Resistance to Spot Blotch in Two Mapping Populations of Common Wheat Is Controlled by Multiple QTL of Minor Effects

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
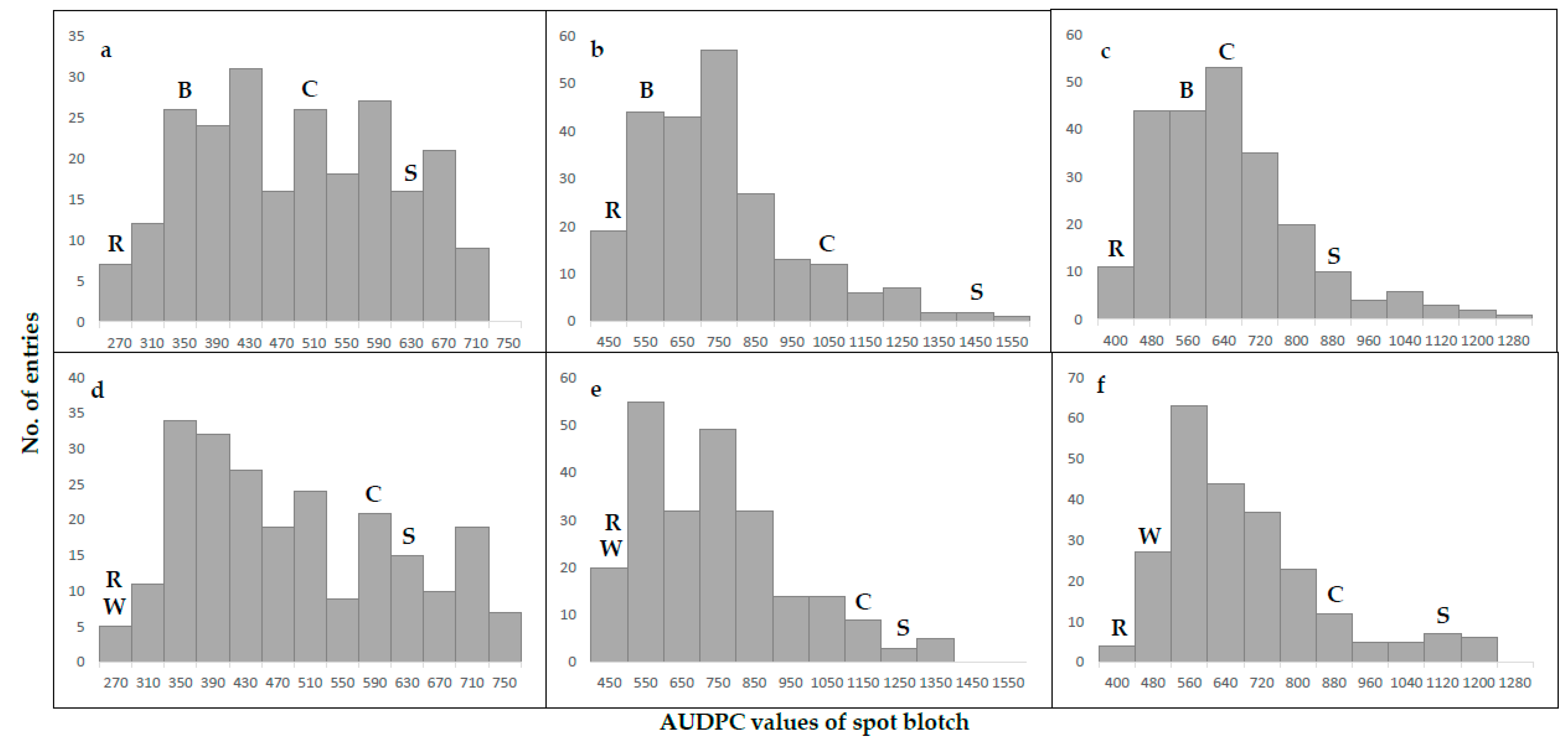
2.1. Phenotypic Analysis
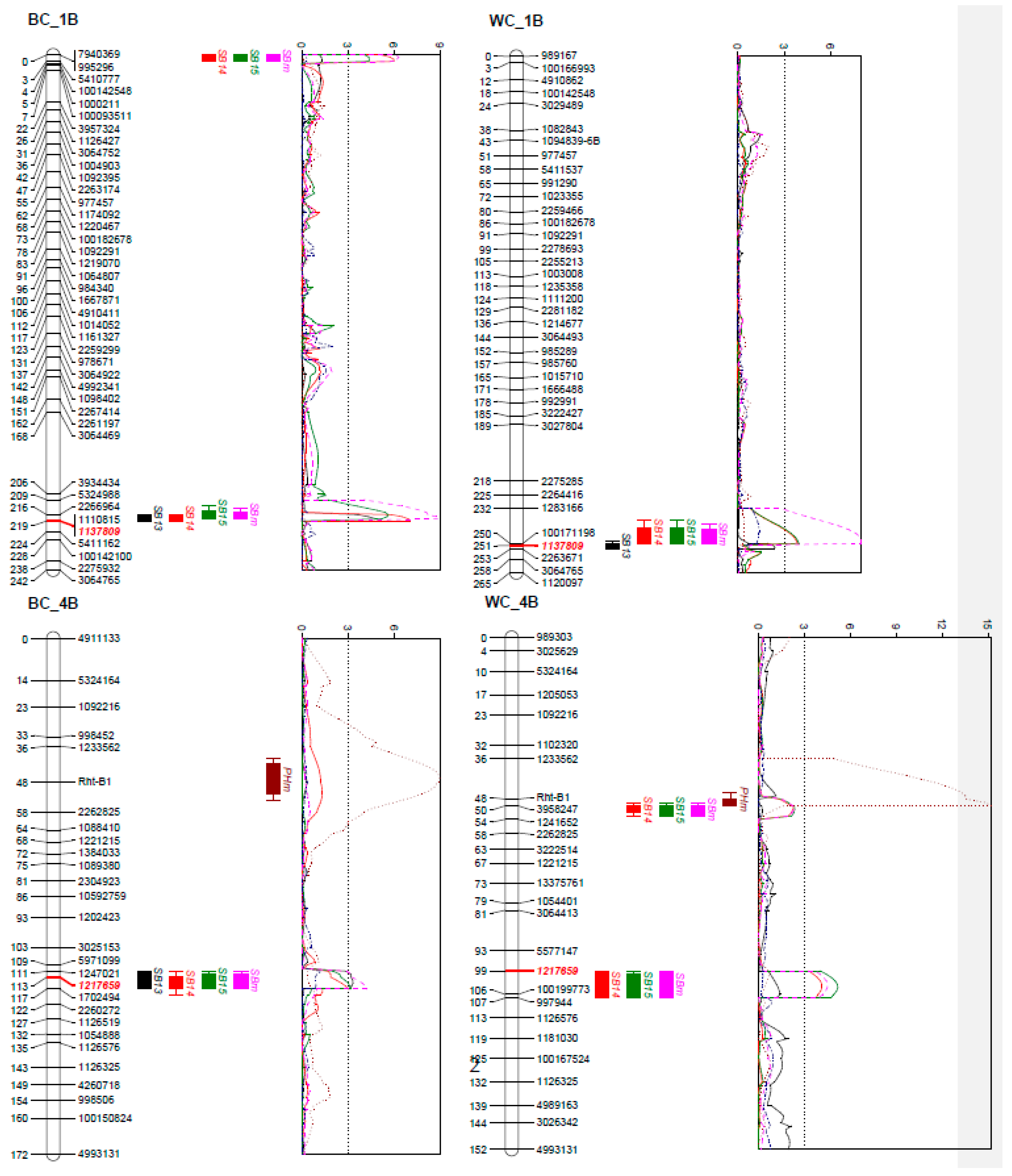
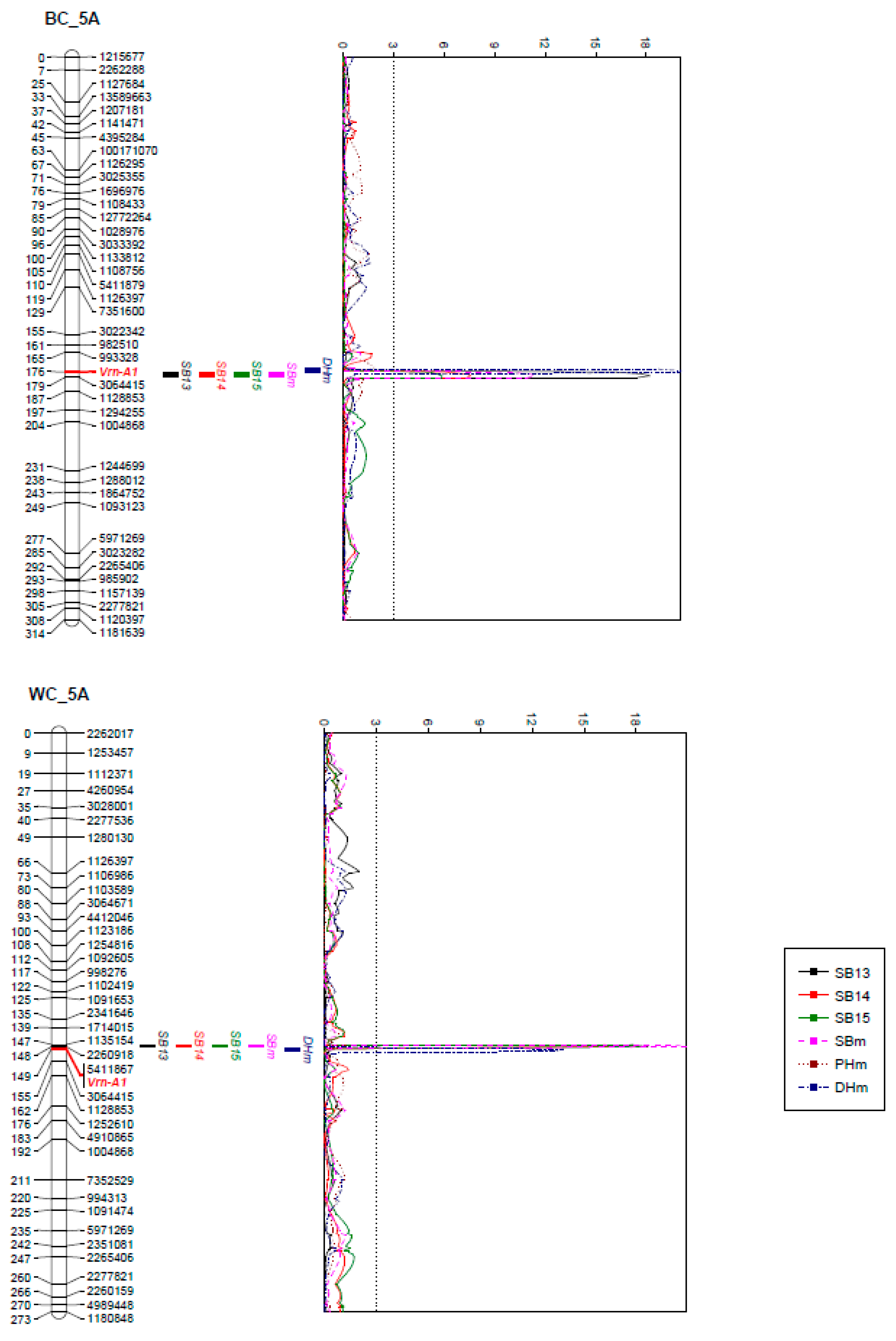
2.2. Genotyping and Linkage Analysis
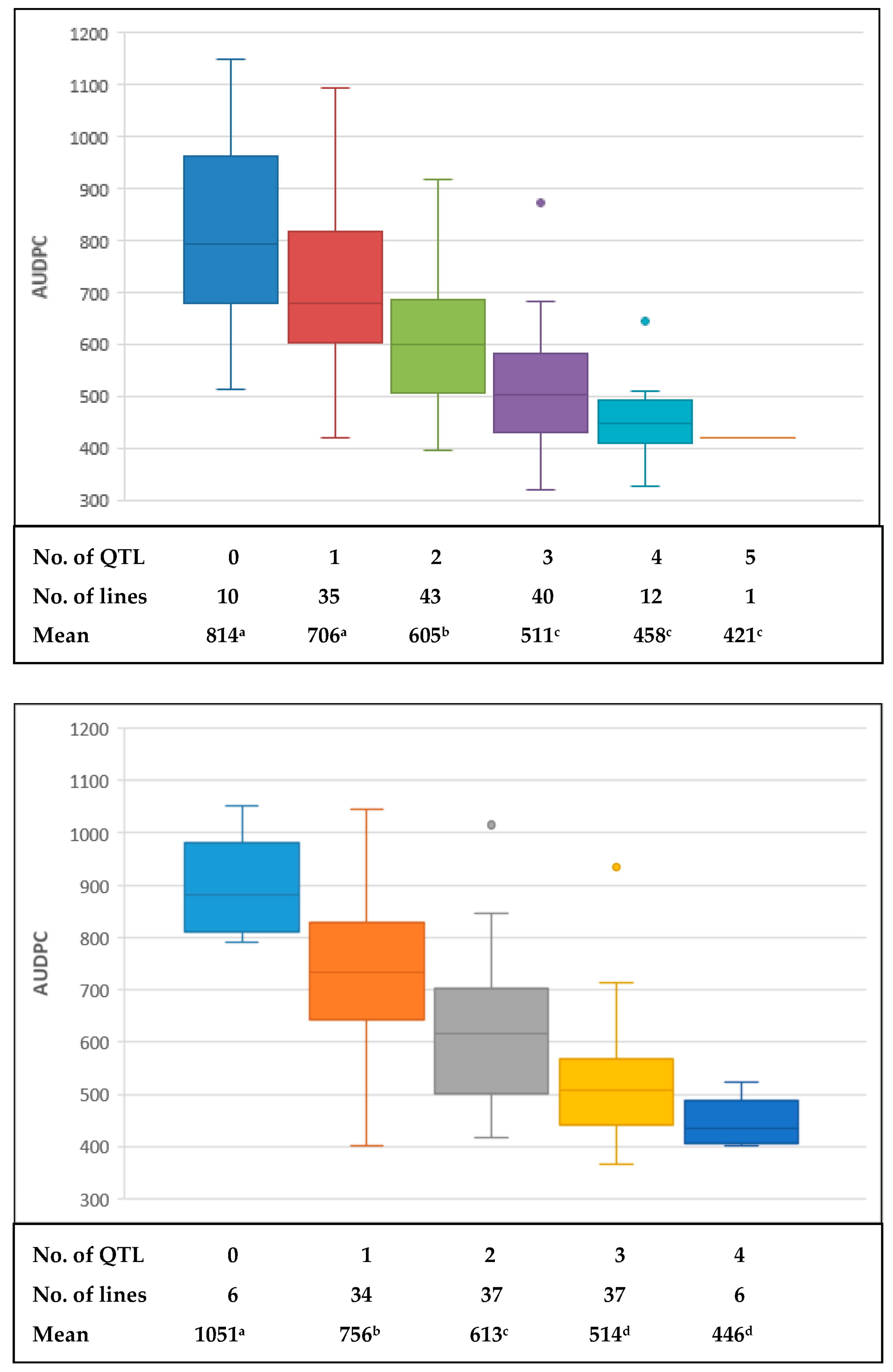
2.3. QTL Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Field Trials
4.2. Disease Screening Protocols
4.3. Statistical Analyses
4.4. Genotyping
4.5. Linkage and QTL Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gupta, P.K.; Chand, R.; Vasistha, N.K.; Pandey, S.P.; Kumar, U.; Mishra, V.K.; Joshi, A.K. Spot blotch disease of wheat: The current status of research on genetics and breeding. Plant Pathol. 2018, 67, 508–531. [Google Scholar] [CrossRef]
- Duveiller, E.; Sharma, R.C. Wheat resistance to spot blotch or foliar blight. In Disease Resistance in Wheat; Sharma, I., Ed.; CABI: Wallingford, UK; Cambridge, MA, USA, 2012; pp. 120–135. [Google Scholar]
- Maraite, H. Evolution of the nomenclature used for Helminthosporium spp. Causing leaf blight of wheat. In Helminthosporium Blights of Wheat: Spot Blotch and Tan Spot; Duveiller, E., Dubin, H.J., Reeves, J., McNab, A., Eds.; CIMMYT: El Batán, Mexico, 1998; pp. 6–9. [Google Scholar]
- Raemaekers, R.H. Helminthosporium sativum: Disease complex on wheat and sources of resistance in Zambia. In Wheat Production Constraints in Tropical Environments; Klatt, A.R., Ed.; CIMMYT: El Batán, Mexico, 1988; pp. 175–186. [Google Scholar]
- Duveiller, E.M.; Sharma, R.C. Genetic improvement and crop management strategies to minimize yield losses in warm non-traditional wheat growing areas due to spot blotch pathogen Cochliobolus sativus. J. Phytopathol. 2009, 157, 521–534. [Google Scholar] [CrossRef]
