Transcriptome Sequence Analysis Elaborates a Complex Defensive Mechanism of Grapevine (Vitis vinifera L.) in Response to Salt Stress

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Global Transcriptome Sequence Analysis
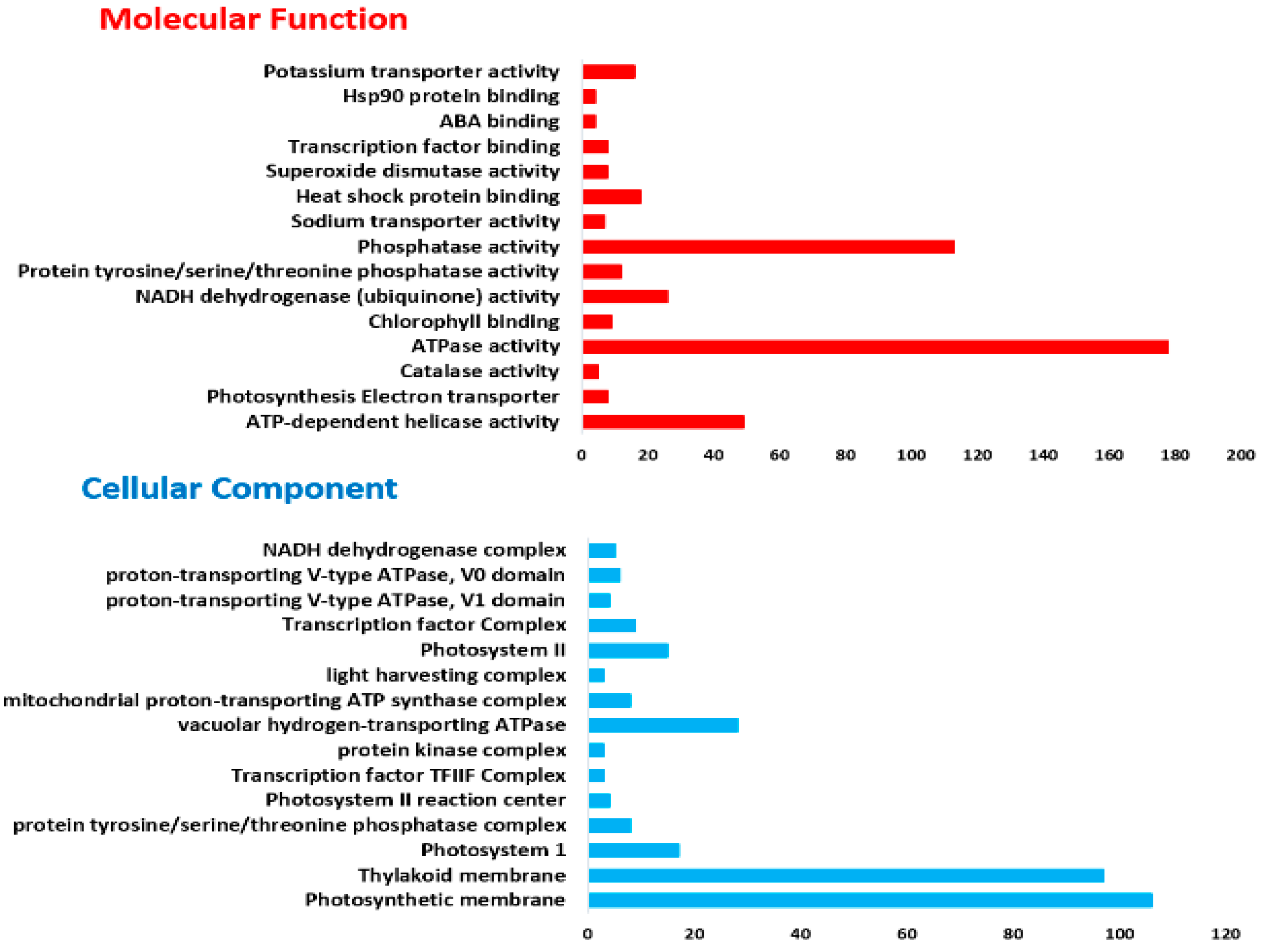
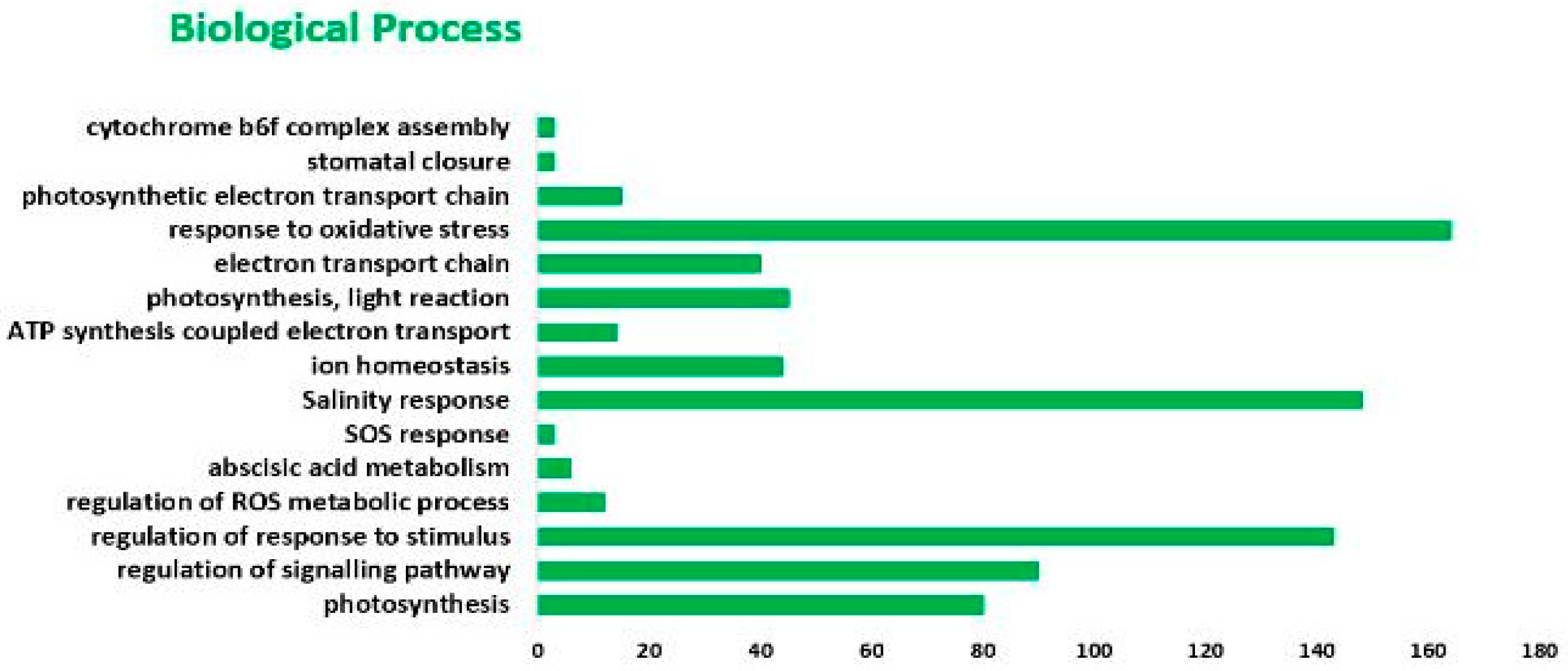
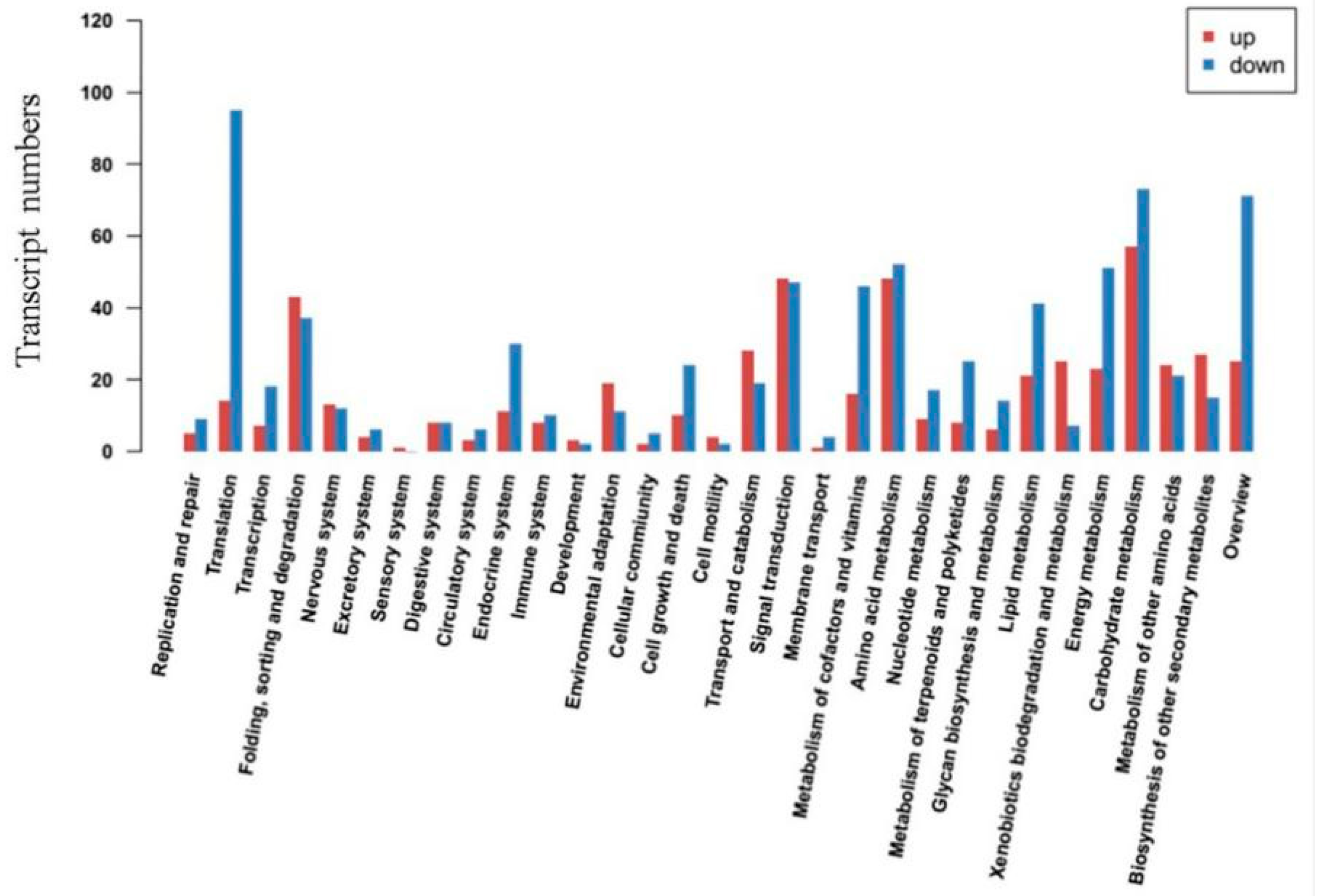
2.2. GO and KEGG Analysis of DEGs in Response to Salt Stress
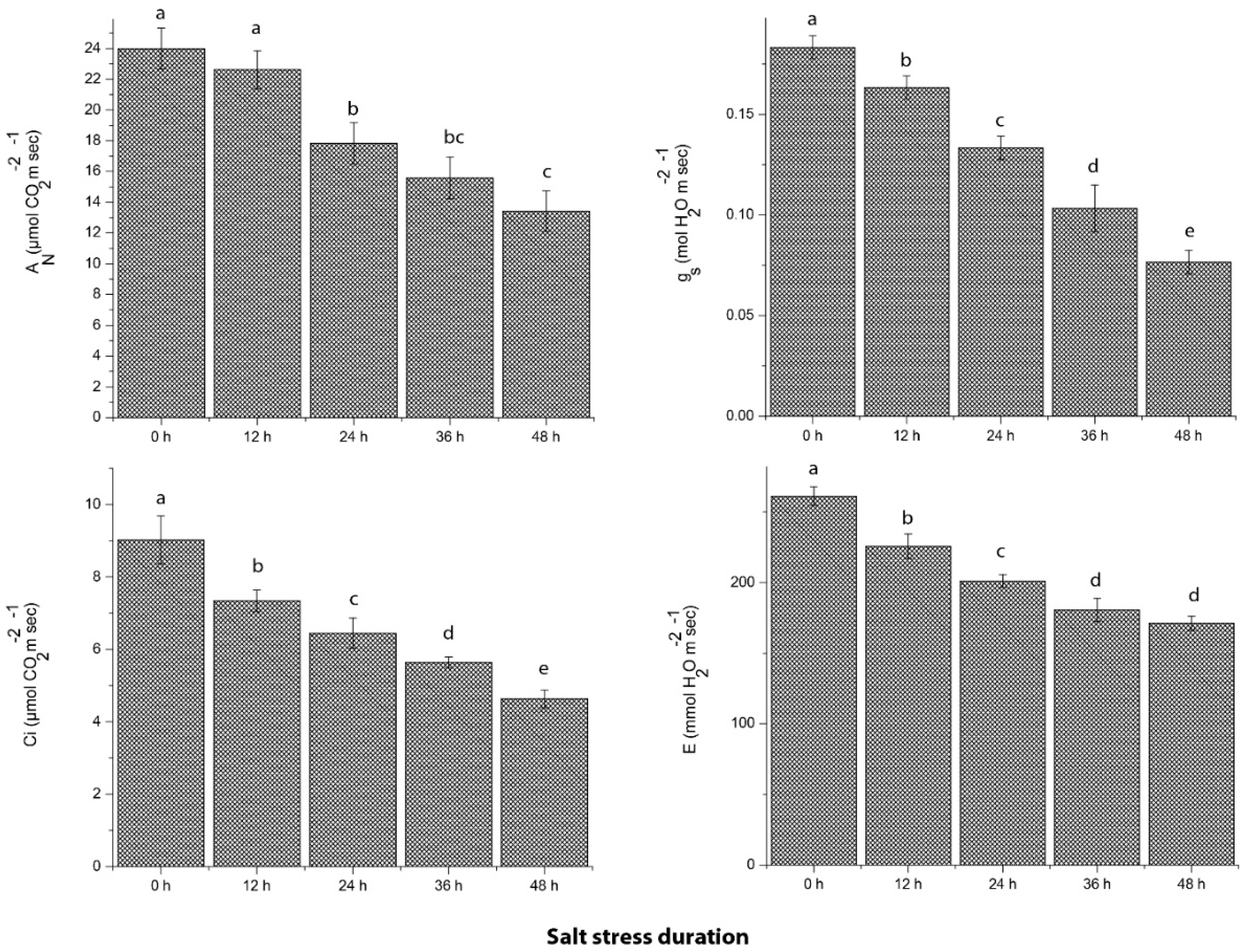
2.3. Photosynthetic Efficiency of Grapevine in Response to Salt Stress
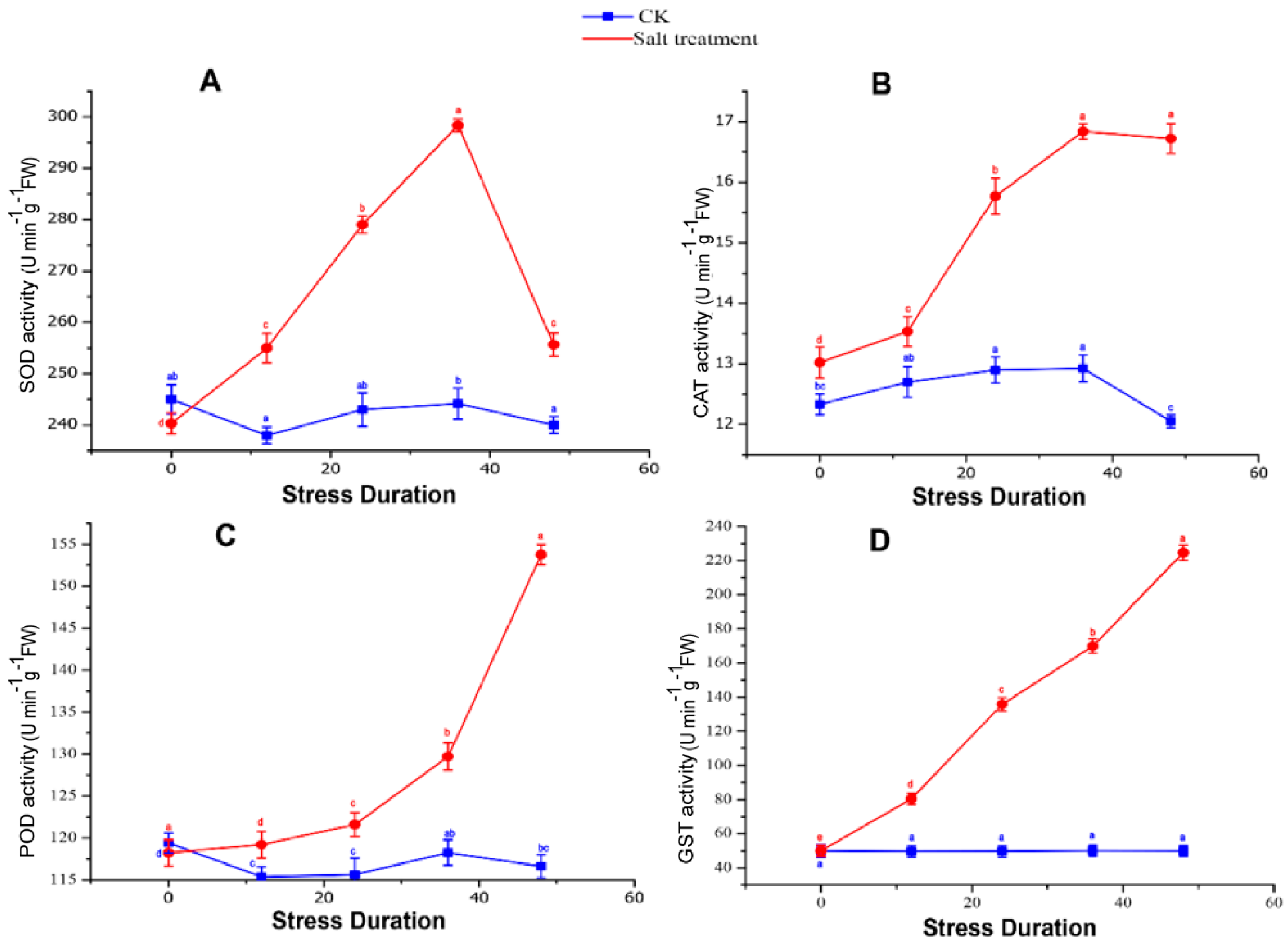
2.4. Production and Scavenging of Reactive Oxygen Species (ROS) in Response to Salt Stress
2.5. Heat Shock Protein (HSP) and Pathogenesis-Related Proteins (PR) in Response to Salt Stress
2.6. Regulation of Hormonal Signaling in Response to Salt Stress
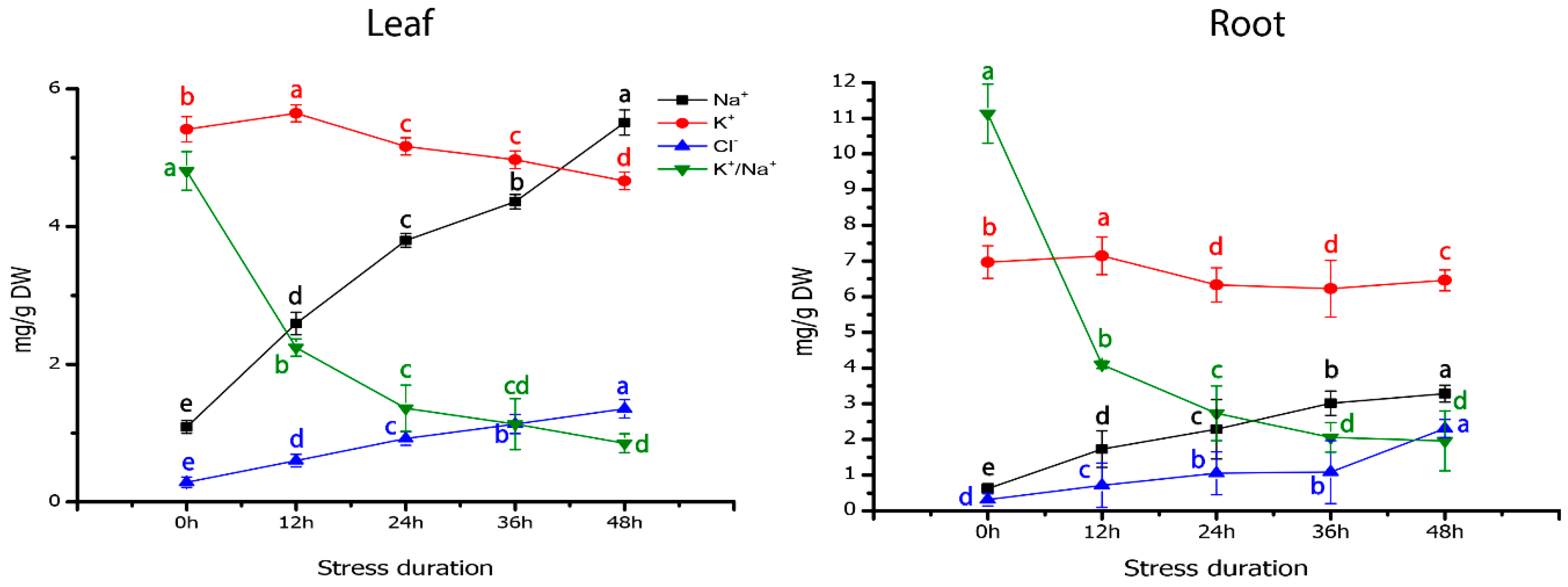
2.7. Ion Transport Systems Mediating Na+ Homeostasis in Response to Salt Stress
2.8. Transcription Factors in Response to Salt Stress
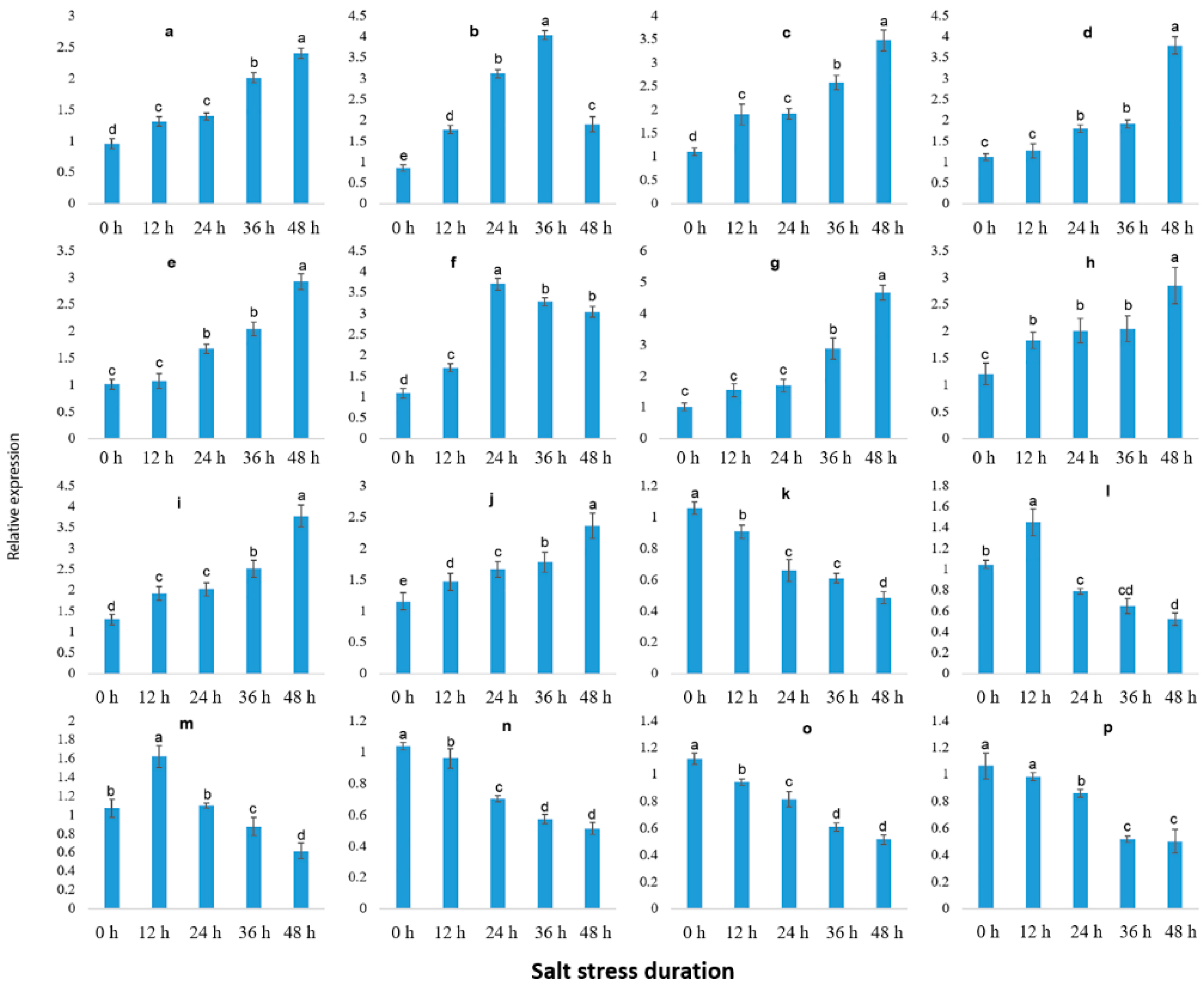
2.9. qRT-PCR Validation of Illumina RNA-Seq Results
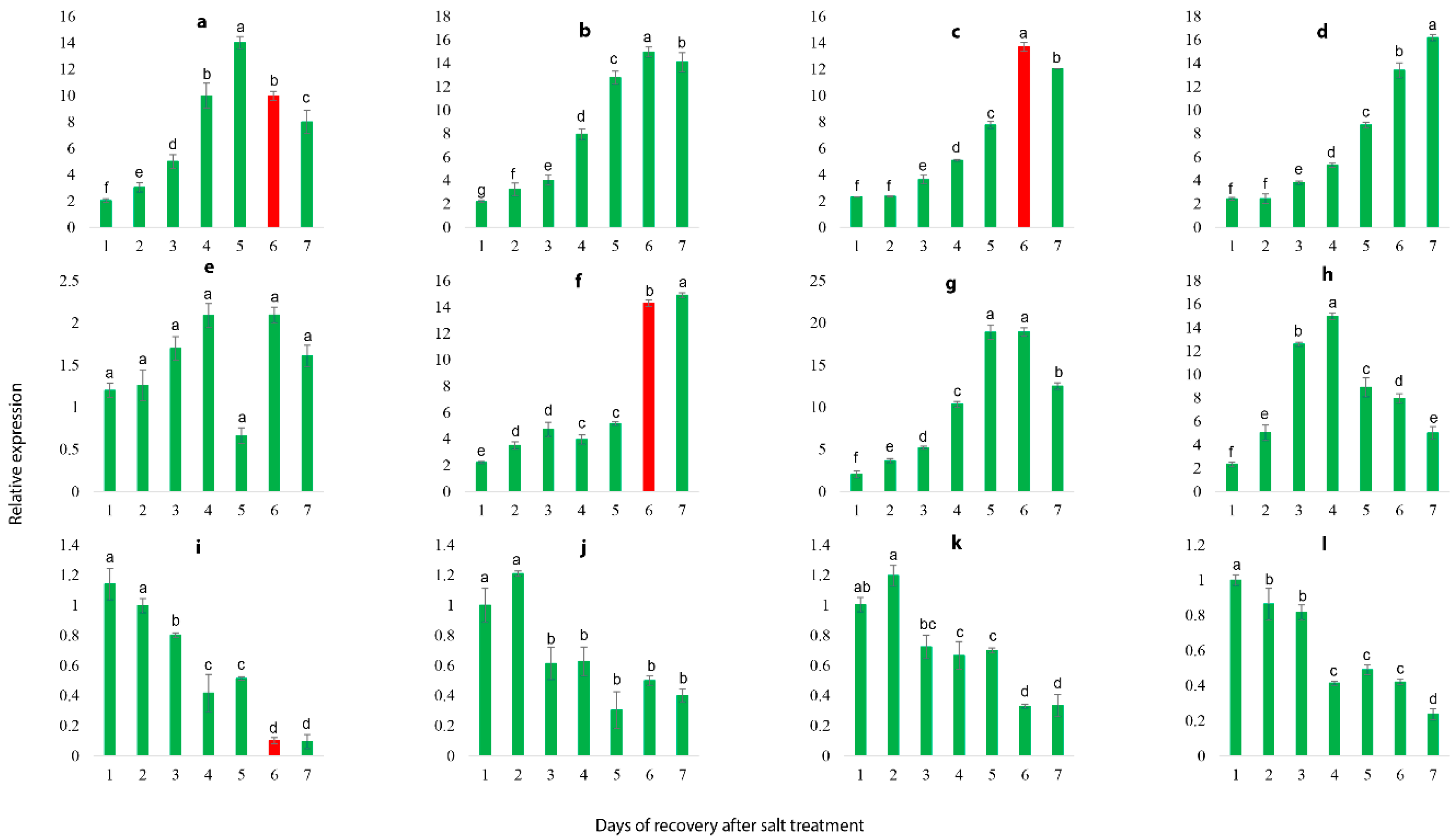
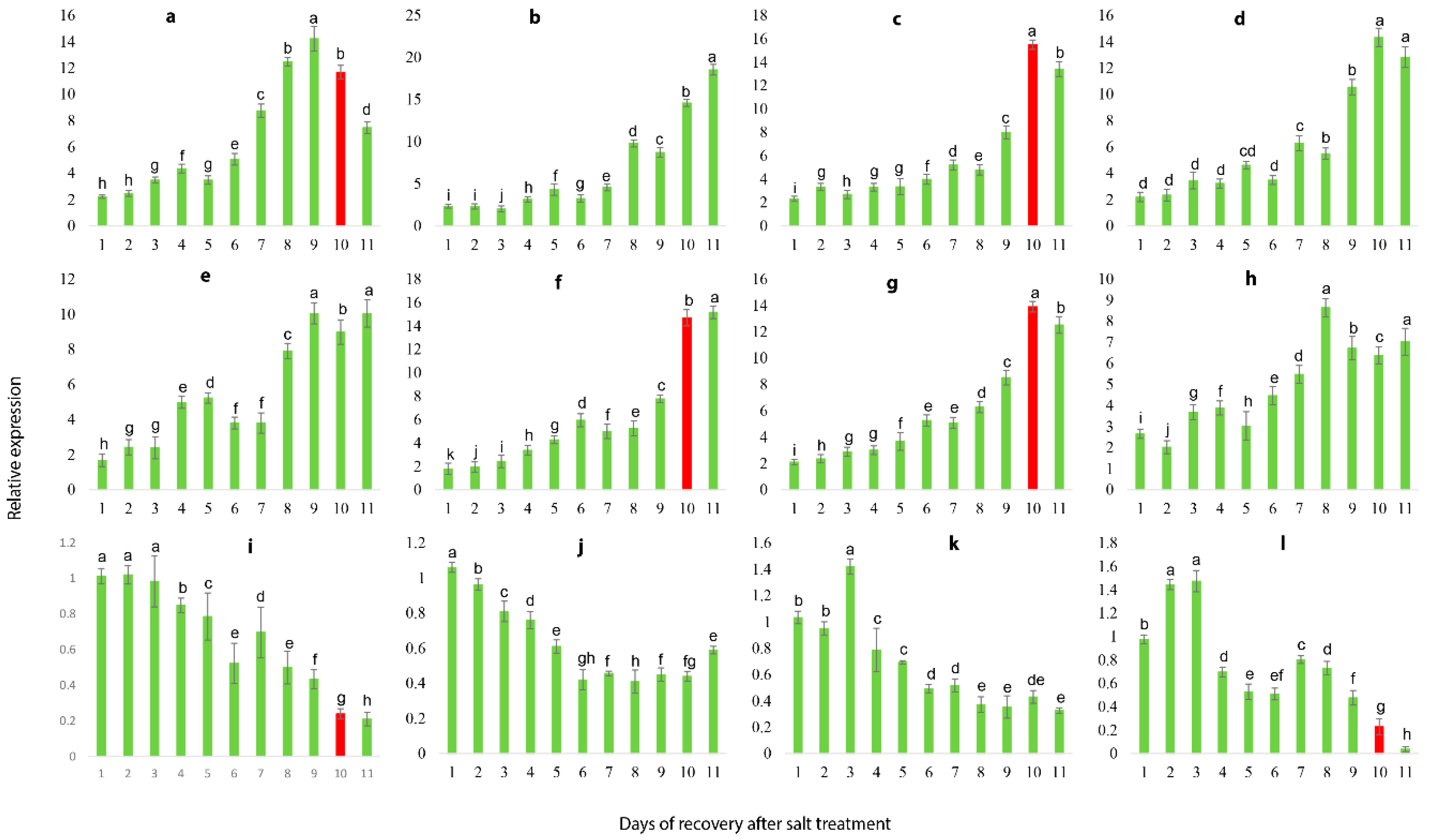
2.10. Salt Stress Recovery and the Selection and Validation of Marker Genes
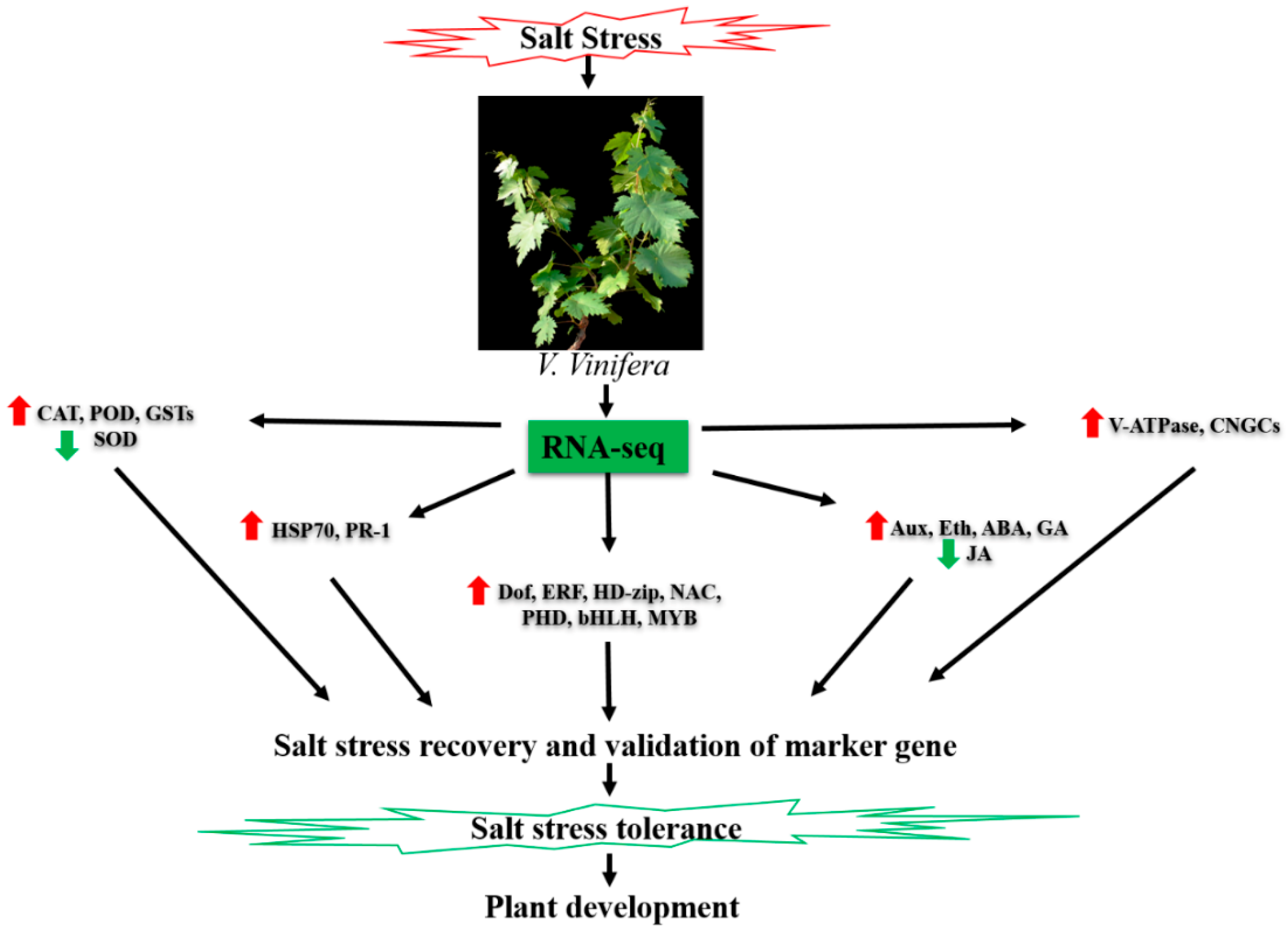
3. Discussion
4. Materials and Methods
4.1. Plant Material and Salt Treatments
4.2. RNA Extraction, cDNA Library Construction and Illumina Deep Sequencing
4.3. Mapping of reads, Gene Annotation and Analysis of Gene Expression Level
4.4. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomics (KEGG)
4.5. Estimation of Photosynthesis Rate and Determination of Several Enzymatic and Ionic Concentrations
4.6. Quantitative Real-Time PCR (qRT-PCR) Analysis of DEGs and Validation of Illumina RNA-Seq Results
4.7. Salt Stress and Recovery Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CAT | Catalase |
| Cl− | Chloride |
| DEGs | Differentially expressed genes |
| GO | Gene Ontology |
| GST | Glutathione S transferase |
| HSPs | Heat shock proteins |
| K+ | Potassium |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| Na+ | Sodium |
| POD | Peroxidase |
| PRs | Pathogenesis-related proteins |
| qRT-PCR | Quantitative reverse transcriptome-PCR |
| ROS | Reactive oxygen species |
| RNA-seq | RNA-sequencing |
| SOD | Superoxide dismutase |
| SS | Salt stress |
| TFs | Transcription factors |
References
