Reduced Corneal Innervation in the CD25 Null Model of Sjögren Syndrome †


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
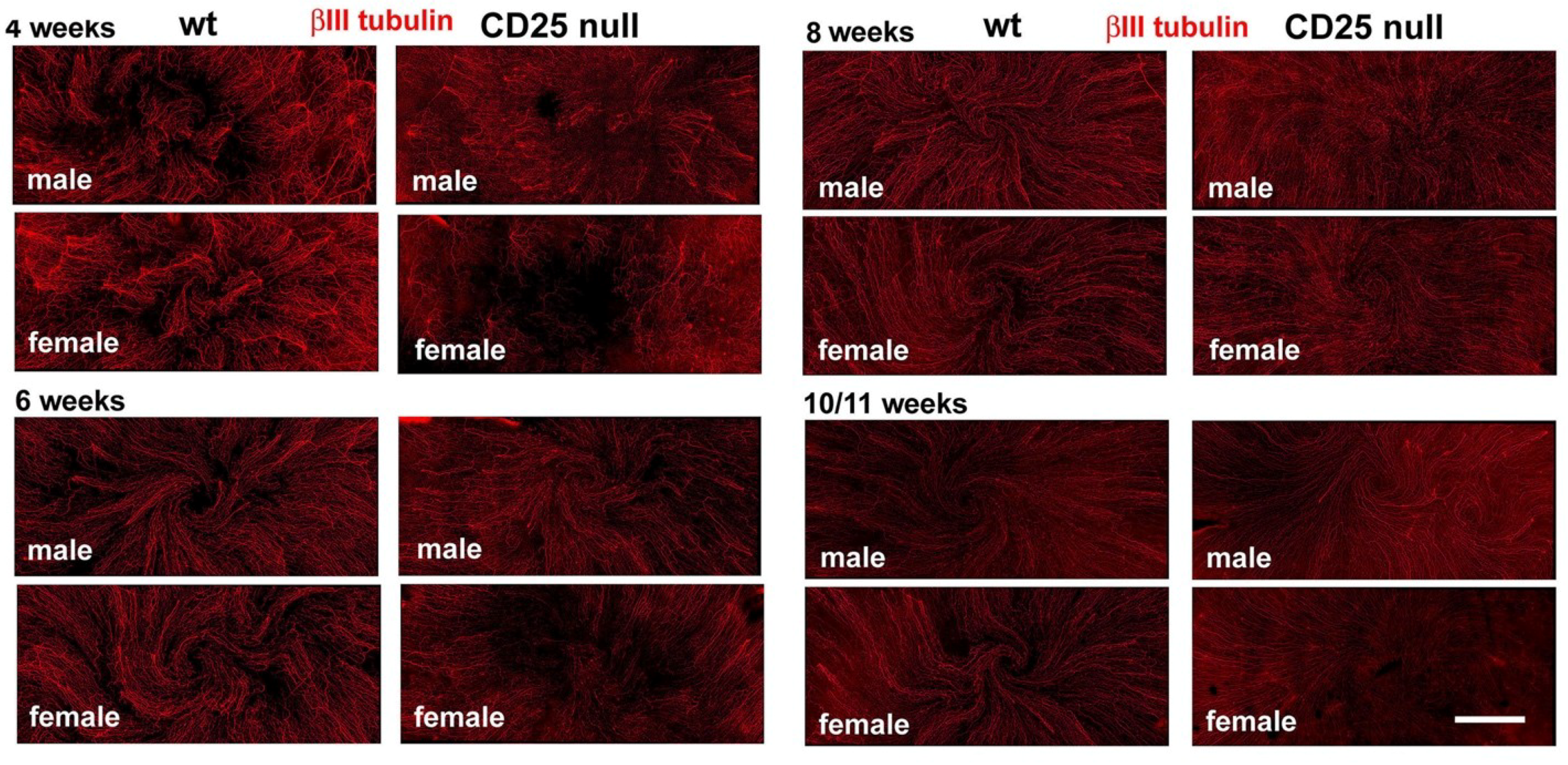
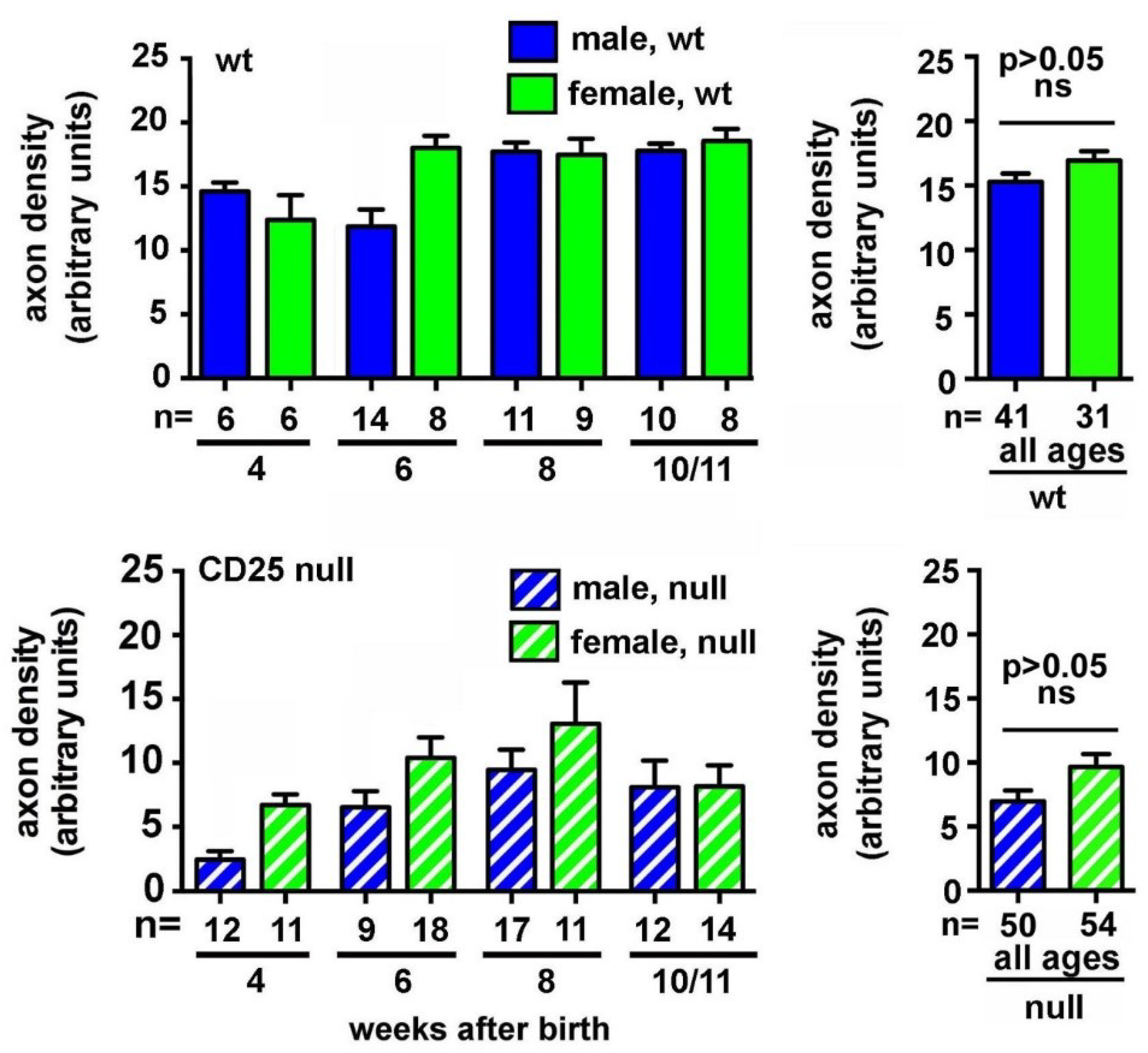
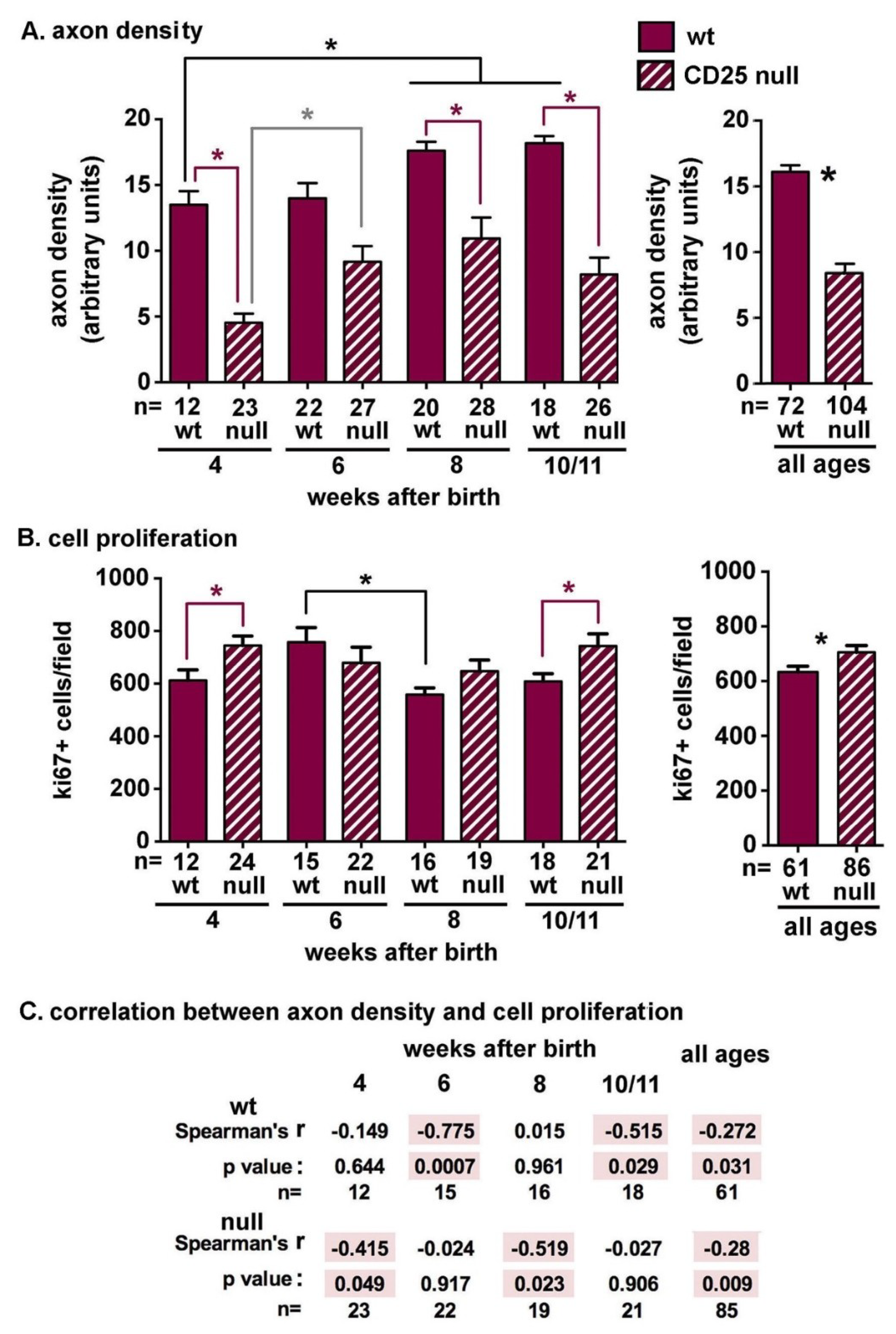
2.1. CD25 Null Mice Have Reduced Axon Density and Increased Corneal Epithelial Cell Proliferation as Early as Four Weeks after Birth
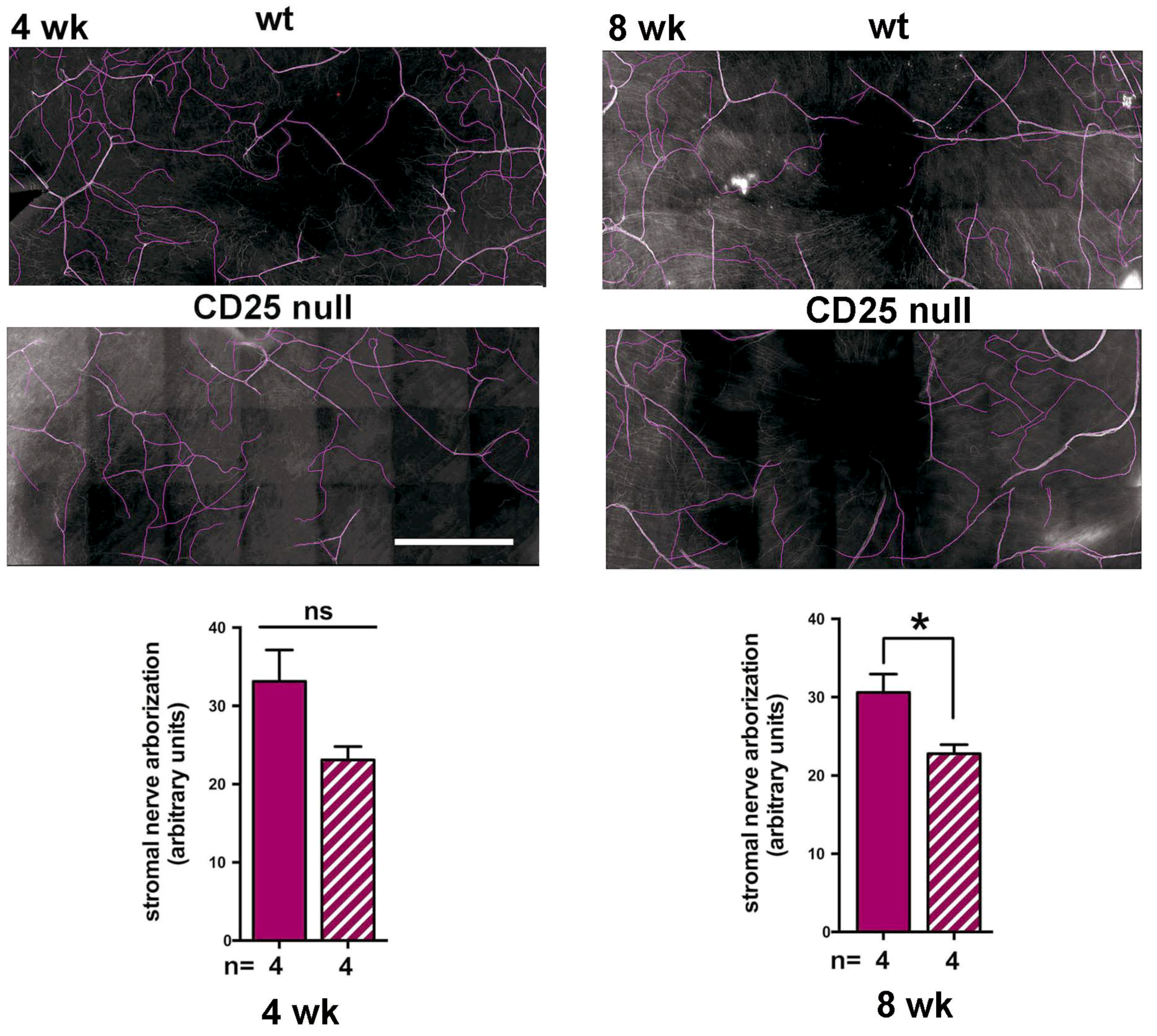
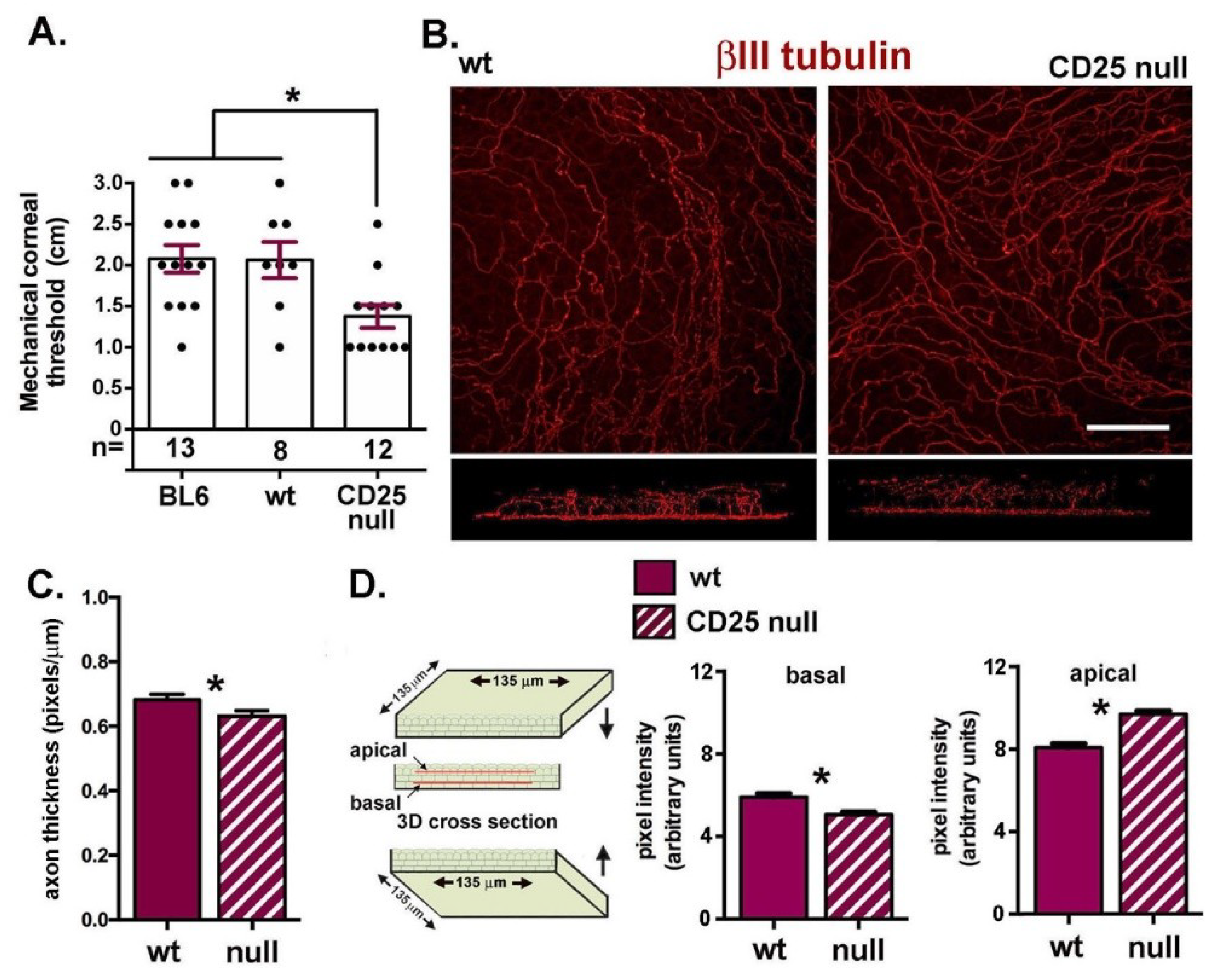
2.2. CD25 Null Corneas Have Fewer Stromal Axons
2.3. CD25 Null Corneas Have Reduced Corneal Sensitivity but Increased Apical Extension and Branching of Their Intraepithelial Nerve Terminals
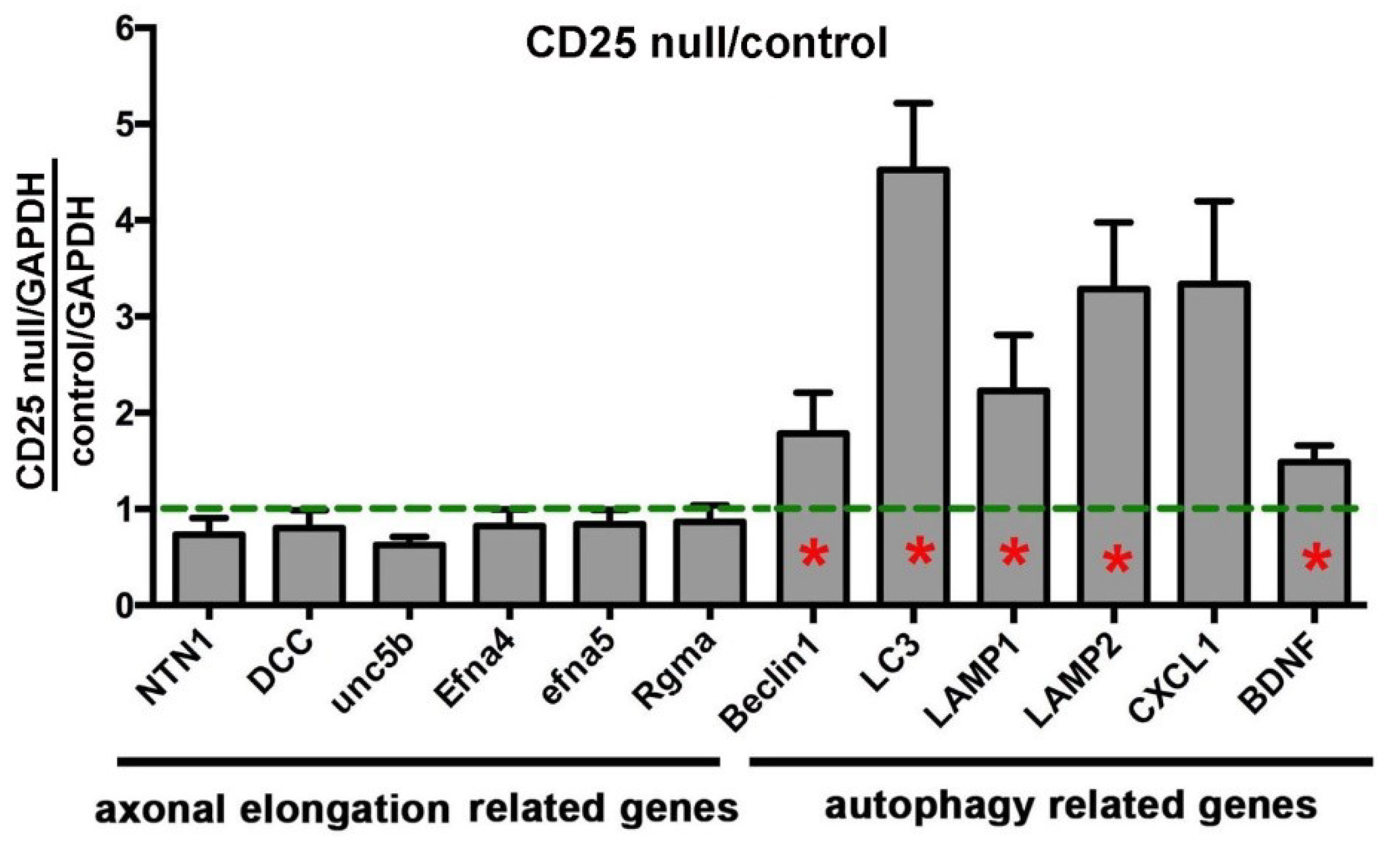
2.4. Loss of Axon Density and Reduced Corneal Sensitivity in CD25 Null Mice is Accompanied by Increased Corneal Epithelial Expression of mRNAs Encoding Proteins That Mediate Autophagy and Phagocytosis
3. Discussion
4. Methods
4.1. Animals
4.2. Corneal Mechanical Sensitivity
4.3. Immunofluorescence
4.4. Microscopy
4.5. qPCR
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grant, C.R.; Liberal, R.; Mieli-Vergani, G.; Vergani, D.; Longhi, M.S. Regulatory T-cells in autoimmune diseases: Challenges, controversies and—yet—unanswered questions. Autoimmun. Rev. 2015, 14, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Caldirola, M.S.; Broggi, M.G.R.; Gaillard, M.I.; Bezrodnik, L.; Zwirner, N.W. Primary Immunodeficiencies Unravel the Role of IL-2/CD25/STAT5b in Human Natural Killer Cell Maturation. Front. Immunol. 2018, 9, 1429. [Google Scholar] [CrossRef] [PubMed]
- Sharfe, N.; Dadi, H.K.; Shahar, M.; Roifman, C.M. Human immune disorder arising from mutation of the alpha chain of the interleukin-2 receptor. Proc. Natl. Acad. Sci. USA 1997, 94, 3168–3171. [Google Scholar] [CrossRef] [PubMed]
