A Thioredoxin Domain-Containing Protein Interacts with Pepino mosaic virus Triple Gene Block Protein 1


 ,
,  and
and 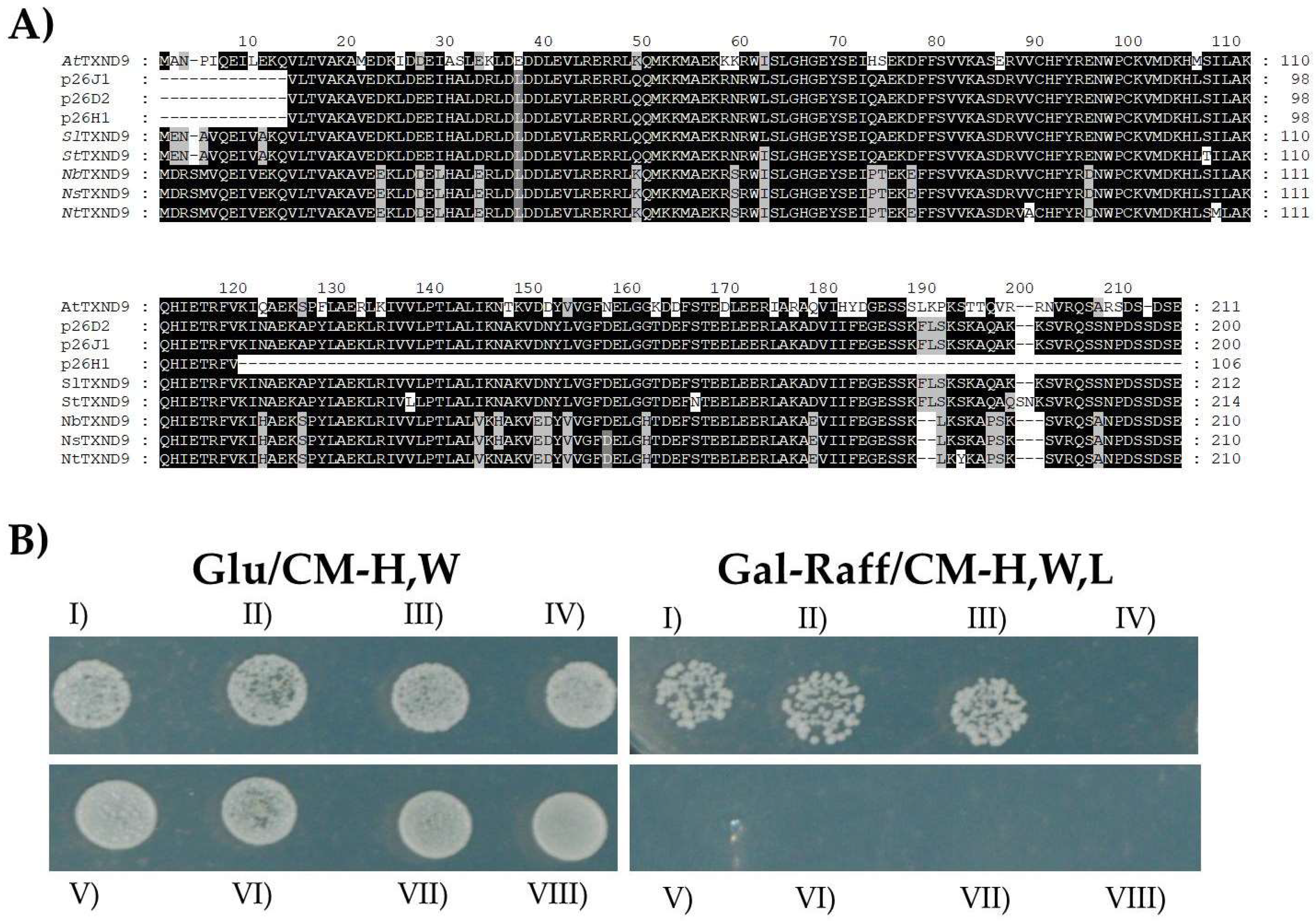{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. PepMV TGBp1 Interacts with an Uncharacterized Tomato cDNA in a Yeast Library Screen Assay
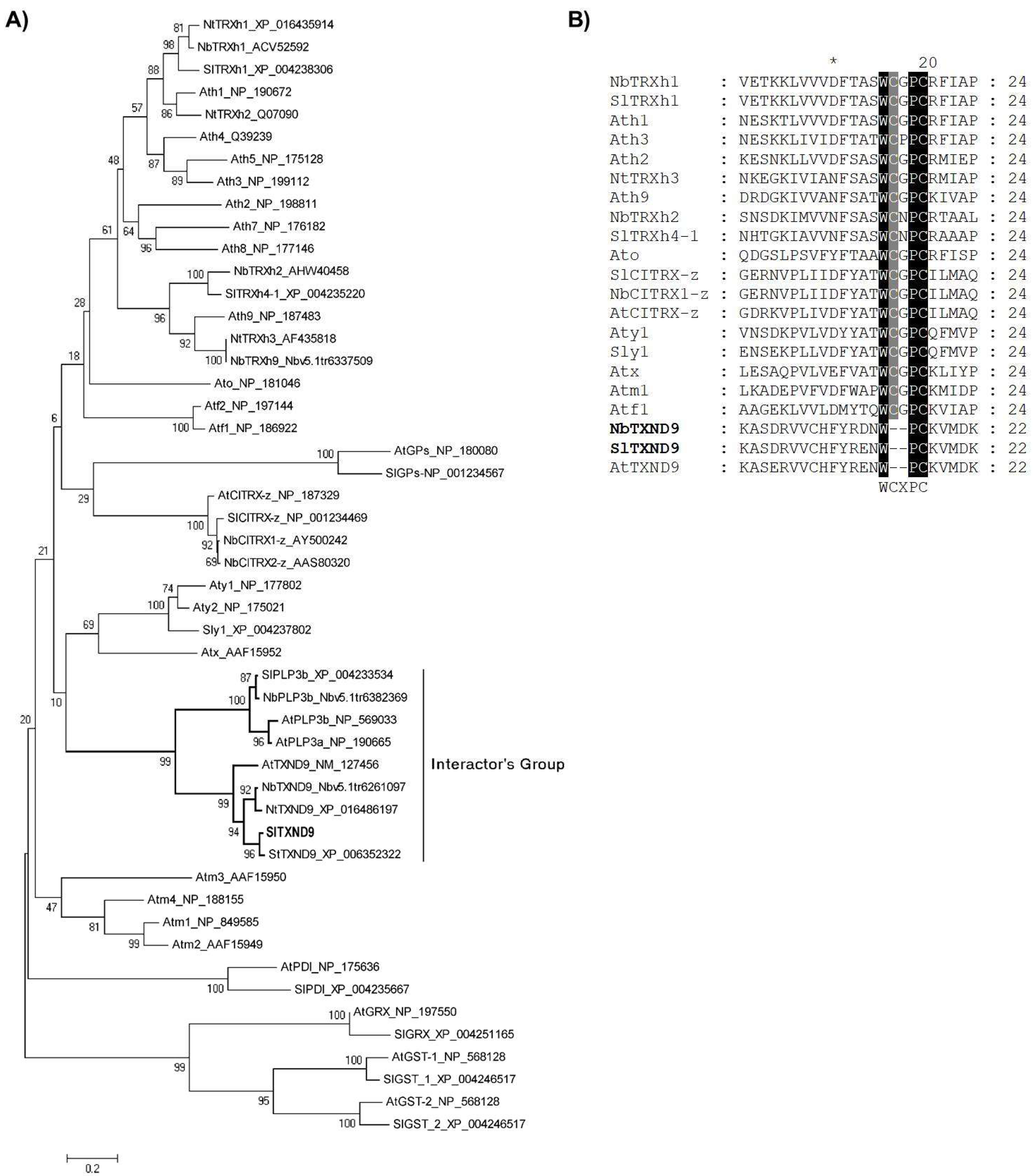
2.2. Tomato SlTXND9 Clusters Phylogenetically with a Phosducin-Like Protein 3
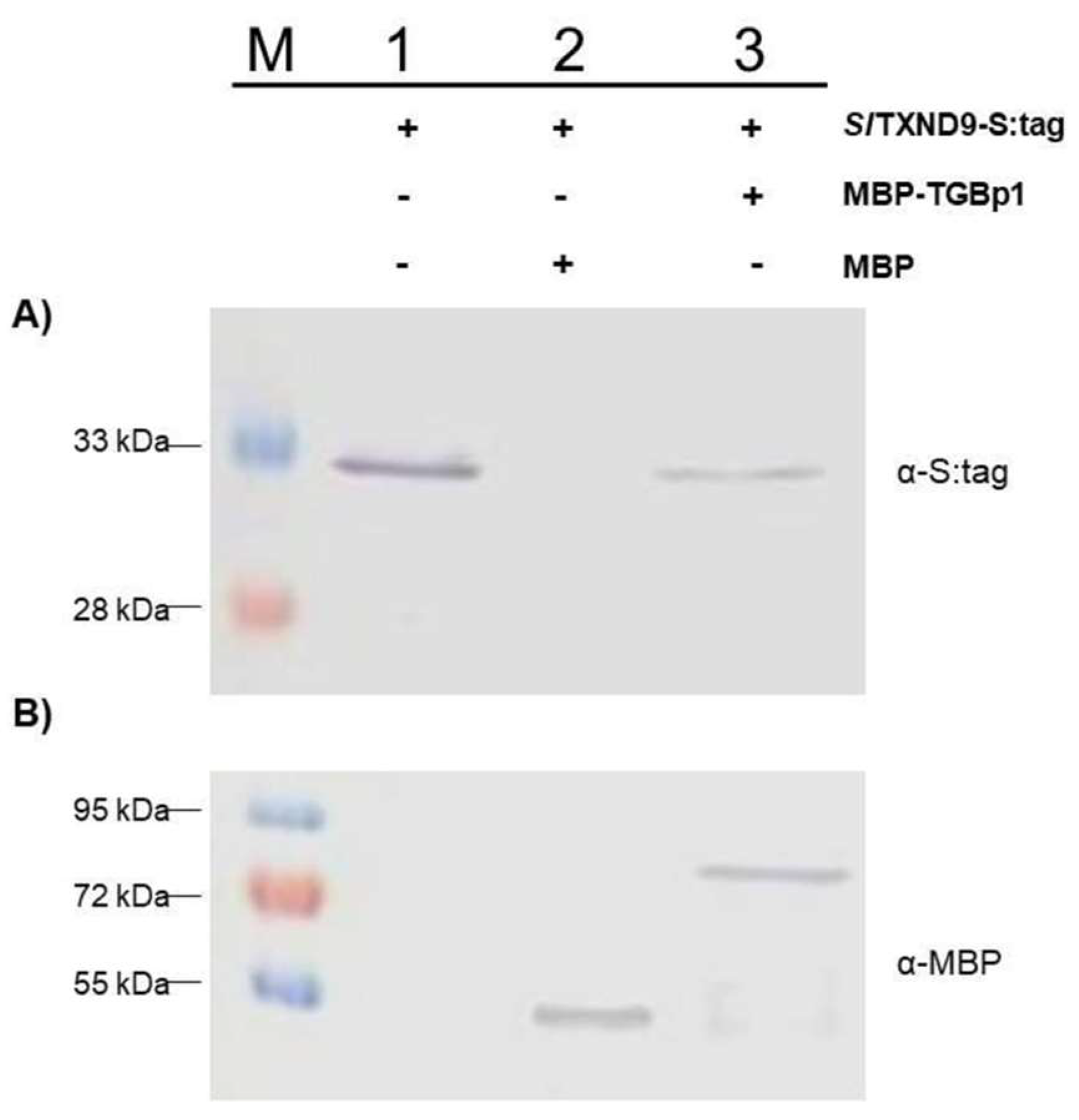
2.3. In Vitro Confirmation of the TGBp1-SlTXND9 Interaction
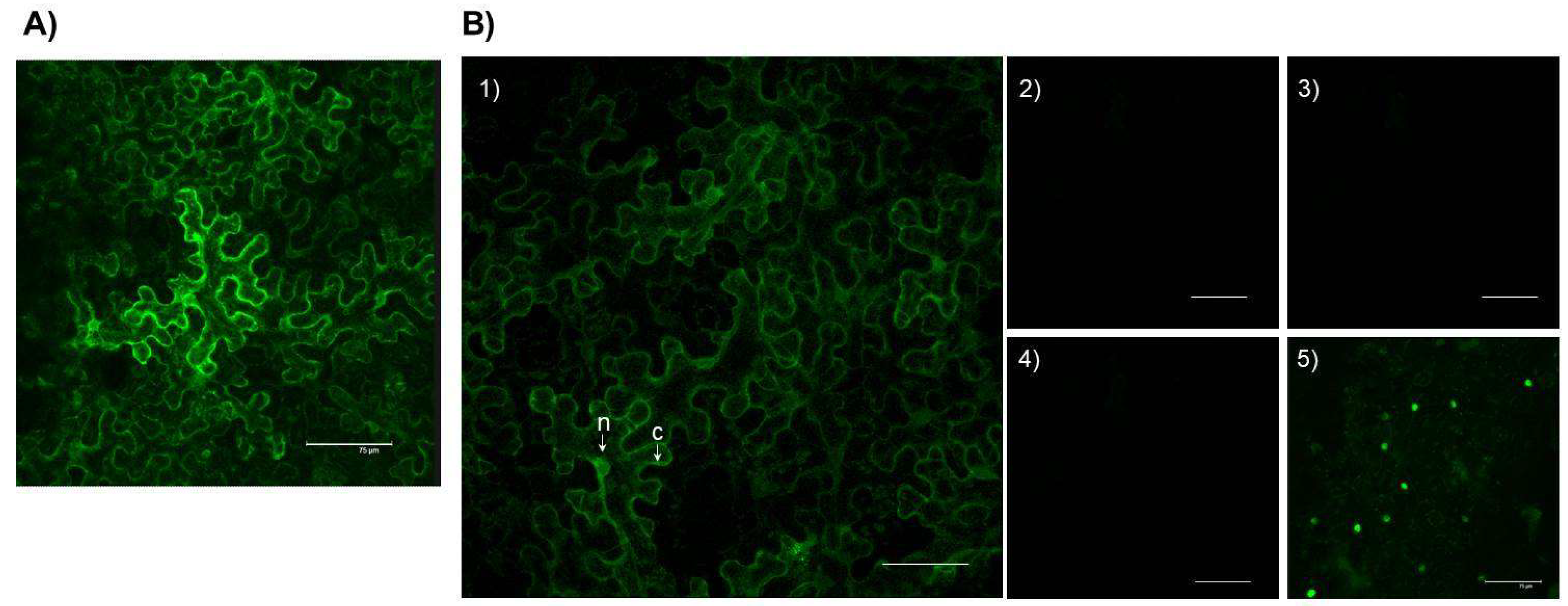
2.4. In Planta Localization of the SlTXND9-GFP and TGBp1:SlTXND9 Interaction
2.5. Measurement of NbTXND9 mRNA Levels on PepMV Infection
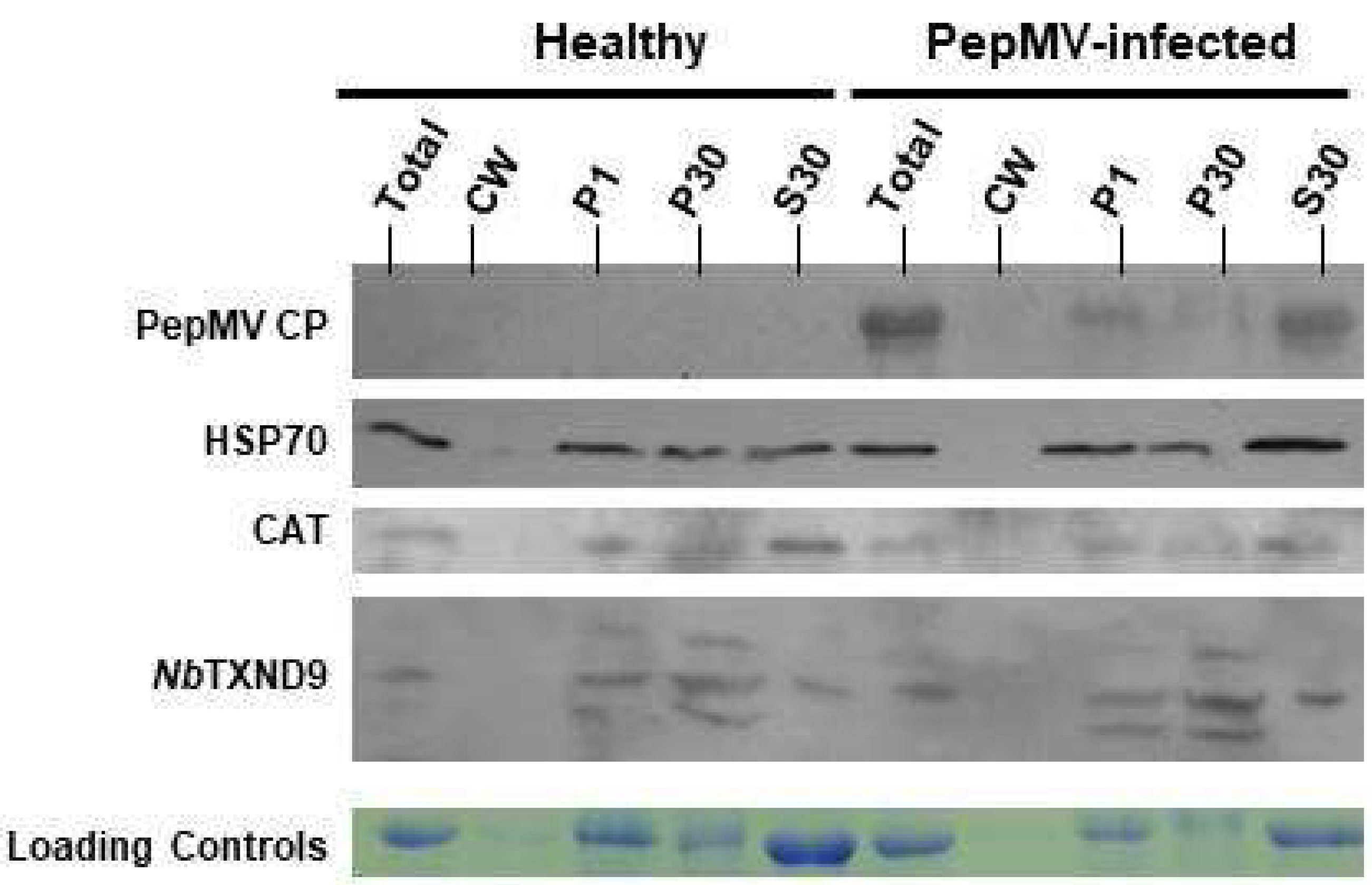
2.6. NbTXND9 Presence in Fractionated N. benthamiana Leaf Proteins
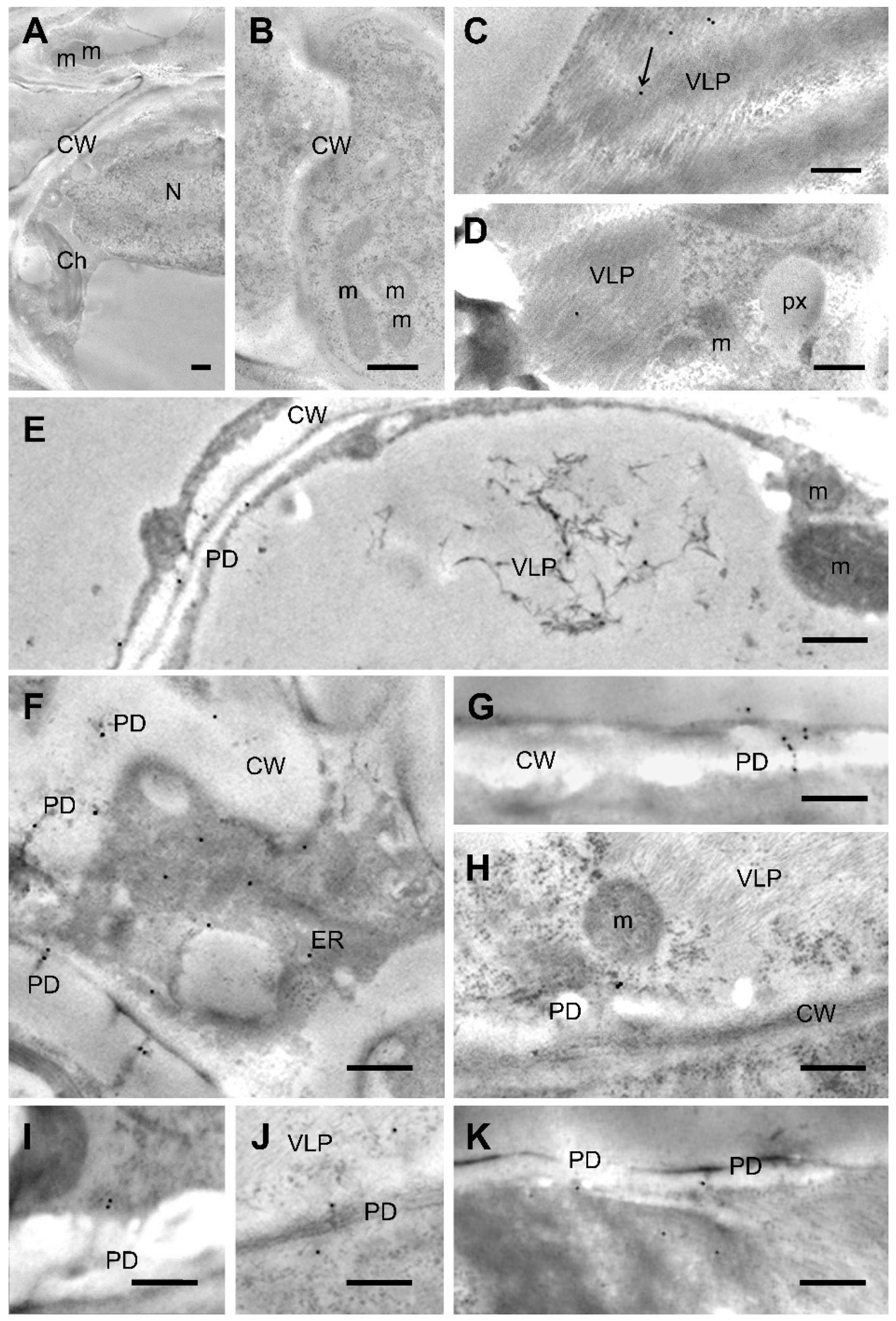
2.7. NbTXND9 Localizes in the Plasmodesmata and ER of PepMV-Infected Leaf Cells
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Conditions and Plant Inoculations
4.2. Tomato cDNA Library Screening in Yeast
4.3. Sequence and Phylogenetic Analysis
4.4. In Vitro Protein Binding—Pull Down Assay
4.5. BiFC Assay
4.6. Plant Response to PepMV Inoculation
4.7. Antiserum Production and Processing
4.8. Subcellular Fractionation
4.9. Immuno-Gold Labelling and Electron Microscopy
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanssen, I.M.; Thomma, B.P.H.J. Pepino mosaic virus: A successful pathogen that rapidly evolved from emerging to endemic in tomato crops. Mol. Plant Pathol. 2010, 11, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, I.M.; Lapidot, M.; Thomma, B.P.H.J. Emerging viral diseases of tomato crops. Mol. Plant-Microbe Interact. 