of vagotomy. Prog. Brain Res. 1996, 107, 79–96. [Google Scholar] [PubMed]
- Halliday, G.M.; Li, Y.W.; Blumbergs, P.C.; Joh, T.H.; Cotton, R.G.; Howe, P.R.; Blessing, W.W.; Geffen, L.B. Neuropathology of immunohistochemically identified brainstem neurons in Parkinson’s disease. Ann. Neurol. 1990, 27, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, K.; Toyoshima, Y.; Awamori, K.; Anezaki, T.; Yoshimoto, M.; Tsuji, S.; Takahashi, H. Restricted occurrence of Lewy bodies in the dorsal vagal nucleus in a patient with late-onset parkinsonism. J. Neurol. Sci. 1999, 165, 188–191. [Google Scholar] [CrossRef]
- Del Tredici, K.; Rüb, U.; de Vos, R.A.; Bohl, J.R.; Braak, H. Where does parkinson disease pathology begin in the brain? J. Neuropathol. Exp. Neurol. 2002, 61, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Sastre, M.; Bohl, J.R.; de Vos, R.A.; del Tredici, K. Parkinson’s disease: Lesions in dorsal horn layer I, involvement of parasympathetic and sympathetic pre- and postganglionic neurons. Acta Neuropathol. 2007, 113, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Böttner, M.; Zorenkov, D.; Hellwig, I.; Barrenschee, M.; Harde, J.; Fricke, T.; Deuschl, G.; Egberts, J.H.; Becker, T.; Fritscher-Ravens, A.; et al. Expression pattern and localization of alpha-synuclein in the human enteric nervous system. Neurobiol. Dis. 2012, 48, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Adler, C.H.; Dugger, B.N.; Hinni, M.L.; Lott, D.G.; Driver-Dunckley, E.; Hidalgo, J.; Henry-Watson, J.; Serrano, G.; Sue, L.I.; Nagel, T.; et al. Submandibular gland needle biopsy for the diagnosis of Parkinson disease. Neurology 2014, 82, 858–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beach, T.G.; Adler, C.H.; Dugger, B.N.; Serrano, G.; Hidalgo, J.; Henry-Watson, J.; Shill, H.A.; Sue, L.I.; Sabbagh, M.N.; Akiyama, H.; et al. Submandibular gland biopsy for the diagnosis of Parkinson disease. J. Neuropathol. Exp. Neurol. 2013, 72, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, P.S.; Esiri, M.M.; Reading, M.; McShane, R.; Nagy, Z. α-Synuclein pathology in the olfactory pathways of dementia patients. J. Anat. 2007, 211, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Beach, T.G.; White, C.L.; Hladik, C.L.; Sabbagh, M.N.; Connor, D.J.; Shill, H.A.; Sue, L.I.; Sasse, J.; Bachalakuri, J.; Henry-Watson, J.; et al. Olfactory bulb alpha-synucleinopathy has high specificity and sensitivity for Lewy body disorders. Acta Neuropathol. 2009, 117, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L. Olfaction in Parkinson’s disease and related disorders. Neurobiol. Dis. 2012, 46, 527–552. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; del Tredici, K. Neuropathological Staging of Brain Pathology in Sporadic Parkinson’s disease: Separating the Wheat from the Chaff. J. Parkinsons Dis. 2017, 7, S71–S85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braak, H.; Ghebremedhin, E.; Rüb, U.; Bratzke, H.; del Tredici, K. Stages in the development of Parkinson’s disease-related pathology. Cell Tissue Res. 2004, 318, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Del Tredici, K.; Braak, H. Spinal cord lesions in sporadic Parkinson’s disease. Acta Neuropathol. 2012, 124, 643–664. [Google Scholar] [CrossRef] [PubMed]
- Borghammer, P. How does Parkinson’s disease begin? Perspectives on neuroanatomical pathways, prions, and histology. Mov. Disord. 2018, 33, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.H.; del Tredici, K.; Braak, H. Parkinson’s disease: A dual-hit hypothesis. Neuropathol. Appl. Neurobiol. 2007, 33, 599–614. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.T.; Munoz, D.G.; Gray, D.A.; Schlossmacher, M.G.; Woulfe, J.M. Alpha-synuclein in the appendiceal mucosa of neurologically intact subjects. Mov. Disord. 2014, 29, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Fang, F.; Pedersen, N.L.; Tillander, A.; Ludvigsson, J.F.; Ekbom, A.; Svenningsson, P.; Chen, H.; Wirdefeldt, K. Vagotomy and Parkinson disease: A Swedish register-based matched-cohort study. Neurology 2017, 88, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Kurowska, Z.; Englund, E.; Widner, H.; Lindvall, O.; Li, J.Y.; Brundin, P. Signs of degeneration in 12-22-year old grafts of mesencephalic dopamine neurons in patients with Parkinson’s disease. J. Parkinsons Dis. 2011, 1, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Peelaerts, W.; Bousset, L.; Baekelandt, V.; Melki, R. α-Synuclein strains and seeding in Parkinson’s disease, incidental Lewy body disease, dementia with Lewy bodies and multiple system atrophy: Similarities and differences. Cell Tissue Res. 2018, 373, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.M. Environmental toxins and Parkinson’s disease. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 141–164. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.J.; Walter, G.C.; Wilder, S.L.; Baronowsky, E.A.; Powley, T.L. α-Synuclein-immunopositive myenteric neurons and vagal preganglionic terminals: Autonomic pathway implicated in Parkinson’s disease? Neuroscience 2008, 153, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Ulusoy, A.; Phillips, R.J.; Helwig, M.; Klinkenberg, M.; Powley, T.L.; di Monte, D.A. Brain-to-stomach transfer of α-synuclein via vagal preganglionic projections. Acta Neuropathol. 2017, 133, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.M.; Li, Z.; Jiao, Y.; Gaborit, N.; Pani, A.K.; Orrison, B.M.; Bruneau, B.G.; Giasson, B.I.; Smeyne, R.J.; Gershon, M.D.; et al. Extensive enteric nervous system abnormalities in mice transgenic for artificial chromosomes containing Parkinson disease-associated alpha-synuclein gene mutations precede central nervous system changes. Hum. Mol. Genet. 2010, 19, 1633–1650. [Google Scholar] [CrossRef] [PubMed]
- Ubeda-Bañon, I.; Saiz-Sanchez, D.; de la Rosa-Prieto, C.; Mohedano-Moriano, A.; Fradejas, N.; Calvo, S.; Argandoña-Palacios, L.; Garcia-Muñozguren, S.; Martinez-Marcos, A. Staging of α-synuclein in the olfactory bulb in a model of Parkinson’s disease: Cell types involved. Mov. Disord. 2010, 25, 1701–1707. [Google Scholar] [CrossRef] [PubMed]
- Liddle, R.A. Parkinson’s disease from the gut. Brain Res. 2018, 1693, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Antonioli, L.; Colucci, R.; Blandizzi, C.; Fornai, M. Interplay among gut microbiota, intestinal mucosal barrier and enteric neuro-immune system: A common path to neurodegenerative diseases? Acta Neuropathol. 2018, 136, 345–361. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Qian, Y.; Xu, S.; Song, Y.; Xiao, Q. Longitudinal Analysis of Fecal Microbiome and Pathologic Processes in a Rotenone Induced Mice Model of Parkinson’s Disease. Front. Aging Neurosci. 2017, 9, 441. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.G.; Stribinskis, V.; Rane, M.J.; Demuth, D.R.; Gozal, E.; Roberts, A.M.; Jagadapillai, R.; Liu, R.; Choe, K.; Shivakumar, B.; et al. Exposure to the Functional Bacterial Amyloid Protein Curli Enhances α-Synuclein Aggregation in Aged Fischer 344 Rats and Caenorhabditis elegans. Sci. Rep. 2016, 6, 34477. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.P.; Carvey, P.M.; Keshavarzian, A.; Shannon, K.M.; Shaikh, M.; Bakay, R.A.; Kordower, J.H. Progression of intestinal permeability changes and alpha-synuclein expression in a mouse model of Parkinson’s disease. Mov. Disord. 2014, 29, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Arentsen, T.; Khalid, R.; Qian, Y.; Diaz Heijtz, R. Sex-dependent alterations in motor and anxiety-like behavior of aged bacterial peptidoglycan sensing molecule 2 knockout mice. Brain Behav. Immun. 2018, 67, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M.M. Microbiota-Brain-Gut Axis and Neurodegenerative Diseases. Curr. Neurol. Neurosci. Rep. 2017, 17, 94. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Yun, M.; Oh, Y.J.; Choi, H.J. Mind-altering with the gut: Modulation of the gut-brain axis with probiotics. J. Microbiol. 2018, 56, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Breid, S.; Bernis, M.E.; Babila, J.T.; Garza, M.C.; Wille, H.; Tamgüney, G. Neuroinvasion of α-Synuclein Prionoids after Intraperitoneal and Intraglossal Inoculation. J. Virol. 2016, 90, 9182–9193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Suk, J.E.; Lee, K.W.; Park, S.H.; Blumbergs, P.C.; Gai, W.P.; Lee, S.J. Transmission of Synucleinopathies in the Enteric Nervous System of A53T α-Synuclein Transgenic Mice. Exp. Neurobiol. 2011, 20, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Holmqvist, S.; Chutna, O.; Bousset, L.; Aldrin-Kirk, P.; Li, W.; Björklund, T.; Wang, Z.Y.; Roybon, L.; Melki, R.; Li, J.Y. Direct evidence of Parkinson pathology spread from the gastrointestinal tract to the brain in rats. Acta Neuropathol. 2014, 128, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacino, A.N.; Brooks, M.; Thomas, M.A.; McKinney, A.B.; Lee, S.; Regenhardt, R.W.; McGarvey, N.H.; Ayers, J.I.; Notterpek, L.; Borchelt, D.R.; et al. Intramuscular injection of α-synuclein induces CNS α-synuclein pathology and a rapid-onset motor phenotype in transgenic mice. Proc. Natl. Acad. Sci. USA 2014, 111, 10732–10737. [Google Scholar] [CrossRef] [PubMed]
- Rey, N.L.; Steiner, J.A.; Maroof, N.; Luk, K.C.; Madaj, Z.; Trojanowski, J.Q.; Lee, V.M.; Brundin, P. Widespread transneuronal propagation of α-synucleinopathy triggered in olfactory bulb mimics prodromal Parkinson’s disease. J. Exp. Med. 2016, 213, 1759–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koller, E.J.; Brooks, M.M.; Golde, T.E.; Giasson, B.I.; Chakrabarty, P. Inflammatory pre-conditioning restricts the seeded induction of α-synuclein pathology in wild type mice. Mol. Neurodegener. 2017, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Sacino, A.N.; Brooks, M.; McKinney, A.B.; Thomas, M.A.; Shaw, G.; Golde, T.E.; Giasson, B.I. Brain injection of α-synuclein induces multiple proteinopathies, gliosis, and a neuronal injury marker. J. Neurosci. 2014, 34, 12368–12378. [Google Scholar] [CrossRef] [PubMed]