- Haider, M.S.; Kurjogi, M.M.; Khalil-Ur-Rehman, M.; Fiaz, M.; Pervaiz, T.; Jiu, S.; Haifeng, J.; Chen, W.; Fang, J. Grapevine immune signaling network in response to drought stress as revealed by transcriptomic analysis. Plant Physiol. Biochem. 2017, 121, 187–195. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Sakamoto, A.; Nishiyama, Y.; Murata, N. Ionic and Osmotic Effects of Nacl-Induced Inactivation of Photosystems I and Ii in Synechococcus sp. Plant Physiol. 2000, 123, 1047–1056. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Jagendorf, A.; Zhu, J.K. Understanding and Improving Salt Tolerance in Plants. Crop Sci. 2005, 45, 437–448. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ Tolerance and Na+ Transport in Higher Plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Postnikova, O.A.; Shao, J.; Nemchinov, L.G. Analysis of the alfalfa root transcriptome in response to salinity stress. Plant Cell Physiol. 2013, 54, 1041–1055. [Google Scholar] [CrossRef]
- Fan, X.D.; Wang, J.Q.; Yang, N.; Dong, Y.Y.; Liu, L.; Wang, F.W.; Wang, N.; Chen, H.; Liu, W.C.; Sun, Y.P. Gene expression profiling of soybean leaves and roots under salt, saline-alkali and drought stress by high-throughput Illumina sequencing. Gene 2013, 512, 392–402. [Google Scholar] [CrossRef]
- Wang, B.; Lv, X.Q.; He, L.; Zhao, Q.; Xu, M.S.; Zhang, L.; Jia, Y.; Zhang, F.; Liu, F.L.; Liu, Q.L. Whole-transcriptome sequence analysis of verbena bonariensis in response to drought stress. Int. J. Mol. Sci. 2018, 19, 1751. [Google Scholar] [CrossRef] [PubMed]
- Jaffar, M.A.; Song, A.; Faheem, M.; Chen, S.; Jiang, J.; Chen, L.; Fan, Q.; Chen, F. Involvement of cmwrky10 in drought tolerance of chrysanthemum through the aba-signaling pathway. Int. J. Mol. Sci. 2016, 17, 693. [Google Scholar] [CrossRef]
- Wang, C.; Chen, H.F.; Hao, Q.N.; Shan, Z.H.; Zhou, R.; Zhi, H.J.; Zhou, X.A. Transcript profile of the response of two soybean genotypes to potassium deficiency. PLoS ONE 2012, 7, e39856. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tao, X.; Tang, X.M.; Xiao, L.; Sun, J.L.; Yan, X.F.; Li, D.; Deng, H.Y.; Ma, X.R. Comparative transcriptome analysis of tomato (Solanum lycopersicum) in response to exogenous abscisic acid. BMC Genom. 2013, 14, 841. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.A.; Corpas, F.J.; Gómez, M.; Río, L.A.D.; Sevilla, F. Salt-induced oxidative stress mediated by activated oxygen species in pea leaf mitochondria. Physiol. Plant. 2010, 89, 103–110. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Grimm, B.; Wobus, U.; Weschke, W. Differential response of antioxidant compounds to salinity stress in salt-tolerant and salt-sensitive seedlings of foxtail millet (Setaria italica). Physiol. Plant. 2010, 109, 435–442. [Google Scholar] [CrossRef]
- Chaparzadeh, N.; D’Amico, M.L.; Khavari-Nejad, R.A.; Izzo, R.; Navari-Izzo, F. Antioxidative responses of Calendula officinalis under salinity conditions. Plant Physiol. Biochem. 2004, 42, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Marrs, K.A. The functions and regulation of glutathione-S-transferses in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 127–158. [Google Scholar] [CrossRef]