- Caudy, A.A.; Reddy, S.T.; Chatila, T.; Atkinson, J.P.; Verbsky, J.W. CD25 deficiency causes an immune dysregulation, polyendocrinopathy, enteropathy, X-linked–like syndrome, and defective IL-10 expression from CD4 lymphocytes. J. Allergy Clin. Immunol. 2007, 119, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Goudy, K.; Aydin, D.; Barzaghi, F.; Gambineri, E.; Vignoli, M.; Mannurita, S.C.; Doglioni, C.; Ponzoni, M.; Cicalese, M.P.; Assanelli, A.; et al. Human IL2RA null mutation mediates immunodeficiency with lymphoproliferation and autoimmunity. Clin. Immunol. 2013, 146, 248–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezrodnik, L.; Caldirola, M.S.; Seminario, A.G.; Moreira, I.; Gaillard, M.I. Follicular bronchiolitis as phenotype associated with CD25 deficiency. Clin. Exp. Immunol. 2014, 175, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayraktutar, B.N.; Baş, Z.; Uçakhan, Ö.Ö. Severe Dry Eye in CD25 Deficiency Syndrome. Eye Contact Lens: Sci. Clin. Pract. 2017, 44, e1–e3. [Google Scholar] [CrossRef]
- Foulsham, W.; Coco, G.; Amouzegar, A.; Chauhan, S.K.; Dana, R. When Clarity Is Crucial: Regulating Ocular Surface Immunity. Trends Immunol. 2018, 39, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Niederkorn, J.Y.; Stern, M.E.; Pflugfelder, S.C.; De Paiva, C.; Corrales, R.M.; Gao, J.; Siemasko, K. Desiccating Stress Induces T Cell-Mediated Sjogren’s Syndrome-Like Lacrimal Keratoconjunctivitis. J. Immunol. 2006, 176, 3950–3957. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.K.; El Annan, J.; Ecoiffier, T.; Goyal, S.; Zhang, Q.; Saban, D.R.; Dana, R. Autoimmunity in Dry Eye Is Due to Resistance of Th17 to Treg Suppression. J. Immunol. 