- Masuda-Suzukake, M.; Nonaka, T.; Hosokawa, M.; Oikawa, T.; Arai, T.; Akiyama, H.; Mann, D.M.; Hasegawa, M. Prion-like spreading of pathological α-synuclein in brain. Brain 2013, 136, 1128–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peelaerts, W.; Bousset, L.; van der Perren, A.; Moskalyuk, A.; Pulizzi, R.; Giugliano, M.; van den Haute, C.; Melki, R.; Baekelandt, V. α-Synuclein strains cause distinct synucleinopathies after local and systemic administration. Nature 2015, 522, 340–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmotilib, H.; Maltbie, T.; Delic, V.; Liu, Z.; Hu, X.; Fraser, K.B.; Moehle, M.S.; Stoyka, L.; Anabtawi, N.; Krendelchtchikova, V.; et al. α-Synuclein fibril-induced inclusion spread in rats and mice correlates with dopaminergic Neurodegeneration. Neurobiol. Dis. 2017, 105, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Ou, M.T.; Karuppagounder, S.S.; Kam, T.I.; Yin, X.; Xiong, Y.; Ge, P.; Umanah, G.E.; Brahmachari, S.; Shin, J.H.; et al. Pathological α-synuclein transmission initiated by binding lymphocyte-activation gene 3. Science 2016, 353, aah3374. [Google Scholar] [CrossRef] [PubMed]
- Luk, K.C.; Kehm, V.M.; Zhang, B.; O’Brien, P.; Trojanowski, J.Q.; Lee, V.M. Intracerebral inoculation of pathological α-synuclein initiates a rapidly progressive neurodegenerative α-synucleinopathy in mice. J. Exp. Med. 2012, 209, 975–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sargent, D.; Verchère, J.; Lazizzera, C.; Gaillard, D.; Lakhdar, L.; Streichenberger, N.; Morignat, E.; Bétemps, D.; Baron, T. ‘Prion-like’ propagation of the synucleinopathy of M83 transgenic mice depends on the mouse genotype and type of inoculum. J. Neurochem. 2017, 143, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.C.; Giles, K.; Oehler, A.; Middleton, L.; Dexter, D.T.; Gentleman, S.M.; DeArmond, S.J.; Prusiner, S.B. Transmission of multiple system atrophy prions to transgenic mice. Proc. Natl. Acad. Sci. USA 2013, 110, 19555–19560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mougenot, A.L.; Nicot, S.; Bencsik, A.; Morignat, E.; Verchère, J.; Lakhdar, L.; Legastelois, S.; Baron, T. Prion-like acceleration of a synucleinopathy in a transgenic mouse model. Neurobiol. Aging 2012, 33, 2225–2228. [Google Scholar] [CrossRef] [PubMed]
- Recasens, A.; Dehay, B.; Bové, J.; Carballo-Carbajal, I.; Dovero, S.; Pérez-Villalba, A.; Fernagut, P.O.; Blesa, J.; Parent, A.; Perier, C.; et al. Lewy body extracts from Parkinson disease brains trigger α-synuclein pathology and neurodegeneration in mice and monkeys. Ann. Neurol. 2014, 75, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulusoy, A.; Rusconi, R.; Pérez-Revuelta, B.I.; Musgrove, R.E.; Helwig, M.; Winzen-Reichert, B.; di Monte, D.A. Caudo-rostral brain spreading of α-synuclein through vagal connections. EMBO Mol. Med. 2013, 5, 1119–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angot, E.; Steiner, J.A.; Lema Tomé, C.M.; Ekström, P.; Mattsson, B.; Björklund, A.; Brundin, P. α-synuclein cell-to-cell transfer and seeding in grafted dopaminergic neurons in vivo. PLoS ONE 2012, 7, e39465. [Google Scholar] [CrossRef] [PubMed]
- Desplats, P.; Lee, H.J.; Bae, E.J.; Patrick, C.; Rockenstein, E.; Crews, L.; Spencer, B.; Masliah, E.; Lee, S.J. Inclusion formation and neuronal cell death through neuron-to-neuron transmission of α-synuclein. Proc. Natl. Acad. Sci. USA 2009, 106, 13010–13015. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Hiniker, A.; Kuo, Y.M.; Nussbaum, R.L.; Liddle, R.A. α-Synuclein in gut endocrine cells and its implications for Parkinson’s disease. JCI Insight 2017, 2, 92295. [Google Scholar] [CrossRef] [PubMed]
- Kalaitzakis, M.E.; Graeber, M.B.; Gentleman, S.M.; Pearce, R.K. The dorsal motor nucleus of the vagus is not an obligatory trigger site of Parkinson’s disease: A critical analysis of α-synuclein staging. Neuropathol. Appl. Neurobiol. 2008, 34, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Attems, J.; Jellinger, K.A. The dorsal motor nucleus of the vagus is not an obligatory trigger site of Parkinson’s disease. Neuropathol. Appl. Neurobiol. 2008, 34, 466–467. [Google Scholar] [CrossRef] [PubMed]
- Fumimura, Y.; Ikemura, M.; Saito, Y.; Sengoku, R.; Kanemaru, K.; Sawabe, M.; Arai, T.; Ito, G.; Iwatsubo, T.; Fukayama, M.; et al. Analysis of the adrenal gland is useful for evaluating pathology of the peripheral autonomic nervous system in Lewy body disease. J. Neuropathol. Exp. Neurol. 2007, 66, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Tysnes, O.B.; Kenborg, L.; Herlofson, K.; Steding-Jessen, M.; Horn, A.; Olsen, J.H.; Reichmann, H. Does vagotomy reduce the risk of Parkinson’s disease? Ann. Neurol. 2015, 78, 1011–1012. [Google Scholar] [CrossRef] [PubMed]
