Inhibition of Ceramide De Novo Synthesis Affects Adipocytokine Secretion and Improves Systemic and Adipose Tissue Insulin Sensitivity

 , , , , ,
, , , , ,
Abstract
:
1. Introduction
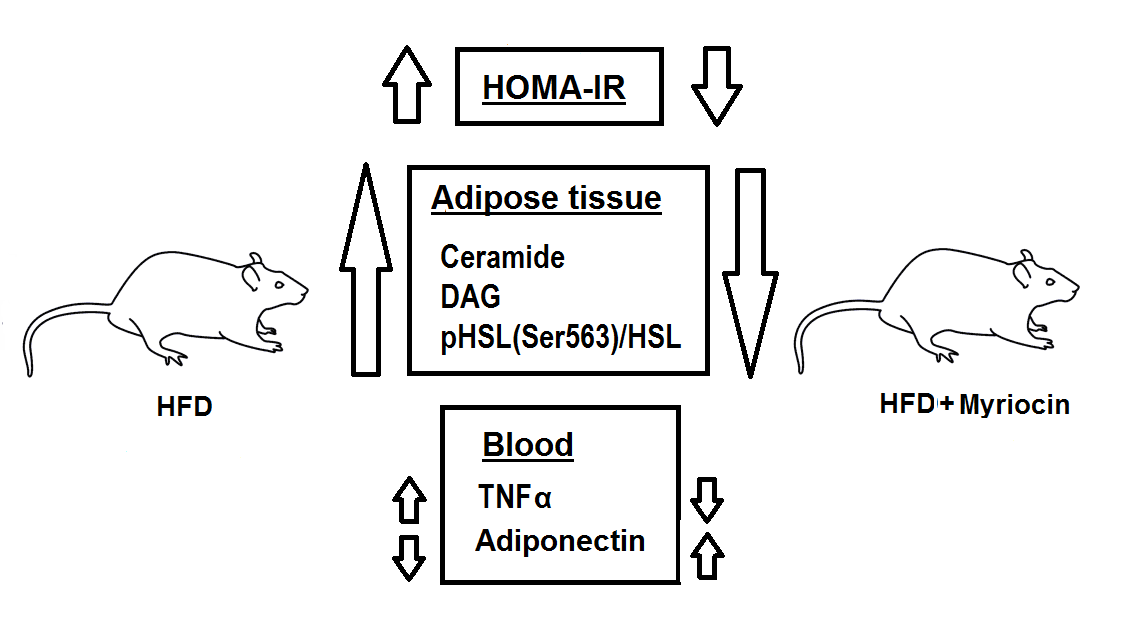
2. Results
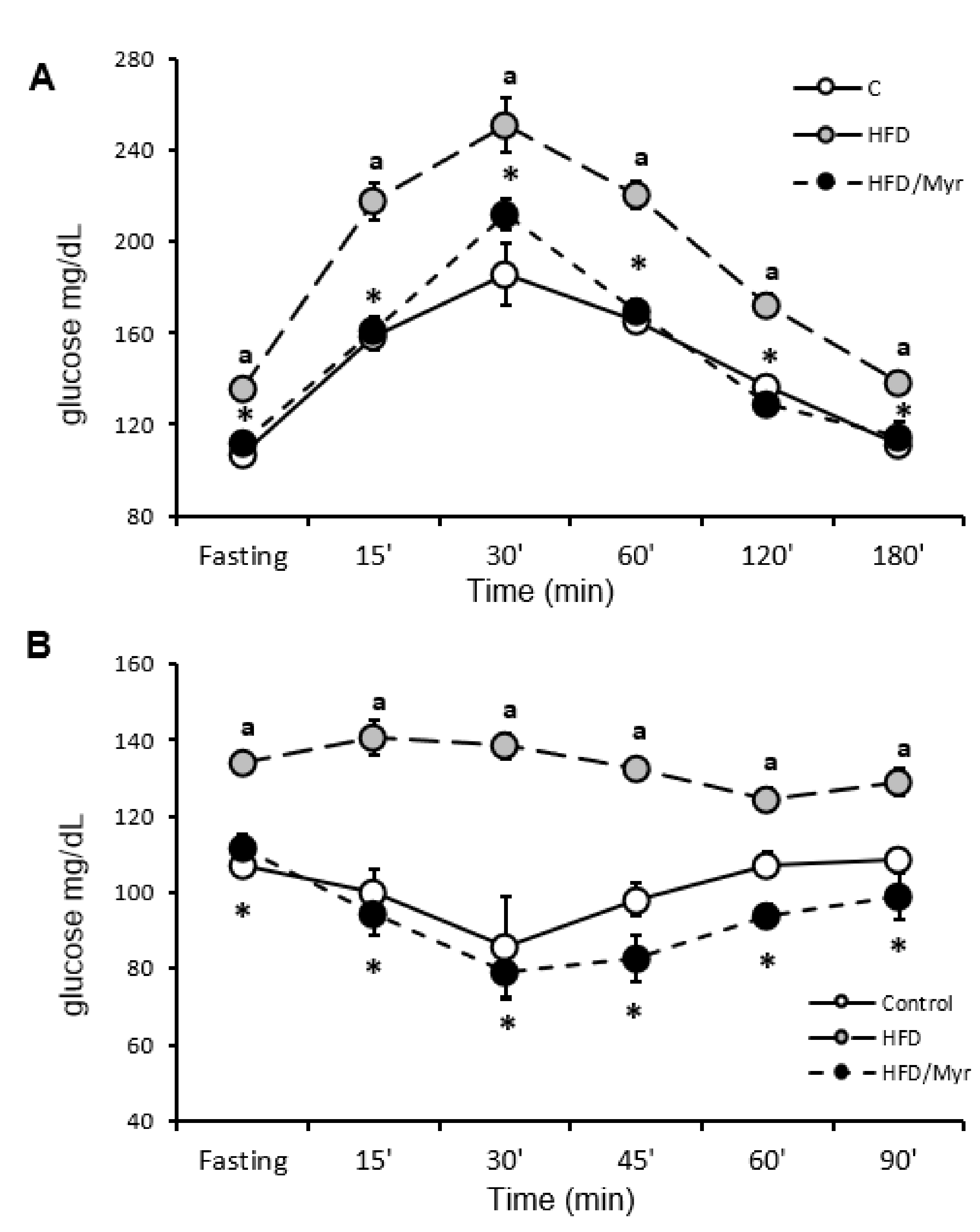
2.1. HOMA-IR, OGTT and ITT
2.2. Plasma FFAs
2.3. Plasma Adiponectin and TNF-α Concentration
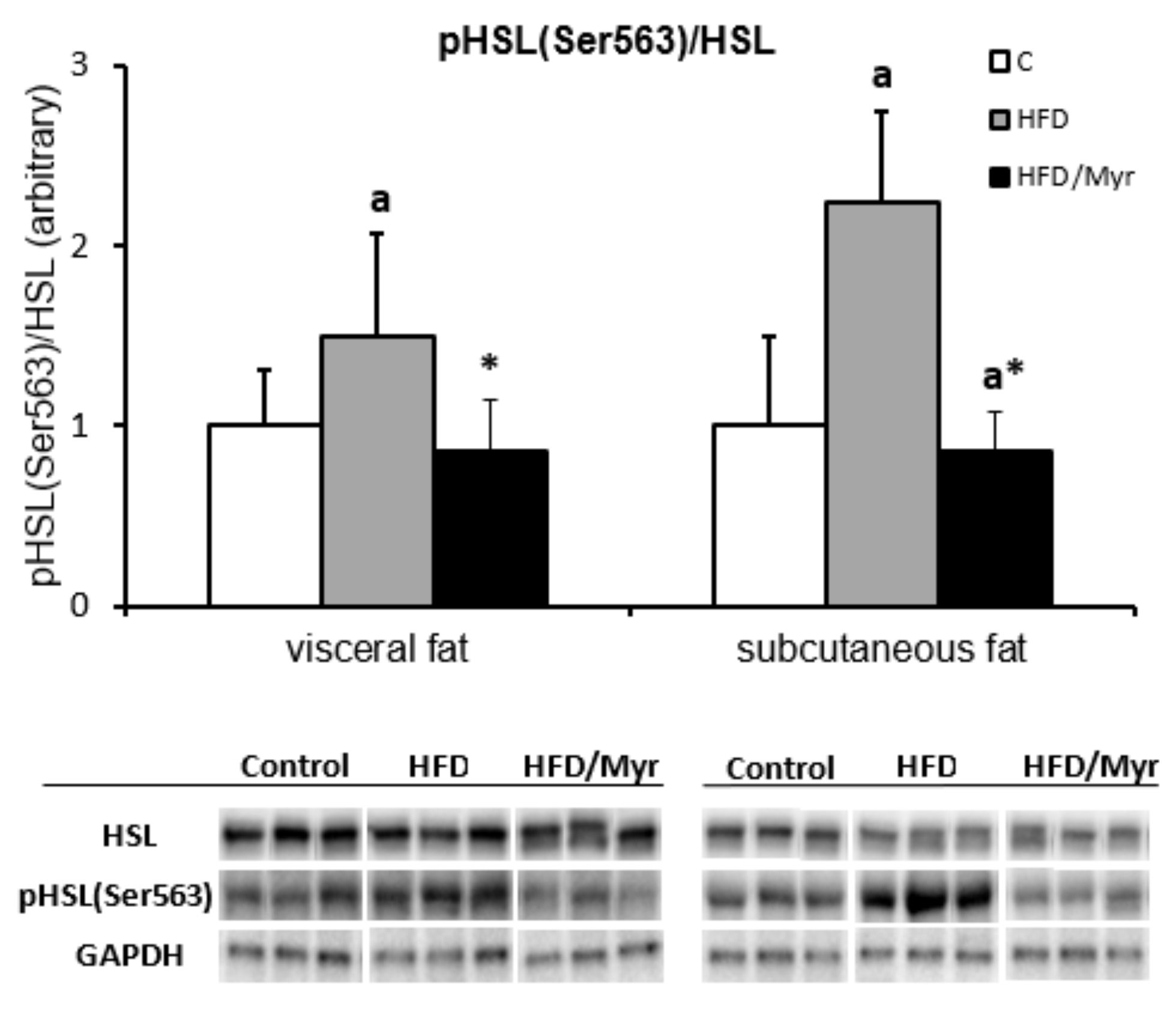
2.4. Hormone Sensitive Lipase pHSLSer563
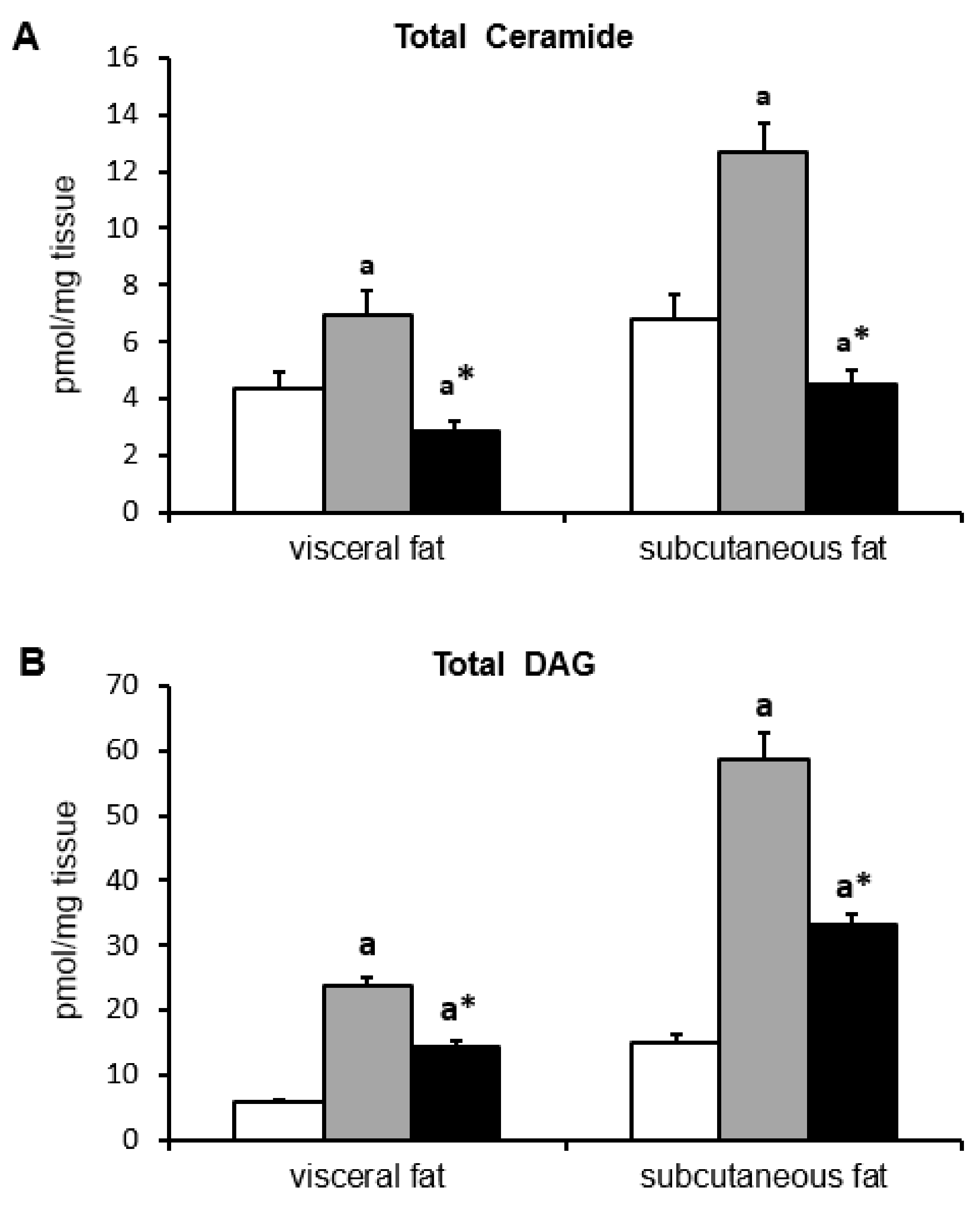
2.5. Ceramide and DAG Concentration
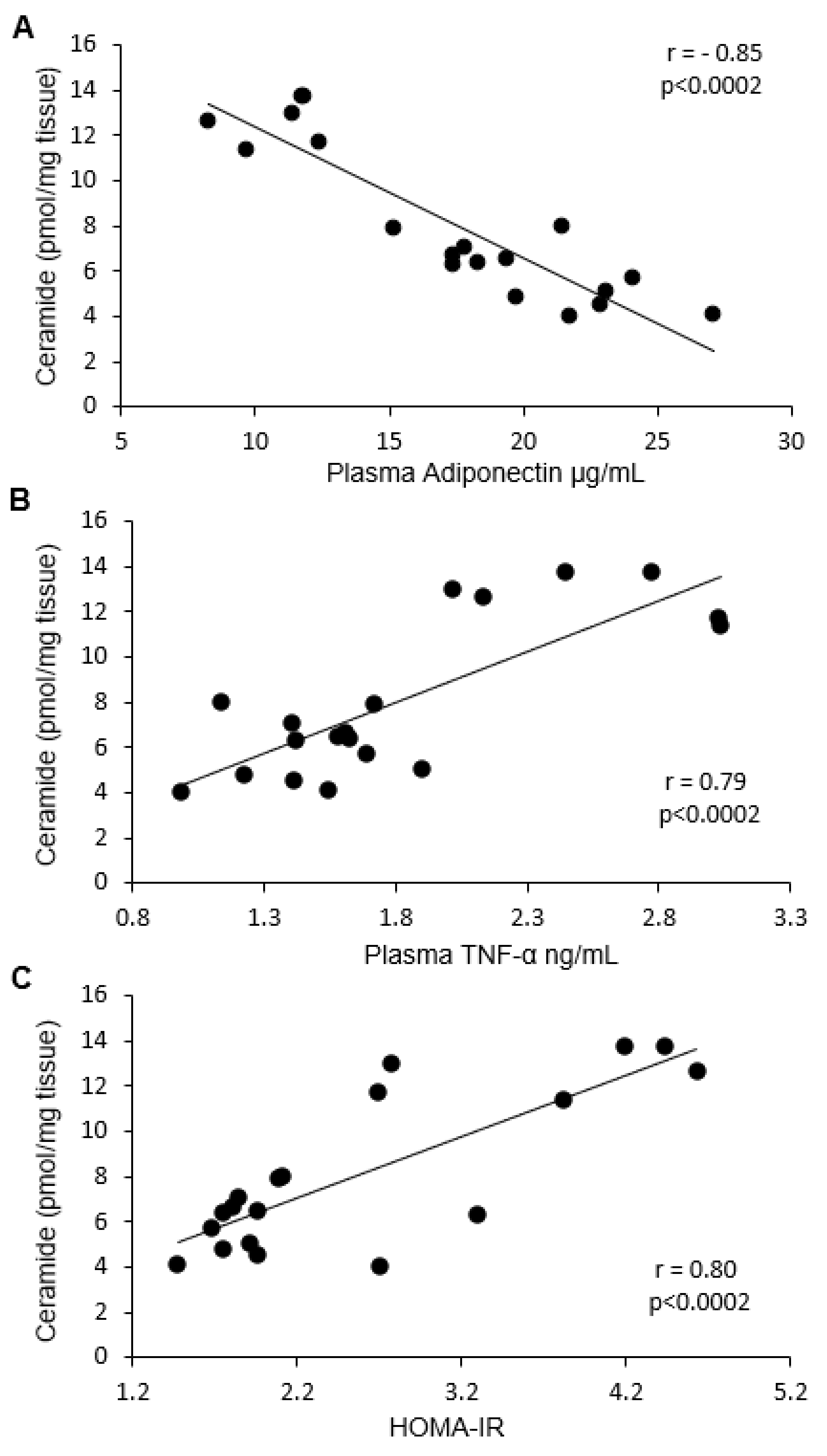
2.6. Correlation between Ceramide and DAG with HOMA-IR, Adiponectin, and TNF-α
3. Discussion
4. Materials and Methods
4.1. Animals and Study Design
4.2. Lipids Measurements
4.3. Western Blot
4.4. Plasma Adiponectin and TNF-α Concentration
4.5. Oral Glucose Tolerance Test (OGTT)
4.6. Insulin Tolerance Test (ITT)
4.7. Plasma Insulin and Glucose Concentration
4.8. HOMA-IR
4.9. Protein Concentration
4.10. Statistical Significance Estimation and Correlation Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Cer | Ceramide |
| DAG | Diacylglycerols |
| HFD | High fat diet |
| IRes | Insulin resistance |
| OGTT | Oral glucose tolerance test |
| ITT | Insulin tolerance test |
| HSL | Hormone sensitive lipase |
| HOMA-IR | Homeostatic Model Assessment for Insulin Resistance |
| LC/MS/MS | Liquid chromatography tandem mass spectrometry |
| VAT | Visceral adipose tissue |
| SAT | Subcutaneous adipose tissue |
| FFA | Free fatty acids |
References
- Fernández-Real, J.M.; Ricart, W. Insulin resistance and chronic cardiovascular inflammatory syndrome. Endocr. Rev. 2003, 24, 278–301. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Engeli, S.; Gorzelniak, K.; Kreutz, R.; Runkel, N.; Distler, A.; Sharma, A.M. Co-expression of renin-angiotensin system genes in human adipose tissue. J. Hypertens. 1999, 17, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Winkler, G.; Kiss, S.; Keszthelyi, L.; Sápi, Z.; Ory, I.; Salamon, F.; Kovács, M.; Vargha, P.; Szekeres, O.; Speer, G.; et al. Expression of tumor necrosis factor (TNF)-alpha protein in the subcutaneous and visceral adipose tissue in correlation with adipocyte cell volume, serum TNF-alpha, soluble serum TNF-receptor-2 concentrations and C-peptide level. Eur. J. Endocrinol. 