- Sharma, R.; Duveiller, E. Selection index for improving Helminthosporium leaf blight resistance, maturity, and kernel weight in spring wheat. Crop. Sci. 2003, 43, 2031–2036. [Google Scholar] [CrossRef]
- Joshi, A.K.; Ortiz-Ferrara, G.; Crossa, J.; Singh, G.; Alvarado, G.; Bhatta, M.R.; Duveiller, E.; Sharma, R.C.; Pandit, D.B.; Siddique, A.B.; et al. Associations of environments in South Asia based on spot blotch disease of wheat caused by Cochliobolus sativus. Crop. Sci. 2007, 47, 1071–1081. [Google Scholar] [CrossRef]
- Sharma, R.C.; Duveiller, E.; Jacquemin, J.M. Microsatellite markers associated with spot blotch resistance in spring wheat. J. Phytopathol. 2007, 155, 316–319. [Google Scholar] [CrossRef]
- Kumar, U.; Joshi, A.K.; Kumar, S.; Chand, R.; Röder, M.S. Mapping of resistance to spot blotch disease caused by Bipolaris sorokiniana in spring wheat. Theor. Appl. Genet. 2009, 118, 783–792. [Google Scholar] [CrossRef]
- Kumar, U.; Joshi, A.K.; Kumar, S.; Chand, R.; Röder, M.S. Quantitative trait loci for resistance to spot blotch caused by Bipolaris sorokiniana in wheat (T. aestivum L.) lines ‘Ning 8201’ and ‘Chirya 3’. Mol. Breed. 2010, 26, 477–491. [Google Scholar]
- Lillemo, M.; Joshi, A.K.; Prasad, R.; Chand, R.; Singh, R.P. QTL for spot blotch resistance in bread wheat line SAAR co-locate to the biotrophic disease resistance loci Lr34 and Lr46. Theor. Appl. Genet. 2013, 126, 711–719. [Google Scholar] [CrossRef]
- Joshi, A.K.; Chand, R.; Kumar, S.; Singh, R.P. Leaf tip necrosis: A phenotypic marker associated with resistance to spot blotch disease in wheat. Crop. Sci. 2004, 44, 792–796. [Google Scholar] [CrossRef]