2009, 182, 1247–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coursey, T.G.; Bian, F.; Zaheer, M.; Pflugfelder, S.C.; Volpe, E.A.; De Paiva, C. Age-related spontaneous lacrimal keratoconjunctivitis is accompanied by dysfunctional T regulatory cells. Mucosal Immunol. 2016, 10, 743–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratay, M.L.; Glowacki, A.J.; Balmert, S.C.; Acharya, A.P.; Polat, J.; Andrews, L.P.; Fedorchak, M.V.; Schuman, J.S.; Vignali, D.A.; Little, S.R. Treg-recruiting microspheres prevent inflammation in a murine model of dry eye disease. J. Control. Release 2017, 258, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Rahimy, E.; Pitcher, J.D.; Pangelinan, S.B.; Farley, W.J.; Li, D.-Q.; Pflugfelder, S.C.; Chen, W.; Niederkorn, J.Y.; Stern, M.E.; De Paiva, C.S. Spontaneous Autoimmune Dacryoadenitis in Aged CD25KO Mice Spontaneous Autoimmune Dacryoadenitis in Aged CD25KO Mice. Am. J. Pathol. 2010, 177, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Pelegrino, F.S.A.; Volpe, E.A.; Gandhi, N.B.; Li, D.-Q.; Pflugfelder, S.C.; Pelegrino, F.S.; De Paiva, C.S. Deletion of interferon-γ delays onset and severity of dacryoadenitis in CD25KO mice. Arthritis Res. Ther. 2012, 14, R234. [Google Scholar] [CrossRef] [PubMed]
- De Paiva, C.S.; Hwang, C.S.; Pitcher, J.D., 3rd; Pangelinan, S.B.; Rahimy, E.; Chen, W.; Yoon, K.C.; Farley, W.J.; Niederkorn, J.Y.; Stern, M.E.; et al. Age-related T-cell cytokine profile parallels corneal disease severity in Sjogren’s syndrome-like keratoconjunctivitis sicca in CD25KO mice. Rheumatology 2010, 49, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Bian, F.; Barbosa, F.L.; Corrales, R.M.; Pelegrino, F.S.; Volpe, E.A.; Pflugfelder, S.C.; De Paiva, C.S. Altered balance of interleukin-13/interferon-gamma contributes to lacrimal gland destruction and secretory dysfunction in CD25 knockout model of Sjögren’s syndrome. Arthritis Res. Ther. 2015, 17, 53. [Google Scholar] [CrossRef] [PubMed]
- Stepp, M.A.; Tadvalkar, G.; Hakh, R.; Pal-Ghosh, S. Corneal epithelial cells function as surrogate Schwann cells for their sensory nerves. Glia 2016, 65, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Labetoulle, M.; Baudouin, C.; Calonge, M.; Merayo-Lloves, J.; Boboridis, K.G.; Akova, Y.A.; Aragona, P.; Geerling, G.; Messmer, E.M.; Benítez-Del-Castillo, J. Role of corneal nerves in ocular surface homeostasis and disease. Acta Ophthalmol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, G.; Chauhan, S.K.; Ueno, H.; Nallasamy, N.; Gandolfi, S.; Borges, L.; Dana, R. A Novel Mouse Model for Neurotrophic Keratopathy: Trigeminal Nerve Stereotactic Electrolysis through the Brain. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2532–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bignami, F.; Ferrari, G.; Ueno, H.; Rama, P.; Dana, R. Trigeminal stereotactic electrolysis induces dry eye in mice. Acta Ophthalmol. 2012, 91, e162–e163. [Google Scholar] [CrossRef]
- Alcalde, I.; Íñigo-Portugués, A.; Gonzalez-Gonzalez, O.; Almaraz, L.; Artime, E.; Morenilla-Palao, C.; Gallar, J.; Viana, F.; Merayo-Lloves, J.; Belmonte, C. Morphological and functional changes in TRPM8-expressing corneal cold thermoreceptor neurons during aging and their impact on tearing in mice. J. Comp. Neurol. 