2003, 149, 129–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A novel serum protein similar to C1q, produced exclusively in adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Arner, P.; Caro, J.F.; Atkinson, R.L.; Spiegelman, B.M. Increased adipose tissue expression of tumor necrosis factor-alpha in human obesity and insulin resistance. J. Clin. Investig. 1995, 95, 2409–2415. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Murray, D.L.; Choy, L.N.; Spiegelman, B.M. Tumor necrosis factor alpha inhibits signaling from the insulin receptor. Proc. Natl. Acad. Sci. USA 1994, 91, 4854–4858. [Google Scholar] [CrossRef]
- Clarke, C.J.; Cloessner, E.A.; Roddy, P.L.; Hannun, Y.A. Neutral sphingomyelinase 2 (nSMase2) is the primary neutral sphingomyelinase isoform activated by tumour necrosis factor-α in MCF-7 cells. Biochem. J. 2011, 435, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Kannel, W.B. Lipids, diabetes, and coronary heart disease: Insights from the Framingham Study. Am. Heart J. 1985, 110, 1100–1107. [Google Scholar] [CrossRef]
- Larsson, B. Obesity, fat distribution and cardiovascular disease. Int. J. Obes 1991, 15 (Suppl. 2), 53–57. [Google Scholar] [PubMed]
- Lee, M.J.; Wu, Y.; Fried, S.K. Adipose tissue heterogeneity: Implication of depot differences in adipose tissue for obesity complications. Mol. Aspects Med. 2013, 34, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Chen, Y.; Cline, G.W.; Zhang, D.; Zong, H.; Wang, Y.; Bergeron, R.; Kim, J.K.; Cushman, S.W.; Cooney, G.J.; et al. Mechanism by which fatty acids inhibit insulin activation of insulin receptor substrate-1 (IRS-1)-associated phosphatidylinositol 3-kinase activity in muscle. J. Biol. Chem. 2002, 277, 50230–50236. [Google Scholar] [CrossRef] [PubMed]