- Zhu, Z.; Bonnett, D.; Ellis, M.; Singh, P.; Heslot, N.; Dreisigacker, S.; Gao, C.; Mujeeb-Kazi, A. Mapping resistance to spot blotch in a CIMMYT synthetic-derived bread wheat. Mol. Breed. 2014, 34, 1215–1228. [Google Scholar] [CrossRef]
- Kumar, S.; Röder, M.S.; Tripathi, S.B.; Kumar, S.; Chand, R.; Joshi, A.K.; Kumar, U. Mendelization and fine mapping of a bread wheat spot blotch disease resistance QTL. Mol. Breed. 2015, 35, 218. [Google Scholar] [CrossRef]
- Lu, P.; Liang, Y.; Li, D.; Wang, Z.; Li, W.; Wang, G.; Wang, Y.; Zhou, S.; Wu, Q.; Xie, J.; et al. Fine genetic mapping of spot blotch resistance gene Sb3 in wheat (Triticum aestivum). Theor. Appl. Genet. 2016, 129, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Singh, G.; Chaudhury, A.; Ojha, A.; Tyagi, B.S.; Chowdhary, A.K.; Sheoran, S. Phenotyping at hot spots and tagging of QTLs conferring spot blotch resistance in bread wheat. Mol. Biol. Rep. 2016, 43, 1293–1303. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, T.B.; Gurung, S.; Hansen, J.M.; Jackson, E.W.; Bonman, J.M. Association mapping of quantitative trait loci in spring wheat landraces conferring resistance to bacterial leaf streak and spot blotch. Plant Genome 2012, 5, 1–16. [Google Scholar] [CrossRef]
- Gurung, S.; Mamidi, S.; Bonman, J.M.; Xiong, M.; Brown-Guedira, G.; Adhikari, T.B. Genome-wide association study reveals novel quantitative trait loci associated with resistance to multiple leaf spot diseases of spring wheat. PLoS ONE 2014, 9, e108179. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.C.; Ahren, D.; Simpfendorfer, S.; Milgate, A.; Solomon, P.S. The discovery of the virulence gene ToxA in the wheat and barley pathogen Bipolaris sorokiniana. Mol. Plant Pathol. 2017, 19, 432–439. [Google Scholar] [CrossRef]