- Edwards, R.; Dixon, D.P.; Walbot, V. Plant glutathione S-transferases: Enzymes with multiple functions in sickness and in health. Trends Plant Sci. 2000, 5, 193–198. [Google Scholar] [CrossRef]
- Jakab, G.; Ton, J.; Flors, V.; Zimmerli, L.; Métraux, J.P.; Mauchmani, B. Enhancing Arabidopsis salt and drought stress tolerance by chemical priming for its abscisic acid responses. Plant Physiol. 2005, 139, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Son, T.; Park, S.; Lee, I.; Lee, B.; Kim, H.; Lee, S. Influences of gibberellin and auxin on endogenous plant hormone and starch mobilization during rice seed germination under salt stress. J. Environ. Biol. 2006, 27, 181. [Google Scholar]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Annunziata, M.G.; Pontecorvo, G.; Fuggi, A.; Woodrow, P. Salinity stress and salt tolerance. In Abiotic Stress in Plants-Mechanisms and Adaptations; InTech: Rijeka, Croatia, 2011. [Google Scholar]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes. An overview with the central focus on rice plants. Rice 2012, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.T.; Zhang, Z.S.; Gao, H.Y.; Xue, Z.C.; Yang, C.; Meng, X.L.; Meng, Q.W. Mitochondrial alternative oxidase pathway protects plants against photoinhibition by alleviating inhibition of the repair of photodamaged PSII through preventing formation of reactive oxygen species in Rumex, K.-1 leaves. Physiol. Plant. 2011, 143, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.S.; Kurjogi, M.M.; Khalil-ur-Rehman, M.; Pervez, T.; Songtao, J.; Fiaz, M.; Jogaiah, S.; Wang, C.; Fang, J. Drought stress revealed physiological, biochemical and gene-expressional variations in “Yoshihime”peach (Prunus Persica, L.) cultivar. J. Plant Interact. 2018, 13, 83–90. [Google Scholar] [CrossRef]
- Wang, N.; Qian, Z.; Luo, M.; Fan, S.; Zhang, X.; Zhang, L. Identification of Salt Stress Responding Genes Using Transcriptome Analysis in Green Alga Chlamydomonas reinhardtii. Int. J. Mol. Sci. 2018, 19, 3359. [Google Scholar] [CrossRef] [PubMed]
- Hiroaki, F.; Verslues, P.E.; Jian-Kang, Z. Arabidopsis decuple mutant reveals the importance of SnRK2 kinases in osmotic stress responses in vivo. Proc. Natl. Acad. Sci. USA 2011, 108, 1717–1722. [Google Scholar] [Green Version]
- Aydi, S.; Sassi, S.; Abdelly, C. Growth, nitrogen fixation and ion distribution in Medicago truncatula subjected to salt stress. Plant Soil 2008, 312, 59. [Google Scholar] [CrossRef]
- Teakle, N.; Flowers, T.; Real, D.; Colmer, T. Lotus tenuis tolerates the interactive effects of salinity and waterlogging by “excluding”Na+ and Cl− from the xylem. J. Exp. Bot. 2007, 58, 2169–2180. [Google Scholar] [CrossRef]