2018, 526, 1859–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonen, B.; Alpizar, Y.A.; Meseguer, V.M.; Talavera, K. TRP Channels as Sensors of Bacterial Endotoxins. Toxins 2018, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Labbé, A.; Liang, Q.; Wang, Z.; Zhang, Y.; Xu, L.; Baudouin, C.; Sun, X. Corneal Nerve Structure and Function in Patients with Non-Sjögren Dry Eye: Clinical Correlations. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5144–5150. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Irvine, J.; Tepelus, T.C.; Chiu, G.B.; Sadda, S.R.; Lee, O.L. Correlation between corneal innervation and inflammation evaluated with confocal microscopy and symptomatology in patients with dry eye syndromes: A preliminary study. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 1771–1778. [Google Scholar] [CrossRef]
- Seo, K.Y.; Kitamura, K.; Han, S.J.; Kelsall, B. TH17 cells mediate inflammation in a novel model of spontaneous experimental autoimmune lacrimal keratoconjunctivitis with neural damage. J. Allergy Clin. Immunol. 2018, 142, 96–108.e2. [Google Scholar] [CrossRef] [PubMed]
- Simsek, C.; Kojima, T.; Doğru, M.; Tsubota, K. Alterations of Murine Subbasal Corneal Nerves after Environmental Dry Eye Stress. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1986–1995. [Google Scholar] [CrossRef] [PubMed]
- Stepp, M.A.; Pal-Ghosh, S.; Tadvalkar, G.; Williams, A.; Pflugfelder, S.C.; De Paiva, C.S. Reduced intraepithelial corneal nerve density and sensitivity accompany desiccating stress and aging in C57BL/6 mice. Exp. Eye Res. 2018, 169, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal-Ghosh, S.; Pajoohesh-Ganji, A.; Tadvalkar, G.; Kyne, B.M.; Guo, X.; Zieske, J.D.; Stepp, M.A. Topical