- Friesen, T.L.; Holmes, D.J.; Bowden, R.L.; Faris, J.D. ToxA is present in the U.S. Bipolaris sorokiniana population and is a significant virulence factor on wheat harboring Tsn1. Plant Dis. 2018, 102, 2446–2452. [Google Scholar]
- Joshi, A.K.; Chand, R. Variation and inheritance of leaf angle, and its association with spot blotch (Bipolaris sorokiniana) severity in wheat (Triticum aestivum). Euphytica 2002, 124, 283–291. [Google Scholar] [CrossRef]
- Joshi, A.K.; Chand, R.; Arun, B. Relationship of plant height and days to maturity with resistance to spot blotch in wheat. Euphytica 2002, 123, 221–228. [Google Scholar] [CrossRef]
- Singh, P.K.; Zhang, Y.; He, X.; Singh, R.P.; Chand, R.; Mishra, V.K.; Malaker, P.K.; Reza, M.A.; Rahman, M.M.; Islam, R.; et al. Development and characterization of the 4th CSISA-spot blotch nursery of bread wheat. Eur. J. Plant Pathol. 2015, 143, 595–605. [Google Scholar] [CrossRef]
- He, X.; Lillemo, M.; Shi, J.; Wu, J.; Bjørnstad, Å.; Belova, T.; Dreisigacker, S.; Duveiller, E.; Singh, P. QTL characterization of Fusarium head blight resistance in CIMMYT bread wheat line Soru#1. PLoS ONE 2016, 11, e0158052. [Google Scholar]
- He, X.; Singh, P.K.; Dreisigacker, S.; Singh, S.; Lillemo, M.; Duveiller, E. Dwarfing genes Rht-B1b and Rht-D1b are associated with both type I FHB susceptibility and low anther extrusion in two bread wheat populations. PLoS ONE 2016, 11, e0162499. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.M.; Chartrain, L.; Lasserre-Zuber, P.; Saintenac, C. Genetics of resistance to Zymoseptoria tritici and applications to wheat breeding. Fungal Genet. Biol. 2015, 79, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Dubin, H.; Rajaram, S. Breeding disease-resistant wheats for tropical highlands and lowlands. Annu. Rev. Phytopathol. 1996, 34, 503–526. [Google Scholar] [CrossRef] [PubMed]