2010, 23, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Hasiów-Jaroszewska, B.; Paeleman, A.; Ortega-Parra, N.; Borodynko, N.; Minicka, J.; Czerwoniec, A.; Thomma, B.P.H.J.; Hanssen, I.M. Ratio of mutated versus wild-type coat protein sequences in Pepino mosaic virus determines the nature and severity of yellowing symptoms on tomato plants. Mol. Plant Pathol. 2013, 14, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, J.M.; Hernandez-Gallardo, M.D.; Cenis, J.L.; Lacasa, A.; Aranda, M.A. Complete sequence of the Pepino mosaic virus RNA genome. Arch. Virol. 2002, 147, 2009–2015. [Google Scholar] [CrossRef] [PubMed]
- Osman, T.A.; Olsthoorn, R.C.; Livieratos, I.C. Role of the Pepino mosaic virus 3’-untranslated region elements in negative-strand RNA synthesis in vitro. Virus Res. 2014, 190, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Sempere, R.N.; Gómez, P.; Truniger, V.; Aranda, M.A. Development of expression vectors based on Pepino mosaic virus. Plant Methods 2011, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Hasiów-Jaroszewska, B.; Borodynko, N.; Pospieszny, H. Infectious RNA transcripts derived from cloned cDNA of a Pepino mosaic virus isolate. Arch. Virol. 2009, 154, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Mekuria, T.; Barnunusinghe, D.; Payton, M.; Verchot-Lubicz, J. Phloem unloading of Potato virus X movement proteins is regulated by virus and host factors. Mol. Plant-Microbe Interact. 2008, 21, 1106–1117. [Google Scholar] [CrossRef] [PubMed]
- Okano, Y.; Senshu, H.; Hashimoto, M.; Neriya, Y.; Netsu, O.; Minato, N.; Yoshida, T.; Maejima, K.; Oshima, K.; Komatsu, K.; et al. In planta recognition of a double-stranded RNA sunthesis protein complex by a potexviral RNA silencing suppressor. Plant Cell 2014, 26, 2168–2183. [Google Scholar] [CrossRef] [PubMed]