- Recasens, A.; Carballo-Carbajal, I.; Parent, A.; Bové, J.; Gelpi, E.; Tolosa, E.; Vila, M. Lack of pathogenic potential of peripheral α-synuclein aggregates from Parkinson’s disease patients. Acta Neuropathol. Commun. 2018, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rietdijk, C.D.; Perez-Pardo, P.; Garssen, J.; van Wezel, R.J.; Kraneveld, A.D. Exploring Braak’s Hypothesis of Parkinson’s Disease. Front. Neurol. 2017, 8, 37. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
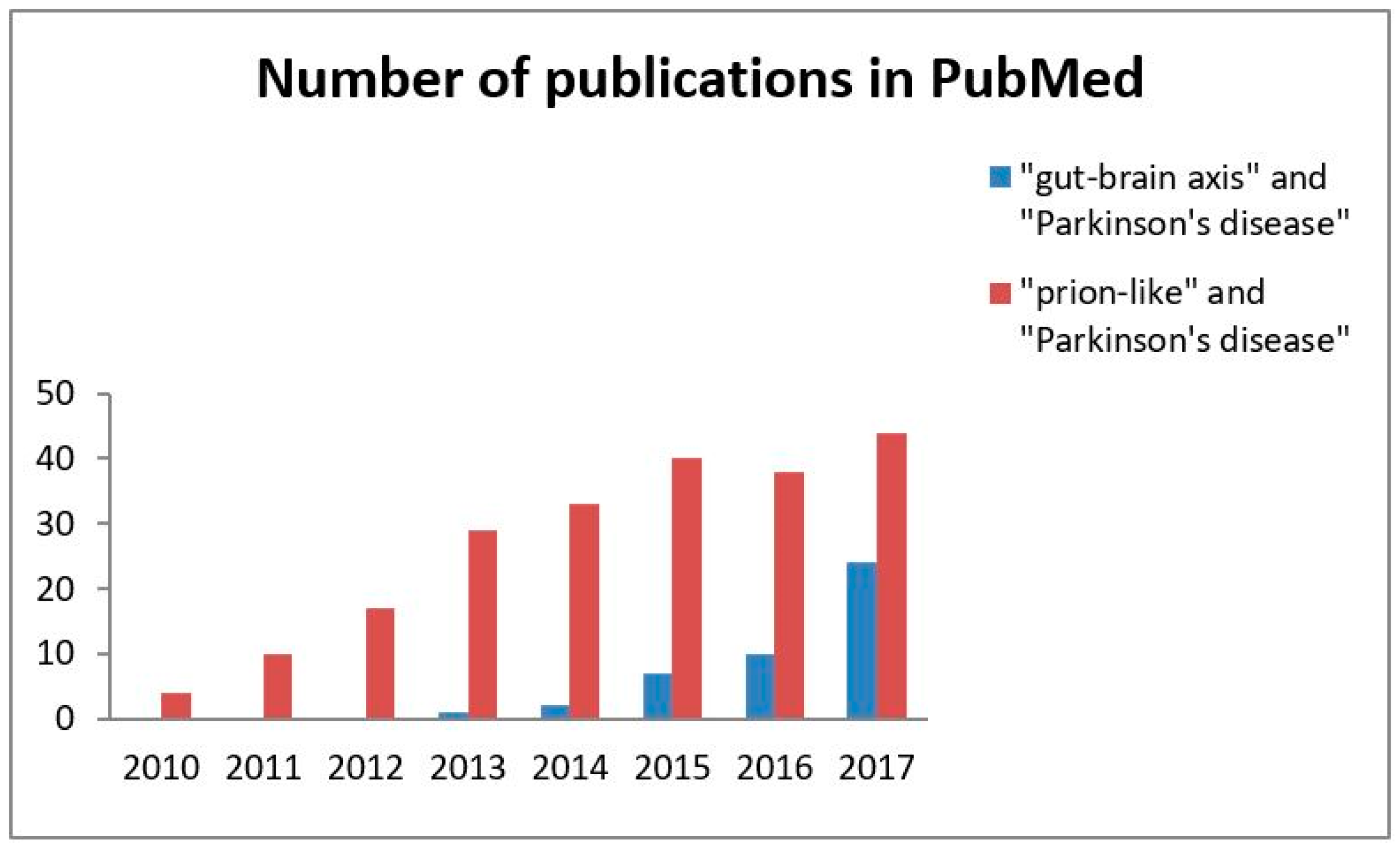{kind=link}
| Subjects/Specimens | Outcome/Results | Reference |
|---|---|---|
| Gut Dysbiosis/SIBO | ||
| Intestinal biopsy samples from PD subjects and HC | ↑ intestinal permeability (↑ urinary sucralose excretion, ↓ plasma LBP level) ↑ E. coli in lamina propria and epithelium ↑ inflammation/oxidative stress (↑ 3-NT) ↑ α-syn immunoreactivity in PD patients | Forsyth et al. 2011 [30] |
| Faecal samples from PD patients and cohabitants | ↓ total fecal bacterial ↓ Clostridium coccoides (C. leptum) and Bacteroides fragilis ↑ Lactobacillus L. gasseri counts correlated positively with disease durations and negatively with stool frequencies C. coccoides counts correlated negatively with disease durations and positively with stool frequencies ↑ intestinal permeability (↓ serum LBP level) in PD patients | Hasegawa et al. 2015 [33] |
| Sigmoid mucosal biopsy and faecal samples from PD subjects and HC | ↑ putative proinflammatory bacteria from the family Oxalobacteraceae (Proteobacteria, class Betaproteobacteria) and genus Ralstonia (Proteobacteria, class Betaproteobacteria, family Oxalobacteraceae) in the mucosa ↓ putative “anti-inflammatory” butyrate-producing bacteria from the genera Blautia, Coprococcus, and Roseburia in faces ↓ microbiome genes involved in normal metabolic activities of microbiota ↑ microbiome genes involved in bacterial secretion, LPS, and ubiquinone biosynthesis in PD patients | Keshavarzian et al. 2015 [31] |
| Fecal samples from PD patients and HC | ↓ Prevotellaceae ↑ Enterobacteriaceae positively correlated with the severity of postural instability and gait difficulty ↑ Verrucomicrobiaceae and Bradyrhizobiaceae associated with the degree of constipation in PD patients | Scheperjans et al. 2015 [34] |
| Fecal samples from PD patients and HC | ↓ Dorea, Bacteroides, Prevotella, Faecalibacterium, Bacteroides massiliensis, Stoquefichus massiliensis, Bacteroides coprocola, Blautia glucerasea, Dorea longicatena, Bacteroides dorei, Bacteroides plebeus, Prevotella copri, Coprococcus eutactus, and Ruminococcus callidus ↑ Christensenella, Catabacter, Lactobacillus, Oscillospira, Bifidobacterium, Christensenella minuta, Catabacter hongkongensis, Lactobacillus mucosae, Ruminococcus bromii, and Papillibacter cinnamivorans in PD patients | Petrov et al. 2016 [35] |
| Fecal samples from PD patients and HC | ↓ phyla Bacteroidetes and Prevotellaceae ↓ Lactobacillaceae and Enterococcaceae ↓ Faecalibacterium prausnitzii ↑ Bifidobacterium and Enterobacteriaceae ↓ SCFAs (acetate, propionate and butyrate) in PD patients | Unger et al. 2016 [29] |
| Stool samples from PD patients and HC | ↑ Verrucomicrobiaceae (genus Akkermansia, including Akkermansia muciniphila and Alistipes shahii.) ↑ Firmicutes ↓ Prevotellaceae (genus Prevotella, including Prevotella copri) ↓ Erysipelotrichaceae (genus Eubacterium, including Eubacterium biforme) ↓ Clostridium saccharolyticum ↓ microbiome glucuronate pathway (d-Glucuronate degradation, d-glucuronate → pyruvate, and d-glyceraldehyde) ↑ microbiome tryptophan pathway (tryptophan→kynurenine→2-aminomuconate) in PD patients | Bedarf et al. 2017 [36] |
| Stool samples of PD patients and HC | ↑ Bifidobacteriaceae, Christensenellaceae, Tissierellaceae, Lactobacillaceae, and Verrucomicrobiaceae families ↓ Lachnospiraceae, Pasteurellaceae 26 microbiome’ metabolic pathways changed in PD patients | Hill-Burns et al. 2017 [37] |
| Faecal samples from drug-naive PD patients HCs. | ↓ Prevotellaceae, Lachnospiraceae, Lactobacillaceae and Streptococcaceae ↓ phages belonged to the Siphoviridae family (Bacillus, Enterobacteria, Lactococcus Streptococcus, and Salmonella phages) and Lactobacillus phages of the Myoviridae family ↑ phages belonged to the Siphoviridae family (Leuconostoc, Lactococcus, and Enterobacteria phages) Enterobacteria phages of the Myoviridae family, and Salmonella phages of the Podoviridae family ↑ phage/bacteria ratio for Lactococcus in PD patients | Tetz et al. 2018 [38] |
| breath samples from PD patients and HC | SIBO (↑ hydrogen level in GBT) ↑ bloating and flatulence ↑ motor complications (UPDRS-III) in PD patients | Gabrielli et al. 2011 [39] |