- Mujeeb-Kazi, A.; Villareal, R.; Gilchrist, L.; Rajaram, S. Registration of five wheat germplasm lines resistant to Helminthosporium leaf blight. Crop. Sci. 1996, 36, 216–217. [Google Scholar] [CrossRef]
- Joshi, A.K.; Kumar, S.; Chand, R.; Ortiz-Ferrara, G. Inheritance of resistance to spot blotch caused by Bipolaris sorokiniana in spring wheat. Plant Breed. 2004, 123, 213–219. [Google Scholar] [CrossRef]
- Saxesena, R.R.; Mishra, V.K.; Chand, R.; Chowdhury, A.K.; Bhattacharya, P.M.; Joshi, A.K. Pooling together spot blotch resistance, high yield with earliness in wheat for eastern Gangetic plains of south Asia. Field Crop. Res. 2017, 214, 291–300. [Google Scholar] [CrossRef]
- Ayana, G.T.; Ali, S.; Sidhu, J.S.; Gonzalez Hernandez, J.L.; Turnipseed, B.; Sehgal, S.K. Genome-wide association study for spot blotch resistance in hard winter wheat. Front. Plant Sci. 2018, 9, 926. [Google Scholar] [CrossRef]
- Jamil, M.; Ali, A.; Gul, A.; Ghafoor, A.; Ibrahim, A.M.H.; Mujeeb-Kazi, A. Genome-wide association studies for spot blotch (Cochliobolus sativus) resistance in bread wheat using genotyping-by-sequencing. Phytopathology 2018, 108, 1307–1314. [Google Scholar] [CrossRef]
- Vasistha, N.K.; Balasubramaniam, A.; Mishra, V.K.; Srinivasa, J.; Chand, R.; Joshi, A.K. Molecular introgression of leaf rust resistance gene Lr34 validates enhanced effect on resistance to spot blotch in spring wheat. Euphytica 2017, 213, 262. [Google Scholar] [CrossRef]