- Shulman, G.I. Cellular mechanisms of insulin resistance. J. Clin. Investig. 2000, 106, 171–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merrill, A.H. De novo sphingolipid biosynthesis: A necessary, but dangerous, pathway. J. Biol. Chem. 2002, 277, 25843–25846. [Google Scholar] [CrossRef] [PubMed]
- Straczkowski, M.; Kowalska, I.; Nikolajuk, A.; Dzienis-Straczkowska, S.; Kinalska, I.; Baranowski, M.; Zendzian-Piotrowska, M.; Brzezinska, Z.; Gorski, J. Relationship between insulin sensitivity and sphingomyelin signaling pathway in human skeletal muscle. Diabetes 2004, 53, 1215–1221. [Google Scholar] [CrossRef]
- Straczkowski, M.; Kowalska, I.; Baranowski, M.; Nikolajuk, A.; Otziomek, E.; Zabielski, P.; Adamska, A.; Blachnio, A.; Gorski, J.; Gorska, M. Increased skeletal muscle ceramide level in men at risk of developing type 2 diabetes. Diabetologia 2007, 50, 2366–2373. [Google Scholar] [CrossRef] [Green Version]
- Straczkowski, M.; Kowalska, I. The role of skeletal muscle sphingolipids in the development of insulin resistance. Rev. Diabet. Stud. 2008, 5, 13–24. [Google Scholar] [CrossRef]
- Turinsky, J.; O’Sullivan, D.M.; Bayly, B.P. 1,2-Diacylglycerol and ceramide levels in insulin-resistant tissues of the rat in vivo. J. Biol. Chem. 1990, 265, 16880–16885. [Google Scholar]
- Magkos, F.; Su, X.; Bradley, D.; Fabbrini, E.; Conte, C.; Eagon, J.C.; Varela, J.E.; Brunt, E.M.; Patterson, B.W.; Klein, S. Intrahepatic diacylglycerol content is associated with hepatic insulin resistance in obese subjects. Gastroenterology 2012, 142, 1444–1446. [Google Scholar] [CrossRef]
- Kumashiro, N.; Erion, D.M.; Zhang, D.; Kahn, M.; Beddow, S.A.; Chu, X.; Still, C.D.; Gerhard, G.S.; Han, X.; Dziura, J.; et al. Cellular mechanism of insulin resistance in nonalcoholic fatty liver disease. Proc. Natl. Acad. Sci. USA 2011, 108, 16381–16385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.J.; Samuel, V.T.; Petersen, K.F.; Shulman, G.I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature 2014, 510, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachnio-Zabielska, A.U.; Pulka, M.; Baranowski, M.; Nikolajuk, A.; Zabielski, P.; Gorska, M.; Gorski, J. Ceramide metabolism is affected by obesity and diabetes in human adipose tissue. J. Cell. Physiol. 