Proteomic Differences in Feline Fibrosarcomas Grown Using Doxorubicin-Sensitive and -Resistant Cell Lines in the Chick Embryo Model

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. In Ovo Assay
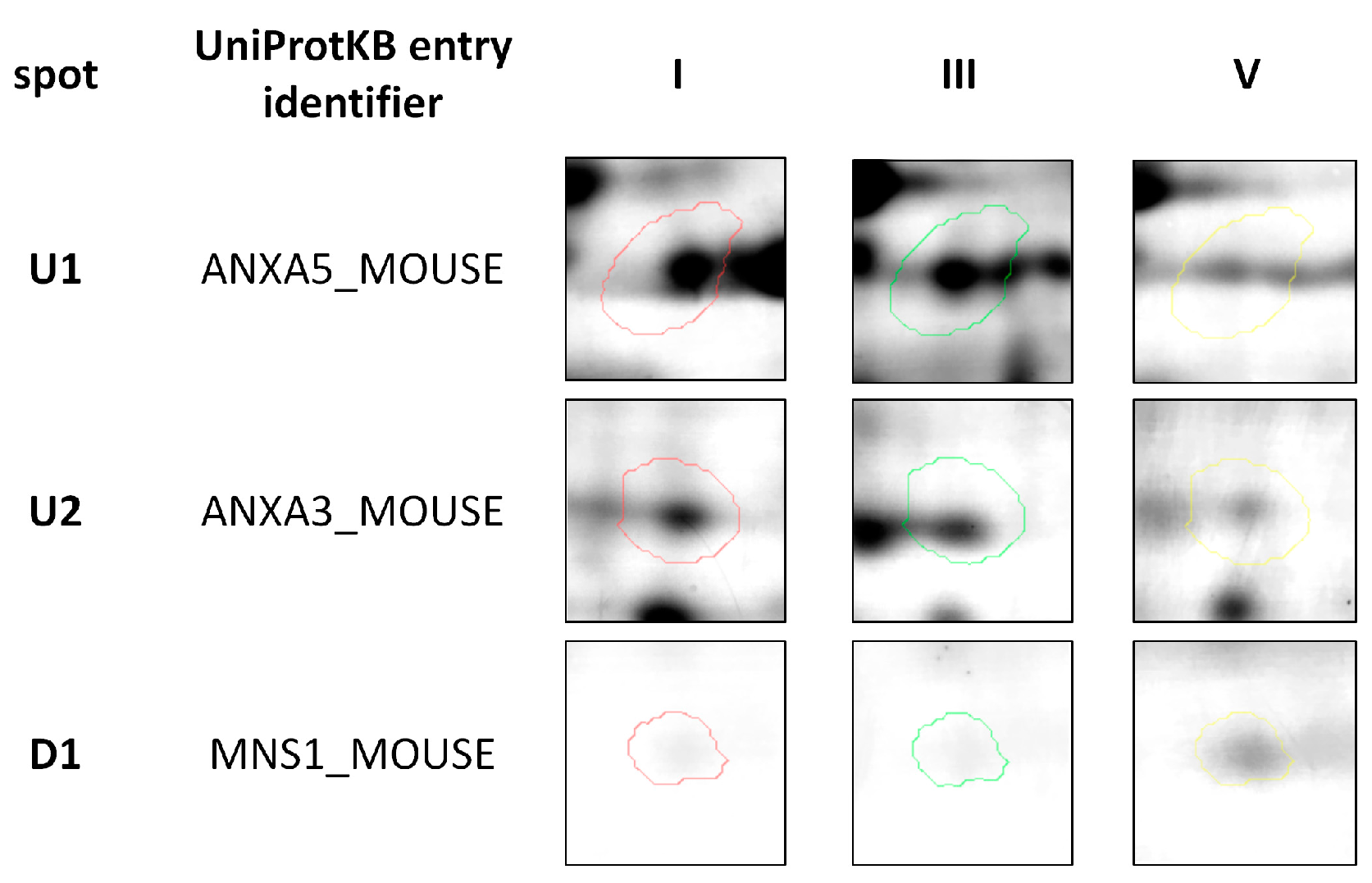
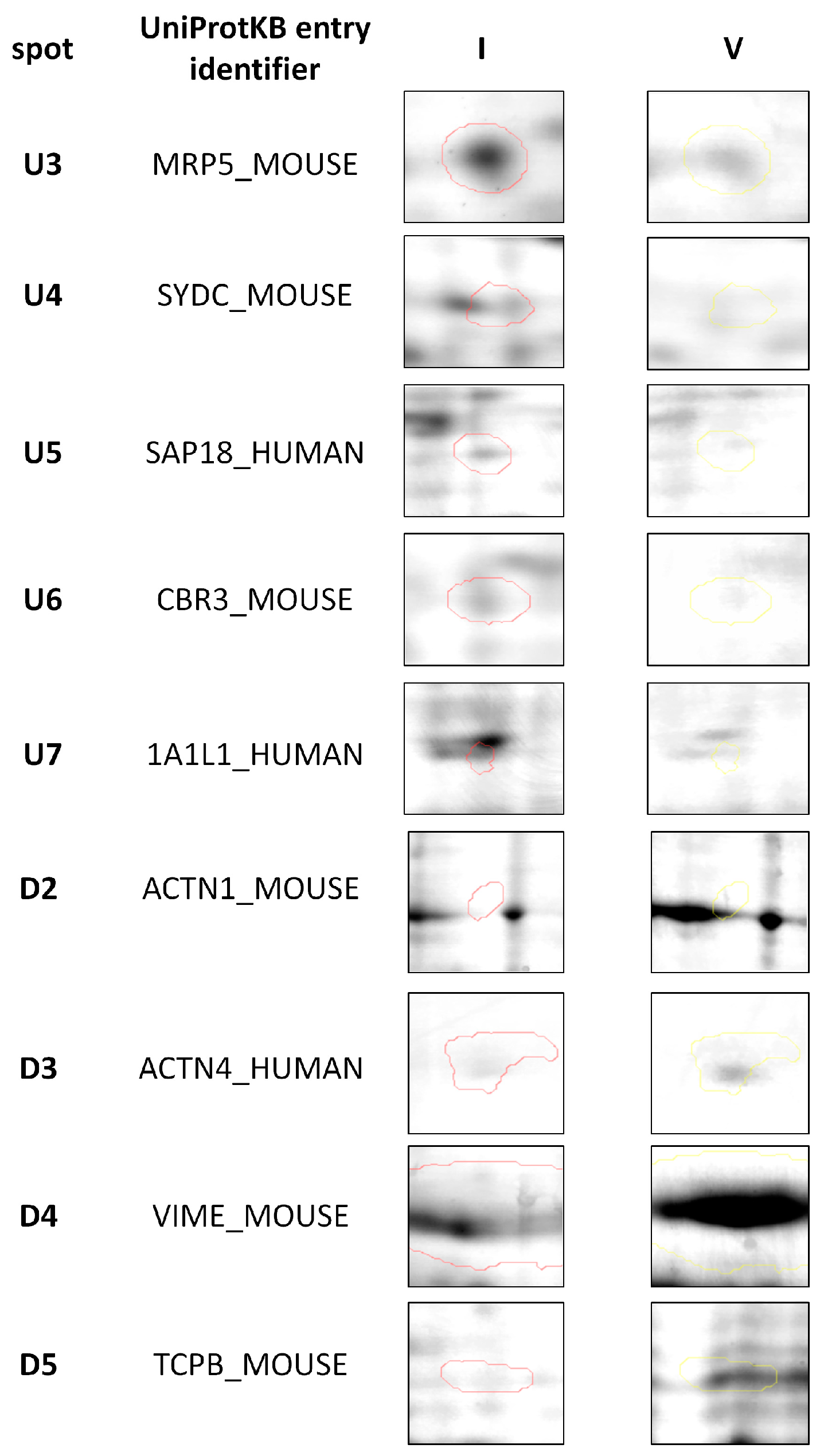
2.2. Protein Identification by 2DE and MALDI-TOF MS Analysis
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. In Ovo Assay
4.3. Protein Cleaning and Precipitation
4.4. Two-Dimensional Gel Electrophoresis (2DE)
4.5. Proteomics Analysis
4.6. MALDI-TOF MS Analysis
4.7. Visual and Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 2DE | Two-dimensional gel electrophoresis |
| 5-FU | 5-fluorouracil |
| ABC | Adenosine triphosphate-binding cassette |
| Ac (P N-term) | Acetylation of protein N-term |
| ACN | Acetonitryle |
| ACTA1 | Actin 1 |
| ACTN4 | Actinin 4 |
| ANXA3 | Annexin A3 |
| ANXA5 | Annexin A5 |
| ATP | Adenosine triphosphate |
| C (C) | Carboamidomethylation of cysteine |
| CAM | Chorioallantoic membrane |
| Diox (M) | Dioxidation of methionine |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| FISH | Fluorescence in situ hybridization |
| FISS | Feline injection-site sarcoma |
| IC50 | The half maximal inhibitory concentration |
| IPG | Immobilized pH gradient |
| MALDI-TOF MS | Matrix-assisted laser desorption ionization-time-of-flight-mass-spectrometry |
| MDR | Multidrug resistance |
| MNS1 | Meiosis-specific nuclear structural protein 1 |
| MS | Mass spectrometry |
| Ox (M) | Oxidation of methionine |
| MW | Molecular weight |
| Ph (ST) | Phosphorylation of serine and threonine |
| pI | Isoelectic point |
| RT-PCR | Reverse transcription-polymerase chain reaction |
| SCP1 | Synaptonemal complex protein 1 |
| Sp17 | Sperm protein 17 |
| TFA | Trifluoroacetic acid |
| TRIS | Tris(hydroxymethyl)aminomethane |
References
- Li, D.; Chan, D.W. Proteomic cancer biomarkers from discovery to approval: It’s worth the effort. Expert Rev. Proteom. 2014, 11, 135–136. [Google Scholar] [CrossRef] [PubMed]