- Tanveer, M.; Shabala, S. Targeting Redox Regulatory Mechanisms for Salinity Stress Tolerance in Crops. In Salinity Responses and Tolerance in Plants; Springer: Basel, Switzerland, 2018; Volume 1, pp. 213–234. [Google Scholar]
- Mandhania, S.; Madan, S.; Sawhney, V. Antioxidant defense mechanism under salt stress in wheat seedlings. Biol. Plant. 2006, 50, 227–231. [Google Scholar] [CrossRef]
- Abogadallah, G.M. Insights into the significance of antioxidative defense under salt stress. Plant Signal. Behav. 2010, 5, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Liu, Y.; Rao, J.; Wang, G.; Li, H.; Ge, F.; Chen, C. Overexpression of the glutathione S-transferase gene from Pyrus pyrifolia fruit improves tolerance to abiotic stress in transgenic tobacco plants. Mol. Biol. 2013, 47, 515–523. [Google Scholar] [CrossRef]
- Wu, Q.; Bai, X.; Zhao, W.; Xiang, D.; Wan, Y.; Yan, J.; Zou, L.; Zhao, G. De novo assembly and analysis of tartary buckwheat (fagopyrum tataricum Garetn.) transcriptome discloses key regulators involved in salt-stress response. Genes 2017, 8, 255. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhou, L.; Yan, P.; Xiaojuan, W.; Dandan, P.; Yaping, L.; Xiaoshuang, H.; Xinquan, Z.; Xiao, M.; Linkai, H. Clones of FeSOD, MDHAR, DHAR Genes from White Clover and Gene Expression Analysis of ROS-Scavenging Enzymes during Abiotic Stress and Hormone Treatments. Molecules 2015, 20, 20939–20954. [Google Scholar] [Green Version]
- Baneh, H.D.; Attari, H.; Hassani, A.; Abdollahi, R. Salinity effects on the physiological parameters and oxidative enzymatic activities of four Iranian grapevines (Vitis vinifera L.) cultivar. Int. J. Agric. Crop Sci. 2013, 5, 1022. [Google Scholar]
- Weisany, W.; Sohrabi, Y.; Heidari, G.; Siosemardeh, A.; Ghassemi-Golezani, K. Changes in antioxidant enzymes activity and plant performance by salinity stress and zinc application in soybean (Glycine max L.). Plant Omics 2012, 5, 60. [Google Scholar]
- Nollen, E.A.; Morimoto, R.I. Chaperoning signaling pathways. molecular chaperones as stress-sensingheat shock’proteins. J. Cell Sci. 2002, 115, 2809–2816. [Google Scholar]
- Li, J.; He, Q.; Sun, H.; Liu, X. Acclimation-dependent expression of heat shock protein 70 in Pacific abalone (Haliotis discus hannai Ino) and its acute response to thermal exposure. Chin. J. Oceanol. Limnol. 2012, 30, 146–151. [Google Scholar] [CrossRef]
- Shao, F.; Zhang, L.; Wilson, I.; Qiu, D. Transcriptomic Analysis of Betula halophila in Response to Salt Stress. Int. J. Mol. Sci. 2018, 19, 3412. [Google Scholar] [CrossRef]