Mitomycin-C enhances subbasal nerve regeneration and reduces erosion frequency in the debridement wounded mouse cornea. Exp. Eye Res. 2016, 146, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal-Ghosh, S.; Tadvalkar, G.; Pajoohesh-Ganji, A.; Stepp, M.A. Removal of the basement membrane enhances corneal wound healing. Exp. Eye Res. 2011, 93, 927–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, K.-C.; De Paiva, C.; Qi, H.; Chen, Z.; Farley, W.J.; Li, D.-Q.; Pflugfelder, S.C. Expression of Th-1 Chemokines and Chemokine Receptors on the Ocular Surface of C57BL/6 Mice: Effects of Desiccating Stress. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2561–2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paiva, C.S.; Yoon, Ky.; Pangelinan, S.B.; Pham, S.; Puthenparambil, L.M.; Chuang, E.Y.; Farley, W.J.; Stern, M.E.; Li, De.; Pflugfelder, S.C. Cleavage of functional IL-2 receptor alpha chain (CD25) from murine corneal and conjunctival epithelia by MMP-9. J. Inflamm. 2009, 6, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepp, M.A.; Pal-Ghosh, S.; Tadvalkar, G.; Li, L.; Brooks, S.R.; Morasso, M.I. Molecular basis of Mitomycin C enhanced corneal sensory nerve repair after debridement wounding. Sci. Rep. 2018, 8, 16960. [Google Scholar] [CrossRef] [PubMed]
- Stepp, M.A.; Pal-Ghosh, S.; Tadvalkar, G.; Pajoohesh-Ganji, A. Syndecan-1 and Its Expanding List of Contacts. Adv. Wound Care 2015, 4, 235–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchette, C.R.; Perrat, P.N.; Thackeray, A.; Benard, C.Y. Glypican Is a Modulator of Netrin-Mediated Axon Guidance. PLoS Biol. 2015, 13, e1002183. [Google Scholar] [CrossRef] [PubMed]