- Singh, R.P.; Huerta-Espino, J.; Bhavani, S.; Herrera-Foessel, S.A.; Singh, D.; Singh, P.K.; Velu, G.; Mason, R.E.; Jin, Y.; Njau, P.; et al. Race non-specific resistance to rust diseases in CIMMYT spring wheats. Euphytica 2011, 179, 175–186. [Google Scholar] [CrossRef]
- Dreisigacker, S.; Sukumaran, S.; Guzmán, C.; He, X.; Lan, C.; Bonnett, D.; Crossa, J. Molecular marker-based selection tools in spring bread wheat improvement: CIMMYT experience and prospects. In Molecular Breeding for Sustainable Crop Improvement; Rajpal, V., Rao, S., Raina, S., Eds.; Springer: Berlin, Germany, 2016; pp. 421–474. [Google Scholar]
- Vasistha, N.K.; Balasubramaniam, A.; Mishra, V.K.; Chand, R.; Srinivasa, J.; Yadav, P.S.; Joshi, A.K. Enhancing spot blotch resistance in wheat by marker-aided backcross breeding. Euphytica 2015, 207, 119–133. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Kozak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Saari, E.E.; Prescott, J.M. A scale for appraising the foliar intensity of wheat disease. Plant Dis. Rep. 1975, 59, 377–380. [Google Scholar]
- Li, H.; Vikram, P.; Singh, R.P.; Kilian, A.; Carling, J.; Song, J.; Burgueno-Ferreira, J.A.; Bhavani, S.; Huerta-Espino, J.; Payne, T.; et al. A high density gbs map of bread wheat and its application for dissecting complex disease resistance traits. BMC Genom. 2015, 16, 216. [Google Scholar] [CrossRef] [PubMed]
- Van Ooijen, J. JoinMap® 4, Software for the Calculation of Genetic Linkage Maps in Experimental Populations; Kyazma BV: Wageningen, The Netherlands, 2006. [Google Scholar]
- Van Ooijen, J. MapQTL® 6, Software for the Mapping of Quantitative Trait Loci in Experimental Populations of Diploid Species; Kyazma BV: Wageningen, The Netherlands, 2009. [Google Scholar]
- Voorrips, R.E. Mapchart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Source | DF | Mean Square | F Value | p Value | Heritability |