| Breath samples from PD patients and HC | SIBO (↑ hydrogen and methane levels in GBT and LBT) ↑ motor complications (UPDRS-IV) in PD patients | Fasano et al. 2013 [40] |
| Breath samples from PD patients and HC | SIBO (↑ hydrogen in LBT) in early stage PD | Tan et al. 2014 [41] |
| Urinary samples from PD patients and HCs | SIBO (↑ urine indican concentrations) in PD patients | Cassani et al. 2015 [42] |
| Peripheral distribution of α-synuclein pathology | ||
| Autopsy GI tissue samples from PD subjects | LBs in the myenteric plexus of the lower oesophagus | Wakabayashi et al. 1988 [7] |
| Postmortem study of the brain from PD subjects | LBs in TH-positive neurons in the midbrain, pons, medulla (DMV), rostral medial lemniscus, raphe obscurus nucleus, dorsal tegmental bundle | Halliday et al. 1990 [72] |
| Autopsy GI specimens from PD subjects | LBs in the paravertebral and celiac sympathetic ganglia and in the Auerbach’s plexus of the lower upper, middle and lower esophagus, duodenum, ileum, descending colon and rectum, and Meissner’s plexuses of the duodenum and descending colon | Wakabayashi et al. 1990 [70] |
| Postmortem study of the brain from PD subjects | LBs in DMV | Wakabayashi et al. 1999 [73] |
| Postmortem study of the brain from PD subjects | LNs and LBs in non-catecholaminergic neurons of the dorsal glossopharyngeus-vagus complex, projection neurons of the intermediate reticular zone, olfactory bulb, olfactory tract, and/or anterior olfactory nucleus | Del Tredici et al. 2002 [74] |
| Autopsy specimens of brains, spinal cords and PANS from PD subjects | LNs in the intermediolateral nucleus of the thoracic spinal cord, sacral parasympathetic cell column of the sacral spinal cord, dorsal and anterior horns, paravertebral chain ganglia, vagus nerve and DMV | Bloch et al. 2006 [69] |
| Autopsy stomach specimens from PD subjects | α-syn aggregations, including LNs and LBs in the gastric wall (peripheral nerve, fibers generated from the Auerbach plexus, nerve fiber bundle of Meissner’s plexus), gastric Meissner plexus and Auerbach plexus | Braak et al. 2006 [55] |
| Postmortem study of the brain and complete spinal cords from PD subjects | α-syn aggregations, including LBs and LNs, in spinal cord lamina I neurons, parasympathetic preganglionic projection neurons of the vagal nerve, sympathetic preganglionic neurons of the spinal cord, postganglionic neurons of the coeliac ganglion | Braak et al. 2007 [75] |
| Autopsy brain tissues from PD subjects | LBs and LNs in the present in olfactory bulb and tract, anterior olfactory nucleus, orbitofrontal cortex, amygdala and hippocampus | Hubbard et al. 2007 [79] |
| Colonic biopsies from PD patients | p-α-syn immunoreactivity in the submucosal neurites | Lebouvier et al. 2008 [64] |
| Olfactory bulb sections from PD patients | synucleinopathy in the olfactory bulb olfactory bulb synucleinopathy density scores correlated significantly with: those in other brain regions (anterior medulla, anterior pons, midbrain, amygdala, gyrus, inferior parietal lobule) mini mental state examinations and UPDRS scale (motor part) score | Beach et al. 2009 [80] |
| A whole-body autopsy of PD subjects | p-α-syn immunoreactivity in the spinal cord and vagus nerve as well as in the submandibular gland, in the submucosa of the lower esophagus, in the stroma of the pancreas, in the submucosa of a primary bronchus, in the submucosa of the larynx, in the adrenal medulla, in the stroma of the parathyroid gland, and in the ovary | Beach et al. 2010 [65] |
| Colonic biopsies from PD patients | p-α-syn immunoreactivity in the submucosal neurites and submucosal plexus LP positively correlated with dysarthria and postural instability, constipation severity | Lebouvier et al. 2010 [61] |
| Gastrointestinal specimens from patients with no history of neurological and psychiatric diseases | α-syn mRNA expression in full-thickness sections and intestinal wall layers (submucosa and tunica muscularis) α-syn mRNA expression in myenteric ganglia. α-syn immunoreactivity in ganglia of the myenteric and submucosal plexus as well as in intramuscular nerve fibers p-α-syn immunoreactivity in ganglia of the myenteric and submucosal plexus | Böttner et al. 2012 [76] |
| Colonic biopsies from PD patients | p-α-syn immunoreactivity (including LNs) in the ascending colon, descending colon and rectum | Pouclet et al. 2012 [62] |
| Colonic biopsies from PD patients | α-syn within neuronal tissues in the colonic submucosa | Shannon et al. 2012 [56] |
| Colonic biopsies from PD patients | α-syn-positive structures in colonic submucosa 2 to 5 years before the first reported symptom of PD | Shannon et al. 2012 [57] |
| Submandibular gland autopsy in an elderly subject with PD | p-α-syn immunoreactivity in submandibular gland | Beach et al. 2013 [78] |
| Colonic specimens from PD patients | α-syn expression in myenteric and submucosal ganglia and extraganglionic nerve fibers | Gold et al. 2013 [58] |