- Gysi, S.; Rhiner, C.; Flibotte, S.; Moerman, D.G.; Hengartner, M.O. A Network of HSPG Core Proteins and HS Modifying Enzymes Regulates Netrin-Dependent Guidance of D-Type Motor Neurons in Caenorhabditis elegans. PLoS ONE 2013, 8, e74908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheller, J.; Chalaris, A.; Garbers, C.; Rose-John, S. ADAM17: A molecular switch to control inflammation and tissue regeneration. Trends Immunol. 2011, 32, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Cunha, L.D.; Park, S.; Yang, M.; Lü, Q.; Orchard, R.; Li, Q.-Z.; Yan, M.; Janke, L.; Guy, C.; et al. Noncanonical autophagy inhibits the autoinflammatory, lupus-like response to dying cells. Nature 2016, 533, 115–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J. LAP it up, fuzz ball: A short history of LC3-associated phagocytosis. Curr. Opin. Immunol. 2018, 55, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.; Sil, P.; Martinez, J. Rubicon: LC3-associated phagocytosis and beyond. FEBS J. 2017, 285, 1379–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muniz-Feliciano, L.; Doggett, T.A.; Zhou, Z.; Ferguson, T.A. RUBCN/rubicon and EGFR regulate lysosomal degradative processes in the retinal pigment epithelium (RPE) of the eye. Autophagy 2017, 13, 2072–2085. [Google Scholar] [CrossRef] [PubMed]
- Byun, Y.-S.; Lee, H.J.; Shin, S.; Chung, S.-H. Elevation of autophagy markers in Sjögren syndrome dry eye. Sci. Rep. 2017, 7, 17280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajoohesh-Ganji, A.; Pal-Ghosh, S.; Tadvalkar, G.; Kyne, B.M.; Saban, D.R.; Stepp, M.A. Partial denervation of sub-basal axons persists following debridement wounds to the mouse cornea. Lab. Investig. 