- Füzéry, A.K.; Levin, J.; Chan, M.M.; Chan, D.W. Translation of proteomic biomarkers into FDA approved cancer diagnostics: Issues and challenges. Clin. Proteom. 2013, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Sallam, R.M. Proteomics in cancer biomarkers discovery: Challenges and applications. Dis. Mark. 2015, 2015, 321370. [Google Scholar] [CrossRef] [PubMed]
- Conrads, T.P.; Zhou, M.; Petricoin, E.F., III; Liotta, L.; Veenstra, T.D. Cancer diagnosis using proteomic patterns. Expert Rev. Mol. Diagn. 2003, 3, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Posadas, E.M.; Simpkins, F.; Liotta, L.A.; MacDonald, C.; Kohn, E.C. Proteomic analysis for the early detection and rational treatment of cancer—Realistic hope? Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2005, 16, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Hondermarck, H.; Vercoutter-Edouart, A.S.; Revillion, F.; Lemoine, J.; el-Yazidi-Belkoura, I.; Nurcombe, V.; Peyrat, J.P. Proteomics of breast cancer for marker discovery and signal pathway profiling. Proteomics 2001, 1, 1216–1232. [Google Scholar] [CrossRef]
- Raval, G.N.; Bharadwaj, S.; Levine, E.A.; Willingham, M.C.; Geary, R.L.; Kute, T.; Prasad, G.L. Loss of expression of tropomyosin-1, a novel class II tumor suppressor that induces anoikis, in primary breast tumors. Oncogene 2003, 22, 6194–6203. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Albrechtsen, R.; Kronqvist, P.; Cox, J.; Mann, M.; Geiger, T. Proteomic maps of breast cancer subtypes. Nat. Commun. 2016, 7, 10259. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, J.; Kikuchi, T.; Shyr, Y.; Carbone, D. Proteomic patterns of chemosensitivity in NSCLC cell lines. Cancer Res. 2006, 66, 1065 LP. [Google Scholar]