| Submandibular gland biopsies from PD patients | p-α-syn immunoreactivity in nerve fibers or puncta within submandibular salivary gland tissue | Adler et al. 2014 [77] |
| Multiorgan (including GI tract) postmortem study of PD subjects | p-α-syn/α-syn immunoreactivity in the distal esophagus, stomach, ileum, colon, and rectum (ganglia of the myenteric plexus) | Gelpi et al. 2014 [66] |
| GI tissue samples from patients with no history of neurological disease | α-syn immunoreactivity in the mucosal plexus of the vermiform appendix α-syn immunoreactivity in the CD68-immunoreactive macrophages in the appendiceal lamina propria α-syn immunoreactivity in the mucosa of the stomach, ileum and colon | Gray et al. 2014 [87] |
| GI tissue samples from PD patients | p-α-syn immunoreactivity in the stomach, small intestine, large intestine, α-syn-positive neurites within the lamina propria and submucosal myenteric nerve fibres α-syn small rounded inclusions within submucosal ganglia α-syn-positive biopsies correlated with autonomic symptoms (sometimes prior to motor symptoms) | Hilton et al. 2014 [59] |
| Colonic biopsies from PD patients | p-α-syn immunoreactivity (LBs) in the submucosa ↓ occludin expression | Clairembault et al. 2015 [46] |
| Stomach biopsies from PD patients | α-syn immunoreactivity (antrum, pylorus, and duodenum) | Sánchez-Ferro et al. 2015 [68] |
| Colonic mucosal biopsies from PD patients | α-syn and p-α-syn immunoreactivity in individuals with early PD and with later PD > control subjects | Visanji et al. 2015 [60] |
| colonic biopsies from PD patients | LTS in the lamina propria of the mucosa, submucosa, and epithelium | Corbillé et al. 2016 [63] |
| PD Multiorgan (including GI tract) postmortem study of PD subjects | p-α-syn/α-syn immunoreactivity in the nasal region, oral region, salivary gland, esophagus, stomach, small intestine and appendix (mucosa, submucosal ganglia, intramuscular nerve fibers, and myenteric ganglia) | Stokholm et al. 2016 [67] |
| Vagotomy and risk of PD | ||
| A cohort of individuals underwent vagotomy and matched general population (control) | ↓ risk of PD in patients who underwent full truncal vagotomy | Svensson et al. 2015 [28] |
| Vagotomized patients (truncal, selective, and unknown); reference individuals | ↓ risk of PD in patients with truncal vagotomy vs. selective vagotomy ↓ risk of PD in patients with truncal vagotomy vs. reference individuals (ns) | Liu et al. 2017 [88] |
| Prion-like spread of α-synuclein | ||
| An individual with PD transplanted bilaterally with human ventral mesencephalon derived from embryos—a post-mortem examination | ↑ α-syn and ubiquitin immunoreactivity ↓ DAT immunoreactivity in grafted neurons 14 years after transplantation | Kordower et al. 2008 [11] |
| An individual with PD transplanted bilaterally with human ventral mesencephalon derived from embryos—a post-mortem examination | ↑ α-syn, ubiquitin and thioflavin S immunoreactivity ↓ DAT an TH immunoreactivity in grafted neurons 14 years after transplantation | Kordower et al. 2008 [12] |
| PD subjects transplanted fetal mesencephalic dopaminergic neurons—a post-mortem examination | ↑ α-syn immunoreactivity ↓ TH-positive neurons in grafted neurons 11-16 years after transplantation | Li et al. 2008 [13] |
| PD subjects transplanted fetal mesencephalic dopaminergic neurons—a post-mortem examination | ↑ α-syn immunoreactivity ↓ DAT immunoreactivity ↓ TH-positive neurons in grafted neurons 13–22 years after transplantation | Kurowska et al. 2011 [89] |
| Experimental Model | Outcome/Results | Reference |
|---|---|---|
| Dysbiosis | ||
| ASO mice under GF condition | ↓ p-α-syn and aggregation-specific α-syn immunoreactivity in the CP and SN ↓ α-syn insoluble expression ↑ numbers and total lengths of microglia branches within the CP and SN (↓ activation state) ↓ Iba-1 immunoreactivity in microglia in the CP ↓ TNF-α and IL-6 in the CP, Mid and FC ↑ motor function (beam traversal, pole descent, and hindlimb clasping reflexes) ↑ fecal pellets ↓ SCFAs production | Sampson et al. 2016 [99] |
| ASO mice under SPF condition | ↑ motor function (beam traversal, pole descent, and hindlimb clasping reflexes) ↓ Iba-1 immunoreactivity in microglia in the CP ↓ microglia diameter in the CP and SN (↓ activation state) ↑ fecal pellets ↓ SCFAs production | Sampson et al. 2016 [99] |
| ASO mice treated (p.o.) with microbiota from PD patients | ↓ fecal pellets ↓ motor function (beam traversal, pole descent and nasal adhesive removal) | Sampson et al. 2016 [99] |
| Male C57BL/6 mice treated with rotenone (i.g.) | ↓ Bacteroidetes ↑ Firmicutes and Firmicutes/Bacteroidetes ratio ↑ 11 metabolic pathways ↑ α-syn and p-α-syn immunoreactivity in the colon ↑ α-syn and p-α-syn immunoreactivity in the midbrain ↓ colon motility and stool water content ↓ locomotor deficits in the open field and pole tests ↑ TNF-α, IL-6 and iNOS expression ↑ TLR2 expression | Yang et al. 2017 [98] |