- Martin, G.B.; Bogdanove, A.J.; Sessa, G. Understanding the functions of plant disease resistance proteins. Annu. Rev. Plant Biol. 2003, 54, 23–61. [Google Scholar] [CrossRef]
- Christensen, A.B.; Cho, B.H.; Næsby, M.; Gregersen, P.L.; Brandt, J.; Madriz-Ordeñana, K.; Collinge, D.B.; Thordal-Christensen, H. The molecular characterization of two barley proteins establishes the novel PR-17 family of pathogenesis-related proteins. Mol. Plant Pathol. 2002, 3, 135–144. [Google Scholar] [CrossRef]
- Adams, P.; Thomas, J.C.; Vernon, D.M.; Bohnert, H.J.; Jensen, R.G. Distinct cellular and organismic responses to salt stress. Plant Cell Physiol. 1992, 33, 1215–1223. [Google Scholar]
- Brini, F.; Masmoudi, K. Ion transporters and abiotic stress tolerance in plants. ISRN Mol. Biol. 2012, 2012, 927436. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.; Sanish, S.; Iyengar, E. Compartmentation of ions and organic compounds in Salicornia brachiata Roxb. Biol. Plant 1993, 35, 547. [Google Scholar] [CrossRef]
- Zhu, J.-K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Rajendran, K.; Tester, M.; Roy, S.J. Quantifying the three main components of salinity tolerance in cereals. Plant Cell Environ. 2009, 32, 237–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huazhong, S.; Quintero, F.J.; Pardo, J.M.; Jian-Kang, Z. The putative plasma membrane Na(+)/H(+) antiporter SOS1 controls long-distance Na(+) transport in plants. Plant Cell 2002, 14, 465–477. [Google Scholar]
- Quan-Sheng, Q.; Yan, G.; Dietrich, M.A.; Schumaker, K.S.; Jian-Kang, Z. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 2002, 99, 8436–8441. [Google Scholar] [Green Version]
- Sottosanto, J.B.; Saranga, Y.; Blumwald, E. Impact of AtNHX1, a vacuolar Na + /H. + antiporter, upon gene expression during short- and long-term salt stress in Arabidopsis thaliana. Bmc Plant Biol. 2007, 7, 18. [Google Scholar] [CrossRef]
- Zhu, J.K.; Shi, J.; Singh, U.; Wyatt, S.E.; Bressan, R.A.; Hasegawa, P.M.; Carpita, N.C. Enrichment of vitronectin-and fibronectin-like proteins in NaCl-adapted plant cells and evidence for their involvement in plasma membrane-cell wall adhesion. Plant J. 1993, 3, 637–646. [Google Scholar] [CrossRef]
- Tomoaki, H.; Jo, M.; Masahiro, K.; Hua, Y.; Kinya, Y.; Rie, H.; Wai-Yin, C.; Ho-Yin, L.; Kazumi, H.; Mami, K. Enhanced salt tolerance mediated by AtHKT1 transporter-induced Na unloading from xylem vessels to xylem parenchyma cells. Plant J. Cell Mol. Biol. 2010, 44, 928–938. [Google Scholar]
- Li, H.; Tang, X.; Zhu, J.; Yang, X.; Zhang, H. De Novo Transcriptome Characterization, Gene Expression Profiling and Ionic Responses of Nitraria sibirica Pall. under Salt Stress. Forests 2017, 8, 211. [Google Scholar] [CrossRef]