- Fridborg, I.; Grainger, J.; Page, A.; Coleman, M.; Findlay, K.; Angell, S. TIP, a novel host factor linking callose degradation with the cell-to-cell movement of Potato virus X. Mol. Plant-Microbe Interact. 2003, 16, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, I.M.; Van Esse, H.P.; Ballester, A.R.; Hogewoning, S.W.; Parra, N.O.; Paeleman, A.; Lievens, B.; Bovy, A.G.; Thomma, B.P.H.J. Differential tomato transcriptomic response induced by Pepino mosaic virus isolates with differential aggressiveness. Plant Physiol. 2011, 156, 301–318. [Google Scholar] [CrossRef] [PubMed]
- Sempere, R.N.; Gómez-Aix, C.; Ruíz-Ramón, F.; Gómez, P.; Hasiów-Jaroszewska, B.; Sánchez-Pina, M.A.; Aranda, M.A. Pepino mosaic virus RNA-dependent RNA polymerase pol domain is a hypersensitive response-like elicitor shared by necrotic and mild isolates. Phytopathology 2016, 106, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Hasiów-Jaroszewska, B.; Borodynko, N.; Jackowiak, P.; Figlerowicz, M.; Pospieszny, H. Single mutation converts mild pathotype of Pepino mosaic virus into necrotic one. Virus Res. 2011, 159, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Duff-Farrier, C.R.; Bailey, A.M.; Boonham, N.; Foster, G.D. A pathogenicity determinant maps to the N-terminal coat protein region of the Pepino mosaic virus genome. Mol. Plant Pathol. 2015, 16, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Candresse, T.; Marais, A.; Faure, C.; Dubrana, M.P.; Gombert, J.; Bendahmane, A. Multiple coat protein mutations abolish recognition of Pepino mosaic potexvirus (PepMV) by the potato Rx resistance gene in transgenic tomatoes. Mol. Plant-Microbe Interact. 