| Peripheral distribution of α-synuclein | ||
| Subdiaphragmatic vagotomy in aged Fischer 344 and SD rats | ↓ α-syn-positive varicosities within ganglia and α-syn-positive axons in fiber bundles | Phillips et al. 2008 [92] |
| Tg A53T and A30P mice | α-syn immunoreactivity within myenteric and submucosal plexuses ↓ WGTT and colonic motility ↓ motor behavior (open field test, rotarod test) | Kuo et al. 2010 [94] |
| C57BL/6J mice treated with rotenone (i.g.) | ↑ α-syn phosphorylation, accumulation and aggregation with gliosis in ENS ganglia ↑ intracellular and axonal α-syn in the spinal cord and the brainstem (IML and DMV); the α-syn pathology progressed into the SN | Pan-Montojo et al. 2010 [25] |
| TgA53T mice | α-syn immunoreactivity in the olfactory bulb (glomerular, mitral, and granule layers) high α-syn immunoreactivity in the calcium binding protein-positive cells low α-syn immunoreactivity in the Nissl-positive cells | Ubeda-Bañon et al. 2010 [95] |
| TGA53T mice injected with human brain tissue extract containing LBs into the stomach wall | ↑ α-syn deposition in the myenteric neurons ↑ activation of macrophages (MHC-II expression) | Lee et al. 2011 [106] |
| Subdiaphragmatic vagotomy in TgA53T mice | ↓ α-syn immunoreactivity in the myenteric plexus of the dorsal and ventral stomach and duodenum | Noorian et al. 2012 [27] |
| Hemivagotomy or partial sympathectomy of the nervus mesentericus inferior in mice orally treated with rotenone | ↓ α-syn expression in the lumbal IML of SRT mice ↓ α-syn expression in the ipsilateral DMV of HRT mice ↑ dopaminergic cell death in the ipsilateral SNc of HRT mice ↓ motor deficits (rotarod test) | Pan-Montojo et al. 2012 [26] |
| SD rats injected with brain lysates from PD patient into the intestine wall of stomach and duodenum | α-syn immunoreactivity in the intestinal wall and in the vagal nerve as well as in ChAT-positive neurons in the DMV (microtubule associated transport) | Holmqvist et al. 2014 [107] |
| C57 /BL6 mice treated with LPS | ↑ α-syn expression in the large intestine p-α-syn in colonic myenteric neurons ↑ intestinal permeability | Kelly et al. 2014 [101] |
| Aged Fischer rats fed with E. coli producing amyloid protein curli | ↑ α-syn deposition in gut ganglion cells (myenteric plexus and submucosa) and in neurons in hippocampus and striatum ↑ Iba-1 and GFAP immunoreactivity in the striatum, hippocampus and neocortex | Chen et al. 2016 [100] |
| Prion-like spread of α-synuclein | ||
| Thy-1 α-syn TG mice injected with MCNSCs into the hippocampus | hα-syn immunoreactivity in MCNSCs caspase 3 immunoreactivity in MCNSCs | Desplats et al. 2009 [123] |
| SD rats with engineered nigral neurons to express hα-syn by AAV2/6 intrastriatally transplanted with embryonic VM neurons | hα-syn immunoreactivity in grafted neurons EEA1-positive transmitted hα-syn PK-sensitive and p-α-syn-negative transmitted hα-syn | Angot et al. 2012 [122] |
| TgM83 mice inoculated intracerebrally with symptomatic aged M83 brain lysates harbouring aggregated α-syn or PFFs | p-α-syn immunoreactivity in the CNS thioflavin-S- and Ub-positive inclusions in the CNS ↓ TH-positive neurons ↑ GFAP and Iba-1 immunoreactivity in the CNS ↓ survival period | Luk et al. 2012 [116] |
| Young TgM83 mice inoculated intracerebrally with homogenates of the brain of old TgM83 mice | p-α-syn expression in the brain homogenate p-α-syn immunoreactivity in diffuse perikaryal inclusions and dystrophic neurites paralysis ↓ survival period | Mougenot et al. 2012 [119] |
| C57BL/6 J mice injected into substantia nigra with recombinant α-synuclein monomer, fibrils or insoluble fraction of brain tissue from DLB patients | p-α-syn immunoreactivity as well as Ub- and p62-positive inclusions in the brain including substantia nigra, amygdala, dentate gyrus, hippocampus, fimbria, stria terminalis, hypothalamus, somatosensory area, visual cortex, cingulate cortex and corpus callosum ↓ enkephalin | Masuda-Suzukake et al. 2013 [112] |
| C57BL/6 mice intracerebrally inoculated with human LB fraction Rhesus monkeys intrastriatally inoculated with human LB fraction | Mice: p-α-syn immunoreactivity in the striatum and neocortical areas ↓ SNpc TH-positive and Nissl-positive neurons ↑ Iba1-positive cells in the SNpc ↑ impaired motor ability (pole test) Monkey p-α-syn immunoreactivity in the in the putamen, SNpc, globus pallidus, precentral gyrus, superior frontal gyrus, and the entorhinal area in the temporal cortex ↓ SNpc TH-positive neurons ↓ striatal dopaminergic innervation and dopaminergic terminal | Recasens et al. 2014 [120] |
| SD rats injected with hα-synuclein-AAVs into the vagus nerve, controls | hα-syn immunoreactivity in DMV and nucleus ambiguous, MO, pons and midbrain ↑ rat and total (rat +human) α-syn mRNA vs. controls in MO | Ulusoy et al. 2013 [121] |