- Henderson, S.W.; Baumann, U.; Blackmore, D.H.; Walker, A.R.; Walker, R.R.; Gilliham, M. Shoot chloride exclusion and salt tolerance in grapevine is associated with differential ion transporter expression in roots. BMC Plant Biol. 2014, 14, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.I.; Wang, L.; Cao, J.; Bingjun, Y.U. Bioinformatics analysis of CLC homologous genes family in soybean genome. J. Nanjing Agric. Univ. 2014, 37, 35–43. [Google Scholar]
- Magnan, F.; Ranty, B.; Charpenteau, M.; Sotta, B.; Galaud, J.P.; Aldon, D. Mutations in AtCML9, a calmodulin-like protein from Arabidopsis thaliana, alter plant responses to abiotic stress and abscisic acid. Plant J. 2008, 56, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liang, X.; Wan, Q.; Wang, X.; Bi, Y. Ethylene and nitric oxide are involved in maintaining ion homeostasis in Arabidopsis callus under salt stress. Planta 2009, 230, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Lu, R.; Dai, Z.; Yan, A.; Tang, Q.; Cheng, C.; Xu, Y.; Yang, W.; Su, J. Salt-Stress Response Mechanisms Using de Novo Transcriptome Sequencing of Salt-Tolerant and Sensitive Corchorus spp. Genotypes. Genes 2017, 8, 226. [Google Scholar] [CrossRef]
- Haider, M.S.; Zhang, C.; Kurjogi, M.M.; Pervaiz, T.; Zheng, T.; Zhang, C.; Lide, C.; Shangguan, L.; Fang, J. Insights into grapevine defense response against drought as revealed by biochemical, physiological and RNA-Seq analysis. Sci. Rep. 2017, 7, 13134. [Google Scholar] [CrossRef] [Green Version]
- Pedranzani, H.; Racagni, G.; Alemano, S.; Miersch, O.; Ramírez, I.; Peña-Cortés, H.; Taleisnik, E.; Machado-Domenech, E.; Abdala, G. Salt tolerant tomato plants show increased levels of jasmonic acid. Plant Growth Regul. 2003, 41, 149–158. [Google Scholar] [CrossRef]
- Nagaoka, S.; Takano, T. Salt tolerance-related protein STO binds to a Myb transcription factor homologue and confers salt tolerance in Arabidopsis. J. Exp. Bot. 2003, 54, 2231–2237. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Dai, X.; Zhang, W.-H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold and dehydration tolerance in rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef]
- Rahaie, M.; Xue, G.-P.; Naghavi, M.R.; Alizadeh, H.; Schenk, P.M. A MYB gene from wheat (Triticum aestivum L.) is up-regulated during salt and drought stresses and differentially regulated between salt-tolerant and sensitive genotypes. Plant Cell Rep. 2010, 29, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought and salt stress in transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Li, J.; Liu, P.; Duan, J.; Zhao, Y.; Guo, X.; Li, Y.; Zhang, H.; Ali, J.; Li, Z. Overexpression of OsMYB48-1, a novel MYB-related transcription factor, enhances drought and salinity tolerance in rice. PLoS ONE 2014, 9, e92913. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gao, Y.; Xu, H.; Dai, Y.; Deng, D.; Chen, J. ZmWRKY33: A WRKY maize transcription factor conferring enhanced salt stress tolerances in Arabidopsis. Plant Growth Regul. 2013, 70, 207–216. [Google Scholar] [CrossRef]