2010, 23, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Mathioudakis, M.M.; Veiga, R.; Ghita, M.; Tsikou, D.; Medina, V.; Canto, T.; Makris, A.M.; Livieratos, I.C. Pepino mosaic virus capsid protein interacts with a tomato heat shock protein cognate 70. Virus Res. 2012, 163, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Mathioudakis, M.M.; Rodríguez-Moreno, L.; Sempere, R.N.; Aranda, M.A.; Livieratos, I. Multifaceted capsid proteins: Multiple interactions suggest multiple roles for Pepino mosaic virus capsid protein. Mol. Plant-Microbe Interact. 2014, 27, 1356–1369. [Google Scholar] [CrossRef] [PubMed]
- Mathioudakis, M.M.; Veiga, R.S.; Canto, T.; Medina, V.; Mossialos, D.; Makris, A.M.; Livieratos, I. Pepino mosaic virus triple gene block protein 1 (TGBp1) interacts with and increases tomato catalase 1 activity to enhance virus accumulation. Mol. Plant Pathol. 2013, 14, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.H.; Chen, H.T.; Huang, Y.P.; Huang, H.C.; Shenkwen, L.L.; Hsu, Y.H.; Tsai, C.H. A thioredoxin NbTRXh2 from Nicotiana benthamiana negatively regulates the movement of Bamboo mosaic virus. Mol. Plant Pathol. 2017, 19, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Chibani, K.; Wingsle, G.; Jacquot, J.P.; Gelhayde, E.; Rouhier, N. Comparative genomic study of the thioredoxin family in photysynthetic organisms with emphasis on Populus trichocarpa. Mol. Plant 2009, 2, 308–322. [Google Scholar] [CrossRef] [PubMed]
- Meyer, Y.; Siala, W.; Bashandy, T.; Riondet, C.; Vignols, F.; Reichheld, J.P. Glutaredoxins and thioredoxins in plants. BBA Mol. Cell Res. 2007, 1783, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Gelhaye, E.; Rouhier, N.; Navrot, N.; Jacquot, J.P. The plant thioredoxin system. Cell. Mol. Life Sci. 2005, 62, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Arsova, B.; Hoja, U.; Wimmelbacher, M.; Greiner, E.; Ustun, S.; Melzer, M.; Petersen, K.; Lein, W.; Bornke, F. Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent manner: Evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana. Plant Cell 2010, 22, 1498–1515. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, C.V.; Rey, P. Plant thioredoxins are key actors in the oxidative stress response. Trends Plant Sci. 2006, 11, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Rivas, S.; Rougon-Cardoso, A.; Smoker, M.; Schauser, L.; Yoshioka, H.; Jones, J.D.G. CITRX thioredoxin interacts with the tomato Cf-9 resistance protein and negatively regulates defence. EMBO J. 2004, 23, 2156–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, C.; Wang, M.H. NADPH-dependent thioredoxin reductase from tomato (Solanum lycopersicum). BMC Rep. 2011, 44, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.; Chaban, C.; Schütze, K.; Batistic, O.; Weckermann, K.; Näke, C.; Blazevic, D.; Grefen, C.; Schumacher, K.; Oecking, C.; et al. Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J. 2004, 40, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Tada, Y.; Spoel, S.H.; Pajerowska-Mukhtar, K.; Mou, Z.; Song, J.; Wang, C.; Zuo, J.; Dong, X. Plant immunity requires conformational changes of NPR1 via S-nitrosylation and thioredoxins. Science 2008, 321, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-B.; Han, L.-B.; Wang, H.-Y.; Zhang, J.; Sun, S.-T.; Feng, D.-Q.; Yang, C.-L.; Sun, Y.-D.; Zhong, N.-Q.; Xia, G.-X. The thioredoxin GbNRX1 plays a crucial role in homeostasis of apoplastic reactive oxygen species in response to Verticillium dahliae infection in cotton. Plant Physiol. 2016, 170, 2392–2406. [Google Scholar] [CrossRef] [PubMed]