| TgM83 mice injected intracerebrally with brain homogenate from MSA patients | p-α-syn in the striatum, motor cortex, and thalamus ↑ GFAP and Iba-1 immunoreactivity in the brainstem neurologic dysfunction (ataxia, dysmetria, bradykinesia, and circling behaviour) ↓ survival periods | Watts et al. 2013 [118] |
| M20 and M83Tg mice intracerebrally injected with recombinant amyloidogenic or soluble α-syn | p-α-syn immunoreactivity in the hippocampus, cortex, striatum, midbrain, and brainstem, amygdala, thalamus, and hypothalamus NFL and p62 -reactive inclusions ↑ GFAP and Iba-1 immunoreactivity in the brain | Sacino et al. 2014 [111] |
| TgM83 TgM20 mice injected intramuscularly with a fibrillar mouse or human α-syn | hind limb paralysis p-α-syn immunoreactivity in the spinal cord, brainstem, midbrain, and cortex (astrocytes and neurons) p62 immunoreactivity in the brain and spinal cord ↑ GFAP and Iba-1 immunoreactivity in the CNS ↓ motor function ↓ survival period | Sacino et al. 2014 [108] |
| Wistar rats injected with α-syn (oligomers, fibrils or ribbons) into SN or striatum and rAAV-driven α-syn overexpression | p-α-syn immunoreactivity in the whole brain p62-positive inclusions ↓ TH-positive neurons, dopaminergic cells, projecting axons ↓ spontaneous bursts of action potentials in synapses ↑ motor deficit (↓ forelimb contacts in the cylinder test) | Peelaerts et al. 2015 [113] |
| TgM83 mice injected intraperitoneally or intraglossally with fibrils of human or mouse α-synuclein | p-α-syn immunoreactivity in the brain and spinal cord Sarkosyl-insoluble and Ub- and p62-positive inclusions in the brain and spinal cord ↑ GFAP and Iba-1 immunoreactivity in the brain paralysis, kyphosis, ↓ activity ↓ body weight | Breid et al. 2016 [105] |
| WT and TgLag3 mice striatally injected with α-syn PFFs | ↑ p-α-syn immunoreactivity in the SNpc TH–positive neurons ↓ TH– and Nissl-positive neurons in the SNpc ↓ DA, DOPAC, HVA ↓ TH and DAT ↓ motor function (pole and hindlimb clasping tests) | Mao et al. 2016 [115] |
| C57BL/6J mice injected with mPFFs or HuPFFs into OB | p-α-syn immunoreactivity within olfactory and brain regions (above 40 brain regions and subregions) ↑ p62-, Ub-, and thioflavin S-positive inclusions ↑ olfactory dysfunction | Rey et al. 2016 [109] |
| C57BL/6 J and C3H/HeJ mice and SD rats intracranially injected with α-syn fibrils or monomer | p-α-syn immunoreactivity in the SNpc, striatum, amygdala, cortex, and thalamus ↓ TH-positive and Nissl-positive neurons in the SNpc | Abdelmotilib et al. 2017 [114] |
| Tg A53T;CCK-GFP l mice | α-syn immunoreactivity in EECs of duodenum and colon α-syn immunoreactivity in submucosal enteric nerves TH-positive EECs α-syn in regions of GFAP-positive glial processes | Chandra et al. 2017 [124] |
| B6/C3H mice injected intraventricularly with rAAV-EV and intrahippocampally with α-syn fibrils | p-α-syn immunoreactivity in the whole brain argyrophilic and p62 immunoreactive inclusions ↑ GFAP and Iba-1 immunoreactivity in the forebrain | Koller et al. 2017 [110] |
| TgM83 mice injected intraperitoneally or intracerebrally with brains homogenates of sick M83 mice or MSA patients | p-α-syn immunoreactivity in the cerebellum, cerebral cortex, striatum, brainstem, mesencephalon, and in the spinal cord hind limb paralysis, balance disorder ↓ survival periods p-α-syn expression in the brainstem | Sargent et al. 2017 [117] |
| SD rats injected with hα-synuclein-AAVs into the vagus nerve or the midbrain | hα-syn immunoreactivity in DMV, nodose ganglion, efferent DMV projections and afferent vagal fibers, MO, NTS and gastric wall after vagal injection hα-syn-immunoreactivity in the striatum, hypothalamus, locus coeruleus, MO, DMV neurons, and gastric wall after midbrain injection | Ulusoy et al. 2017 [93] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kujawska, M.; Jodynis-Liebert, J. What Is the Evidence that Parkinson’s Disease Is a Prion Disorder, Which Originates in the Gut? Int. J. Mol. Sci. 2018, 19, 3573. https://doi.org/10.3390/ijms19113573
Kujawska M, Jodynis-Liebert J. What Is the Evidence that Parkinson’s Disease Is a Prion Disorder, Which Originates in the Gut? International Journal of Molecular Sciences. 2018; 19(11):3573. https://doi.org/10.3390/ijms19113573
Chicago/Turabian StyleKujawska, Małgorzata, and Jadwiga Jodynis-Liebert. 2018. "What Is the Evidence that Parkinson’s Disease Is a Prion Disorder, Which Originates in the Gut?" International Journal of Molecular Sciences 19, no. 11: 3573. https://doi.org/10.3390/ijms19113573
APA StyleKujawska, M., & Jodynis-Liebert, J. (2018). What Is the Evidence that Parkinson’s Disease Is a Prion Disorder, Which Originates in the Gut? International Journal of Molecular Sciences, 19(11), 3573. https://doi.org/10.3390/ijms19113573