2015, 95, 1305–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal-Ghosh, S.; Tadvalkar, G.; Stepp, M.A. Alterations in Corneal Sensory Nerves During Homeostasis, Aging, and After Injury in Mice Lacking the Heparan Sulfate Proteoglycan Syndecan-1. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4959–4975. [Google Scholar] [CrossRef] [PubMed]
- Meijering, E.; Jacob, M.; Sarria, J.-C.; Steiner, P.; Hirling, H.; Unser, M.; Sarria, J. Design and validation of a tool for neurite tracing and analysis in fluorescence microscopy images. Cytometry 2004, 58, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stepp, M.A.; Pal-Ghosh, S.; Tadvalkar, G.; Williams, A.R.; Pflugfelder, S.C.; De Paiva, C.S. Reduced Corneal Innervation in the CD25 Null Model of Sjögren Syndrome. Int. J. Mol. Sci. 2018, 19, 3821. https://doi.org/10.3390/ijms19123821
Stepp MA, Pal-Ghosh S, Tadvalkar G, Williams AR, Pflugfelder SC, De Paiva CS. Reduced Corneal Innervation in the CD25 Null Model of Sjögren Syndrome. International Journal of Molecular Sciences. 2018; 19(12):3821. https://doi.org/10.3390/ijms19123821
Chicago/Turabian StyleStepp, Mary Ann, Sonali Pal-Ghosh, Gauri Tadvalkar, Alexa R. Williams, Stephen C. Pflugfelder, and Cintia S. De Paiva. 2018. "Reduced Corneal Innervation in the CD25 Null Model of Sjögren Syndrome" International Journal of Molecular Sciences 19, no. 12: 3821. https://doi.org/10.3390/ijms19123821
APA StyleStepp, M. A., Pal-Ghosh, S., Tadvalkar, G., Williams, A. R., Pflugfelder, S. C., & De Paiva, C. S. (2018). Reduced Corneal Innervation in the CD25 Null Model of Sjögren Syndrome. International Journal of Molecular Sciences, 19(12), 3821. https://doi.org/10.3390/ijms19123821