2012, 227, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Blachnio-Zabielska, A.U.; Koutsari, C.; Tchkonia, T.; Jensen, M.D. Sphingolipid content of human adipose tissue: Relationship to adiponectin and insulin resistance. Obesity (Silver Spring) 2012, 20, 2341–2347. [Google Scholar] [CrossRef] [PubMed]
- Błachnio-Zabielska, A.U.; Baranowski, M.; Hirnle, T.; Zabielski, P.; Lewczuk, A.; Dmitruk, I.; Górski, J. Increased bioactive lipids content in human subcutaneous and epicardial fat tissue correlates with insulin resistance. Lipids 2012, 47, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Samad, F.; Hester, K.D.; Yang, G.; Hannun, Y.A.; Bielawski, J. Altered adipose and plasma sphingolipid metabolism in obesity: A potential mechanism for cardiovascular and metabolic risk. Diabetes 2006, 55, 2579–2587. [Google Scholar] [CrossRef] [PubMed]
- Samad, F.; Badeanlou, L.; Shah, C.; Yang, G. Adipose tissue and ceramide biosynthesis in the pathogenesis of obesity. Adv. Exp. Med. Biol. 2011, 721, 67–86. [Google Scholar] [CrossRef]
- Yang, G.; Badeanlou, L.; Bielawski, J.; Roberts, A.J.; Hannun, Y.A.; Samad, F. Central role of ceramide biosynthesis in body weight regulation, energy metabolism, and the metabolic syndrome. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E211–E224. [Google Scholar] [CrossRef] [Green Version]
- Belfort, R.; Mandarino, L.; Kashyap, S.; Wirfel, K.; Pratipanawatr, T.; Berria, R.; Defronzo, R.A.; Cusi, K. Dose-response effect of elevated plasma free fatty acid on insulin signaling. Diabetes 2005, 54, 1640–1648. [Google Scholar] [CrossRef]
- Boden, G.; Chen, X.; Ruiz, J.; White, J.V.; Rossetti, L. Mechanisms of fatty acid-induced inhibition of glucose uptake. J. Clin. Investig. 1994, 93, 2438–2446. [Google Scholar] [CrossRef]