|---|---|---|---|---|---|---|
| BC | Genotype | 231 | 139905 | 15.38 | <0.0001 | 0.86 |
| Year | 2 | 5939021 | 652.79 | <0.0001 | ||
| Genotype × Year | 462 | 19415 | 2.13 | <0.0001 | ||
| Rep (Year) | 3 | 258974 | 28.47 | <0.0001 | ||
| Error | 693 | 9098 | ||||
| WC | Genotype | 230 | 145412 | 18.75 | <0.0001 | 0.88 |
| Year | 2 | 6256059 | 806.64 | <0.0001 | ||
| Genotype × Year | 460 | 17525 | 2.26 | <0.0001 | ||
| Rep (Year) | 3 | 156580 | 20.19 | <0.0001 | ||
| Error | 690 | 7756 |
| Associated traits | The BC Population | The WC Population | ||||
|---|---|---|---|---|---|---|
| 2013 | 2014 | 2015 | 2013 | 2014 | 2015 | |
| Days to heading | −0.49 ** | −0.33 ** | −0.49 ** | −0.47 ** | −0.48 ** | −0.16 |
| Plant height | −0.12 | −0.43 ** | −0.13 | −0.29 ** | −0.49 ** | −0.16 |
| Population | Linkage Group | Position | Left Marker | Right Marker | 2013 | 2014 | 2015 | Mean | R Source a | Traits Associated b |
|---|---|---|---|---|---|---|---|---|---|---|
| BC | 1B | 0.4–2.8 | 995296 | 5410777 | 7.1 | 5.4 | 6.5 | C | ||
| 1B | 209.4–218.6 | 5324988 | 1110815 | 5.5 | 8.5 | 7.0 | 8.9 | B | ||
| 1D | 48.4–53.3 | 100142243 | 1037975 | 3.5 | 2.9 | C | ||||
| 3A | 103.2–146.0 | 1109808 | 990692 | 5.1 | 3.2 | C | ||||
| 4B | 111.3–117.1 | 1247021 | 1702494 | 5.2 | 2.2 | 3.0 | 3.3 | B | ||
| 4D | 0–9.8 | BS00036421_51 | 1119387 | 3.6 | B | PH | ||||
| 5A | 175.9–179.4 | Vrn-A1 | 3064415 | 21.9 | 8.9 | 6.9 | 12.5 | B | DH | |
| 6D | 21.9–33.2 | 1239681 | 1095962 | 2.4 | 2.6 | 2.7 | B | |||
| Accumulated percentage of variation explained | 40.1 | 32.9 | 28.5 | 37.3 | ||||||
| WC | 1B | 232.3–252.9 | 1283166 | 2263671 | 2.9 | 4.4 | 4.5 | 8.3 | W | |
| 2D | 2.0–3.2 | 1085831 | 1098973 | 7.3 | 4.9 | 4.7 | 9 | W | DH | |