- Mukaihara, T.; Hatanaka, T.; Nakano, M.; Oda, K. Ralstonia solanacearum type III effector RipAY is a glutathione-degrading enzyme that is activated by plant cytosolic thioredoxins and suppresses plant immunity. mBio 2016, 7, e00359-16. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Ren, H.; Liu, R.; Li, B.; Wu, T.; Sun, F.; Liu, H.; Wang, X.; Dong, H. An h-type thioredoxin functions in tobacco defense responses to two species of viruses and an abiotic oxidative stress. Mol. Plant-Microbe Interact. 2010, 23, 1470–1485. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, H.; Gong, Y.; Tao, Y.; Jiang, L.; Zuo, W.; Yang, Q.; Ye, J.; Lai, J.; Wu, J.; et al. An Atypical Thioredoxin Imparts Early Resistance to Sugarcane Mosaic Virus in Maize. Mol. Plant 2017, 10, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Luna-Rivero, M.S.; Hernandez-Zepeda, C.; Villanueva-Alonzo, H.; Minero-Garcia, Y.; Castel-Gonzales, S.E.; Moreno-Valenzuela, O.A. Expression of genes involved in the salicylic acid pathway in type h1 thioredoxin transiently silenced pepper plants during a begomovirus compatible interaction. Mol. Genet. Genom. 2016, 29, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Castellano, M.M.; Sablowski, R. Phosducin-like protein 3 is required for microtubule-dependent steps of cell division but not for meristem, growth in Arabidopsis. Plant Cell 2008, 20, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. PLP3 proteins function in microtubule assembly in Arabidopsis. Plant Cell 2008, 20, 821. [Google Scholar] [CrossRef]