- Shi, W.; Liu, D.; Hao, L.; Wu, C.A.; Guo, X.; Li, H. GhWRKY39, a member of the WRKY transcription factor family in cotton, has a positive role in disease resistance and salt stress tolerance. Plant Cell Tissue Organ Cult. 2014, 118, 17–32. [Google Scholar] [CrossRef]
- Wei, W.; Huang, J.; Hao, Y.J.; Zou, H.F.; Wang, H.W.; Zhao, J.Y.; Liu, X.Y.; Zhang, W.K.; Ma, B.; Zhang, J.S. Soybean GmPHD-Type Transcription Regulators Improve Stress Tolerance in Transgenic Arabidopsis Plants. PLoS ONE 2009, 4, e7209. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Bressan, R.A.; Carpita, N.C. Cell Walls of Tobacco Cells and Changes in Composition Associated with Reduced Growth upon Adaptation to Water and Saline Stress. Plant Physiol. 1989, 91, 48–53. [Google Scholar] [Green Version]
- Cameron, K.D.; Teece, M.A.; Smart, L.B. Increased accumulation of cuticular wax and expression of lipid transfer protein in response to periodic drying events in leaves of tree tobacco. Plant Physiol. 2006, 140, 176–183. [Google Scholar]
- Gothandam, K.M.; Nalini, E.; Karthikeyan, S.; Jeongsheop, S. OsPRP3, a flower specific proline-rich protein of rice, determines extracellular matrix structure of floral organs and its overexpression confers cold-tolerance. Plant Mol. Biol. 2010, 72, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Kampkötter, A.; Volkmann, T.E.; de Castro, S.H.; Leiers, B.; Klotz, L.O.; Johnson, T.E.; Link, C.D.; Henkle-Dührsen, K. Functional analysis of the glutathione S-transferase 3 from Onchocerca volvulus (Ov-GST-3): A parasite GST confers increased resistance to oxidative stress in Caenorhabditis elegans. J. Mol. Biol. 2003, 325, 25–37. [Google Scholar] [CrossRef]
- Pervaiz, T.; Haifeng, J.; Salman, H.M.; Cheng, Z.; Cui, M.; Wang, M.; Cui, L.; Wang, X.; Fang, J. Transcriptomic Analysis of Grapevine (cv. Summer Black) Leaf, Using the Illumina Platform. PLoS ONE 2016, 11, e0147369. [Google Scholar]
- Haider, M.S.; Khan, I.A.; Naqvi, S.A.; Jaskani, M.J.; Khan, R.W.; Nafees, M.; Pasha, I. Fruit developmental stages effects on biochemical attributes in date palm. Pak. J. Agric. Sci. 2013, 50, 577–583. [Google Scholar]