| 4B | 47.9–54.1 | Rht-B1 | 1241652 | 2.6 | 2.7 | 2.3 | C | PH | ||
| 4B | 99.2–107.4 | 1217659 | 997944 | 4.7 | 6 | 4.5 | W | |||
| 5A | 147.5–148.4 | 1135154 | 2260918 | 27.1 | 21.9 | 24.3 | 25.1 | W | DH | |
| Accumulated percentage of variation explained | 37.3 | 38.5 | 42.2 | 49.2 | ||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.K.; He, X.; Sansaloni, C.P.; Juliana, P.; Dreisigacker, S.; Duveiller, E.; Kumar, U.; Joshi, A.K.; Singh, R.P. Resistance to Spot Blotch in Two Mapping Populations of Common Wheat Is Controlled by Multiple QTL of Minor Effects. Int. J. Mol. Sci. 2018, 19, 4054. https://doi.org/10.3390/ijms19124054
Singh PK, He X, Sansaloni CP, Juliana P, Dreisigacker S, Duveiller E, Kumar U, Joshi AK, Singh RP. Resistance to Spot Blotch in Two Mapping Populations of Common Wheat Is Controlled by Multiple QTL of Minor Effects. International Journal of Molecular Sciences. 2018; 19(12):4054. https://doi.org/10.3390/ijms19124054
Chicago/Turabian StyleSingh, Pawan Kumar, Xinyao He, Carolina Paola Sansaloni, Philomin Juliana, Susanne Dreisigacker, Etienne Duveiller, Uttam Kumar, Arun Kumar Joshi, and Ravi Prakash Singh. 2018. "Resistance to Spot Blotch in Two Mapping Populations of Common Wheat Is Controlled by Multiple QTL of Minor Effects" International Journal of Molecular Sciences 19, no. 12: 4054. https://doi.org/10.3390/ijms19124054
APA StyleSingh, P. K., He, X., Sansaloni, C. P., Juliana, P., Dreisigacker, S., Duveiller, E., Kumar, U., Joshi, A. K., & Singh, R. P. (2018). Resistance to Spot Blotch in Two Mapping Populations of Common Wheat Is Controlled by Multiple QTL of Minor Effects. International Journal of Molecular Sciences, 19(12), 4054. https://doi.org/10.3390/ijms19124054