- Verchot-Lubicz, J.; Torrance, L.; Solovyev, A.G.; Morozov, S.Y.; Jackson, A.O.; Gilmer, D. Varied movement strategies employed by triple gene block-encoding viruses. Mol. Plant-Microbe Interact. 2010, 23, 1231–1247. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Peña, E.J.; Amari, K.; Heinlein, M. Microtubules in viral replication and transport. Plant J. 2013, 75, 290–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiwatari, Y.; Fujiwara, T.; McFarland, K.C.; Nemoto, K.; Hayashi, H.; Chino, M.; Lucas, W.J. Rice phloem thioredoxin h has the capacity to mediate its own cell-to-cell transport through plasmodesmata. Planta 1998, 205, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Wong, J.H.; Feldman, L.J.; Lemaux, P.G.; Buchanan, B.B. A membrane-associated thioredoxin required for plant growth moves from cell to cell, suggestive of a role in intercellular communication. Proc. Natl. Acad. Sci. USA 2010, 107, 3900–3905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Lob, Y.T.; Bressan, R.A.; Martin, G.B. The tomato gene PtiI encodes a serine/threonine kinase that is phosphorylated by Pto and is involved in the hypersensitive response. Cell 1995, 83, 925–935. [Google Scholar] [CrossRef]
- Gyuris, J.; Golemis, E.; Cherikov, H.; Brent, R. CDI1, a human G1-phase and S-phase protein phosphate that associates with CDK2. Cell 1993, 75, 791–803. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: New York, NY, USA, 1965; pp. 97–166. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Rotenberg, D.; Thompson, T.S.; German, T.L.; Willis, D.K. Methods for effective real-time RT-PCR analysis of virus-induced gene silencing. J. Virol. Meth. 2006, 138, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Donald, R.G.; Zhou, H.; Jackson, A.O. Serological analysis of barley stripe mosaic virus-encoded proteins in infected barley. Virology 