- Boden, G. Role of fatty acids in the pathogenesis of insulin resistance and NIDDM. Diabetes 1997, 46, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; Lebed, B.; Schatz, M.; Homko, C.; Lemieux, S. Effects of acute changes of plasma free fatty acids on intramyocellular fat content and insulin resistance in healthy subjects. Diabetes 2001, 50, 1612–1617. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; Chen, X.; Capulong, E.; Mozzoli, M. Effects of free fatty acids on gluconeogenesis and autoregulation of glucose production in type 2 diabetes. Diabetes 2001, 50, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Boden, G. Free fatty acids-the link between obesity and insulin resistance. Endocr. Pract. 2001, 7, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Boden, G. Obesity and free fatty acids. Endocrinol. Metab. Clin. N. Am. 2008, 37, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Itani, S.I.; Ruderman, N.B.; Schmieder, F.; Boden, G. Lipid-induced insulin resistance in human muscle is associated with changes in diacylglycerol, protein kinase C, and IkappaB-alpha. Diabetes 2002, 51, 2005–2011. [Google Scholar] [CrossRef]
- Blachnio-Zabielska, A.U.; Chacinska, M.; Vendelbo, M.H.; Zabielski, P. The Crucial Role of C18-Cer in Fat-Induced Skeletal Muscle Insulin Resistance. Cell. Physiol. Biochem. 2016, 40, 1207–1220. [Google Scholar] [CrossRef] [Green Version]
- Zabielski, P.; Daniluk, J.; Hady, H.R.; Markowski, A.R.; Imierska, M.; Górski, J.; Blachnio-Zabielska, A.U. The effect of high-fat diet and inhibition of ceramide production on insulin action in liver. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef]
- Adams, J.M.; Pratipanawatr, T.; Berria, R.; Wang, E.; DeFronzo, R.A.; Sullards, M.C.; Mandarino, L.J. Ceramide content is increased in skeletal muscle from obese insulin-resistant humans. Diabetes 2004, 53, 25–31. [Google Scholar] [CrossRef]
- Chavez, J.A.; Summers, S.A. A ceramide-centric view of insulin resistance. Cell Metab. 2012, 15, 585–594. [Google Scholar] [CrossRef]
- Zabielski, P.; Chacinska, M.; Charkiewicz, K.; Baranowski, M.; Gorski, J.; Blachnio-Zabielska, A.U. Effect of metformin on bioactive lipid metabolism in insulin-resistant muscle. J. Endocrinol. 2017, 233, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Zabielski, P.; Hady, H.R.; Chacinska, M.; Roszczyc, K.; Gorski, J.; Blachnio-Zabielska, A.U. The effect of high fat diet and metformin treatment on liver lipids accumulation and their impact on insulin action. Sci. Rep. 2018, 8, 7249. [Google Scholar] [CrossRef]