- Haider, M.S.; Khan, I.A.; Jaskani, M.J.; Naqvi, S.A.; Khan, M.M. Biochemical attributes of dates at three maturation stages. Emir. J. Food Agric. 2014, 11, 953–962. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, J.; Zhong, Y.; Geng, F.; Cramer, G.R.; Cheng, Z.M. Subfunctionalization of cation/proton antiporter 1 genes in grapevine in response to salt stress in different organs. Hortic. Res. 2015, 2, 15031. [Google Scholar] [CrossRef] [PubMed] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait Name | Description | No. of Up-Regulated | No. of Down-Regulated | Sum |
|---|---|---|---|---|
| ROS scavenging | Fe-SOD | 0 | 1 | 1 |
| POD | 8 | 0 | 8 | |
| CAT | 2 | 0 | 2 | |
| GSH-AsA cycle | MDAR | 1 | 0 | 1 |
| APx | 1 | 0 | 1 | |
| GR | 0 | 2 | 2 | |
| Grx | 1 | 1 | 2 | |
| GPX pathway | GST | 8 | 8 | 16 |
| Prx/Trx | Trx | 4 | 5 | 9 |
| Cyanide-resistant respiration | AOX | 0 | 1 | 1 |
| Copper-containing enzymes | PPO | 0 | 1 | 1 |
| Trait Name | Description | No. of Up-Regulated | No. of Down-Regulated | Sum |
|---|---|---|---|---|
| Heat shock proteins | HMW HSPs | 1 | 3 | 4 |
| LMW HSPs | 16 | 1 | 17 | |
| small HSPs | 12 | 6 | 18 | |
| other HSPs | 7 | 5 | 12 | |
| heat-stress transcription factors | 3 | 3 | 6 | |
| PR-1 | pathogenesis-related protein 1 | 4 | 0 | 4 |
| PR-2 | β-1,3-glucanase | 0 | 1 | 1 |
| PR-3,4,8,11 | chitinase | 0 | 2 | 2 |
| PR-5 | Thaumatin-like protein | 1 | 5 | 6 |
| PR-14 | lipid transfer protein | 7 | 1 | 8 |
| PR-15 | germin-like protein | 0 | 1 | 1 |
| Disease resistance proteins | 9 | 4 | 13 | |
| snakin | 2 | 0 | 2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, L.; Haider, M.S.; Khan, N.; Nasim, M.; Jiu, S.; Fiaz, M.; Zhu, X.; Zhang, K.; Fang, J. Transcriptome Sequence Analysis Elaborates a Complex Defensive Mechanism of Grapevine (Vitis vinifera L.) in Response to Salt Stress. Int. J. Mol. Sci. 2018, 19, 4019. https://doi.org/10.3390/ijms19124019
Guan L, Haider MS, Khan N, Nasim M, Jiu S, Fiaz M, Zhu X, Zhang K, Fang J. Transcriptome Sequence Analysis Elaborates a Complex Defensive Mechanism of Grapevine (Vitis vinifera L.) in Response to Salt Stress. International Journal of Molecular Sciences. 2018; 19(12):4019. https://doi.org/10.3390/ijms19124019
Chicago/Turabian StyleGuan, Le, Muhammad Salman Haider, Nadeem Khan, Maazullah Nasim, Songtao Jiu, Muhammad Fiaz, Xudong Zhu, Kekun Zhang, and Jinggui Fang. 2018. "Transcriptome Sequence Analysis Elaborates a Complex Defensive Mechanism of Grapevine (Vitis vinifera L.) in Response to Salt Stress" International Journal of Molecular Sciences 19, no. 12: 4019. https://doi.org/10.3390/ijms19124019
APA StyleGuan, L., Haider, M. S., Khan, N., Nasim, M., Jiu, S., Fiaz, M., Zhu, X., Zhang, K., & Fang, J. (2018). Transcriptome Sequence Analysis Elaborates a Complex Defensive Mechanism of Grapevine (Vitis vinifera L.) in Response to Salt Stress. International Journal of Molecular Sciences, 19(12), 4019. https://doi.org/10.3390/ijms19124019