- Hodgkinson, V.C.; Agarwal, V.; ELFadl, D.; Fox, J.N.; McManus, P.L.; Mahapatra, T.K.; Kneeshaw, P.J.; Drew, P.J.; Lind, M.J.; Cawkwell, L. Pilot and feasibility study: Comparative proteomic analysis by 2-DE MALDI TOF/TOF MS reveals 14-3-3 proteins as putative biomarkers of response to neoadjuvant chemotherapy in ER-positive breast cancer. J. Proteom. 2012, 75, 2745–2752. [Google Scholar] [CrossRef] [PubMed]
- McCaw, D.L.; Chan, A.S.; Stegner, A.L.; Mooney, B.; Bryan, J.N.; Turnquist, S.E.; Henry, C.J.; Alexander, H.; Alexander, S. Proteomics of canine lymphoma identifies potential cancer-specific protein markers. Clin. Cancer Res. 2007, 13, 2496–2503. [Google Scholar] [CrossRef] [PubMed]
- Klose, P.; Weise, C.; Bondzio, A.; Multhaup, G.; Einspanier, R.; Gruber, A.D.; Klop, R. Is There a Malignant Progression Associated with a Linear Change in Protein Expression Levels from Normal Canine Mammary Gland to Metastatic Mammary Tumors? J. Proteome Res. 2011, 10, 4405–4415. [Google Scholar] [CrossRef] [PubMed]
- Klopfleisch, R.; Klose, P.; Weise, C.; Bondzio, A.; Multhaup, G.; Einspanier, R.; Gruber, A.D. Proteome of metastatic canine mammary carcinomas: Similarities to and differences from human breast cancer. J. Proteome Res. 2010, 9, 6380–6391. [Google Scholar] [CrossRef] [PubMed]
- Schlieben, P.; Meyer, A.; Weise, C.; Bondzio, A.; Einspanier, R.; Gruber, A.D.; Klopfleisch, R. Differences in the proteome of high-grade versus low-grade canine cutaneous mast cell tumours. Vet. J. 2012, 194, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, L.; Mian, S.; Slater, K.; King, H.; Napolitano, M.; Aucoin, D.; Mobasheri, A. Proteomic identification and profiling of canine lymphoma patients. Vet. Comp. Oncol. 2009, 7, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Leroy, B.; Painter, A.; Sheppard, H.; Popiolek, L.; Andacht, T.M. Protein expression profi ling of normal and neoplastic canine prostate and bladder tissue. Vet. Comp. Oncol. 