- Kolak, M.; Westerbacka, J.; Velagapudi, V.R.; Wågsäter, D.; Yetukuri, L.; Makkonen, J.; Rissanen, A.; Häkkinen, A.M.; Lindell, M.; Bergholm, R.; et al. Adipose tissue inflammation and increased ceramide content characterize subjects with high liver fat content independent of obesity. Diabetes 2007, 56, 1960–1968. [Google Scholar] [CrossRef]
- Persson, X.M.; Blachnio-Zabielska, A.U.; Jensen, M.D. Rapid measurement of plasma free fatty acid concentration and isotopic enrichment using LC/MS. J. Lipid Res. 2010, 51, 2761–2765. [Google Scholar] [CrossRef] [Green Version]
- Blachnio-Zabielska, A.U.; Persson, X.M.; Koutsari, C.; Zabielski, P.; Jensen, M.D. A liquid chromatography/tandem mass spectrometry method for measuring the in vivo incorporation of plasma free fatty acids into intramyocellular ceramides in humans. Rapid Commun. Mass Spectrom 2012, 26, 1134–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachnio-Zabielska, A.U.; Zabielski, P.; Jensen, M.D. Intramyocellular diacylglycerol concentrations and [U-13C]palmitate isotopic enrichment measured by LC/MS/MS. J. Lipid Res. 2013, 54, 1705–1711. [Google Scholar] [CrossRef] [PubMed]
- Cacho, J.; Sevillano, J.; de Castro, J.; Herrera, E.; Ramos, M.P. Validation of simple indexes to assess insulin sensitivity during pregnancy in Wistar and Sprague-Dawley rats. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1269–E1276. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolic Parameter | Control | HFD | HFD/Myr |
|---|---|---|---|
| Fasting plasma glucose concentration [mg/dL] | 97.8 ± 14.3 | 139.2 ± 27.1 a | 86.6 ± 19.9 * |
| Fasting insulin concentration [µU/mL] | 51.7 ± 12.9 | 65.9 ± 9.6 | 55.8 ± 10.0 |
| HOMA-IR | 2.08 ± 0.6 | 3.75 ± 0.84 a | 1.96 ± 0.52 * |
| Plasma FFA concentration [µmol/L] | 279.1 ± 34.7 | 362.2 ± 24.4 a | 403.3 ± 49.3 * |
| Plasma adiponectin concentration [µg/mL] | 18.9 ± 2.8 | 10.8 ± 1.6 a | 22.9 ± 3.1 * |
| Plasma TNF-α concentration [ng/mL] | 1.52 ± 0.19 | 2.57 ± 0.4 a | 1.41 ± 0.40 * |