1993, 195, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Medina, V.; Tian, T.; Wierzchos, J.; Falk, B.W. Specific inclusion bodies are associated with replication of lettuce infectious yellows virus RNAs in Nicotiana benthamiana protoplasts. J. Gen. Virol. 1998, 79, 2325–2329. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathioudakis, M.M.; Khechmar, S.; Owen, C.A.; Medina, V.; Ben Mansour, K.; Tomaszewska, W.; Spanos, T.; Sarris, P.F.; Livieratos, I.C. A Thioredoxin Domain-Containing Protein Interacts with Pepino mosaic virus Triple Gene Block Protein 1. Int. J. Mol. Sci. 2018, 19, 3747. https://doi.org/10.3390/ijms19123747
Mathioudakis MM, Khechmar S, Owen CA, Medina V, Ben Mansour K, Tomaszewska W, Spanos T, Sarris PF, Livieratos IC. A Thioredoxin Domain-Containing Protein Interacts with Pepino mosaic virus Triple Gene Block Protein 1. International Journal of Molecular Sciences. 2018; 19(12):3747. https://doi.org/10.3390/ijms19123747
Chicago/Turabian StyleMathioudakis, Matthaios M., Souheyla Khechmar, Carolyn A. Owen, Vicente Medina, Karima Ben Mansour, Weronika Tomaszewska, Theodore Spanos, Panagiotis F. Sarris, and Ioannis C. Livieratos. 2018. "A Thioredoxin Domain-Containing Protein Interacts with Pepino mosaic virus Triple Gene Block Protein 1" International Journal of Molecular Sciences 19, no. 12: 3747. https://doi.org/10.3390/ijms19123747
APA StyleMathioudakis, M. M., Khechmar, S., Owen, C. A., Medina, V., Ben Mansour, K., Tomaszewska, W., Spanos, T., Sarris, P. F., & Livieratos, I. C. (2018). A Thioredoxin Domain-Containing Protein Interacts with Pepino mosaic virus Triple Gene Block Protein 1. International Journal of Molecular Sciences, 19(12), 3747. https://doi.org/10.3390/ijms19123747