| Fatty Acid | Control | HFD | HFD/Myr |
|---|---|---|---|
| C14:0 | 13.2 ± 2.6 | 14.8 ± 2.6 | 15.7 ± 0.7 |
| C16:1 | 7.3 ± 1.4 | 2.0 ± 0.3 a | 2.5 ± 0.4 a |
| C18:2 | 64.9 ± 12.8 | 59.9 ± 3.2 | 102.8 ± 13.9 |
| C16:0 | 61.2 ± 8.7 | 60.7 ± 4.8 | 83.4 ± 13.5 * |
| C18:1 | 66.2 ± 14.2 | 45.6 ± 8.7 a | 68.7 ± 11.6 * |
| C18:0 | 36.6 ± 4.1 | 91.4 ± 10.5 a | 94.1 ± 15.3 * |
| C20:0 | 1.02 ± 0.2 | 4.3 ± 0.9 a | 3.1 ± 0.3 a,* |
| C22:0 | 4.5 ± 0.7 | 11.7 ± 0.9 a | 15.0 ± 2.4 a |
| C24:1 | 1.7 ± 0.4 | 2.3 ± 0.5 | 11.0 ± 0.4 a,* |
| C24:0 | 22.5 ± 3.0 | 69.4 ± 15.6 a | 6.8 ± 0.4 * |
| Visceral Fat Tissue | Subcutaneous Fat Tissue | |||||
|---|---|---|---|---|---|---|
| Ceramide | Control | HFD | HFD/Myr | Control | HFD | HFD/Myr |
| C14-Cer | 0.007 ± 0.001 | 0.014 ± 0.003 a | 0.003 ± 0.0005 a,* | 0.03 ± 0.006 | 0.04 ± 0.006 a | 0.01 ± 0.000 a,* |
| C16-Cer | 1.05 ± 0.19 | 1.82 ± 0.30 a | 1.36 ± 0.27 a,* | 2.4 ± 0.5 | 4.1 ± 0.6 a | 1.68 ± 0.37 a,* |
| C18:1-Cer | 0.008 ± 0.001 | 0.03 ± 0.006 a | 0.01 ± 0.002 a,* | 0.03 ± 0.01 | 0.08 ± 0.02 a | 0.03 ± 0.003 * |
| C18-Cer | 0.10 ± 0.02 | 0.56 ± 0.10 a | 0.20 ± 0.04 a,* | 0.74 ± 0.07 | 1.22 ± 0.18 a | 0.43 ± 0.06 a,* |
| C20-Cer | 0.09 ± 0.018 | 0.34 ± 0.09 a | 0.09 ± 0.01 a,* | 0.37 ± 0.10 | 0.58 ± 0.05 a | 0.15 ± 0.02 a,* |
| C22-Cer | 0.52 ± 0.08 | 0.63 ± 0.09 a | 0.22 ± 0.03 a,* | 0.65 ± 0.10 | 1.32 ± 0.3 a | 0.41 ± 0.06 a,* |
| C24:1-Cer | 0.53 ± 0.11 | 0.63 ± 0.09 a | 0.38 ± 0.06 a,* | 0.91 ± 0.16 | 1.27 ± 0.3 a | 0.74 ± 0.08 a,* |
| C24-Cer | 2.02 ± 0.39 | 2.9 ± 0.34 a | 0.56 ± 0.09 a,* | 1.72 ± 0.23 | 4.04 ± 0.64 a | 1.05 ± 0.15 a,* |
| Total Cer | 4.35 ± 0.61 | 6.94 ± 0.85 a | 2.84 ± 0.35 a,* | 6.83 ± 0.80 | 12.71 ± 1.0 a | 4.50 ± 0.52 a,* |
| Visceral Fat Tissue | Subcutaneous Fat Tissue | |||||
|---|---|---|---|---|---|---|
| Diacylglycerol | Control | HFD | HFD/Myr | Control | HFD | HFD/Myr |
| 16/16 | 0.41 ± 0.07 | 1.09 ± 0.18 | 0.6 ± 0.06 * | 1.07 ± 0.27 | 3.45 ± 0.64 | 3.07 ± 0.43 * |
| 16/18:1 | 1.8 ± 0.23 | 7.1 ± 1.0 | 5.3 ± 0.67 a,* | 5.20 ± 0.78 | 19.4 ± 3.2 a | 11.0 ± 0.7 a,* |
| 16/18:2 | 1.08 ± 0.19 | 3.09 ± 0.45 a | 2.13 ± 0.22 * | 2.0 ± 0.5 | 6.33 ± 0.97 a | 4.05 ± 0.33 a,* |
| 16/18 | 0.28 ± 0.05 | 1.09 ± 0.18 a | 0.55 ± 0.08 * | 0.93 ± 0.11 | 3.1 ± 0.68 a | 2.14 ± 0.16 a,* |
| 18:1/18:1 | 1.14 ± 0.25 | 6.0 ± 1.02 a | 3.19 ± 0.43 * | 3.39 ± 0.39 | 10.9 ± 1.3 a | 6.08 ± 0.30 * |
| 18:1/18:2 | 1.1 ± 0.20 | 5.32 ± 0.85 a | 2.52 ± 0.27 * | 2.18 ± 0.63 | 14.9 ± 2.1 a | 6.92 ± 0.44 a,* |
| 18/18:1 | 0.01 ± 0.004 | 0.05 ± 0.008 a | 0.027 ± 0.005 a,* | 0.08 ± 0.008 | 0.35 ± 0.049 a | 0.12 ± 0.02 a,* |
| Total DAG | 5.84 ± 0.47 | 23.73 ± 1.35 a | 14.32 ± 1.1 a,* | 14.88 ± 1.31 | 58.57 ± 4.28 a | 33.46 ± 1.46 a,* |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blachnio-Zabielska, A.U.; Razak Hady, H.; Markowski, A.R.; Kurianiuk, A.; Karwowska, A.; Górski, J.; Zabielski, P. Inhibition of Ceramide De Novo Synthesis Affects Adipocytokine Secretion and Improves Systemic and Adipose Tissue Insulin Sensitivity. Int. J. Mol. Sci. 2018, 19, 3995. https://doi.org/10.3390/ijms19123995
Blachnio-Zabielska AU, Razak Hady H, Markowski AR, Kurianiuk A, Karwowska A, Górski J, Zabielski P. Inhibition of Ceramide De Novo Synthesis Affects Adipocytokine Secretion and Improves Systemic and Adipose Tissue Insulin Sensitivity. International Journal of Molecular Sciences. 2018; 19(12):3995. https://doi.org/10.3390/ijms19123995
Chicago/Turabian StyleBlachnio-Zabielska, Agnieszka U., Hady Razak Hady, Adam R. Markowski, Adam Kurianiuk, Alicja Karwowska, Jan Górski, and Piotr Zabielski. 2018. "Inhibition of Ceramide De Novo Synthesis Affects Adipocytokine Secretion and Improves Systemic and Adipose Tissue Insulin Sensitivity" International Journal of Molecular Sciences 19, no. 12: 3995. https://doi.org/10.3390/ijms19123995
APA StyleBlachnio-Zabielska, A. U., Razak Hady, H., Markowski, A. R., Kurianiuk, A., Karwowska, A., Górski, J., & Zabielski, P. (2018). Inhibition of Ceramide De Novo Synthesis Affects Adipocytokine Secretion and Improves Systemic and Adipose Tissue Insulin Sensitivity. International Journal of Molecular Sciences, 19(12), 3995. https://doi.org/10.3390/ijms19123995