2007, 5, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Gerou-Ferriani, M.; Mcbrearty, A.R.; Burchmore, R.J.; Jayawardena, K.G.I.; Eckersall, P.D.; Morris, J.S. Agarose gel serum protein electrophoresis in cats with and without lymphoma and preliminary results of tandem mass fingerprinting analysis. Vet. Clin. Pathol. 2011, 40, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Zabielska-Koczywąs, K.; Wojtalewicz, A.; Lechowski, R. Current knowledge on feline injection-site sarcoma treatment. Acta Vet. Scand. 2017, 59, 47. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, C.G.; Levy, J.K.; Tucker, S.J.; Olmstead, S.M.; Crawford, P.C.; Dubovi, E.J.; Hanlon, C.A. Tail vaccination in cats: A pilot study. J. Feline Med. Surg. 2014, 16, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Liang, X.-J. Overcoming drug efflux-based multidrug resistance in cancer with nanotechnology. Chin. J. Cancer 2012, 31, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Kabata, H.; Shimamoto, N. Visualization of DNA-protein interaction and sliding motion of protein molecules along DNA. Tanpakushitsu Kakusan Koso 1997, 42, 1181–1186. [Google Scholar] [PubMed]
- Han, L.; Zhang, Y.; Wang, N.; Guo, X.-J.; Zang, C.-H.; Jiang, J.-H.; Ma, F.; Wang, Q.-D. Using rhodamine 123 accumulation in CD8+ cells as a surrogate indicator to study the P-glycoprotein modulating effect of cepharanthine hydrochloride in vivo. J. Biomed. Biotechnol. 2011, 2011, 281651. [Google Scholar] [CrossRef]
- Pawłowski, K.M.; Mucha, J.; Majchrzak, K.; Motyl, T.; Król, M. Expression and role of PGP, BCRP, MRP1 and MRP3 in multidrug resistance of canine mammary cancer cells. BMC Vet. Res. 2013, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wu, J.; Zhang, W.; Luo, H.; Shen, Z.; Cheng, H.; Zhu, X. PKM2 enhances chemosensitivity to cisplatin through interaction with the mTOR pathway in cervical cancer. Sci. Rep. 2016, 6, 30788. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Marzese, D.M.; Ohta, K.; Huang, S.K.; Sim, M.S.; Chong, K.; Hoon, D.S.B. Epigenetic regulation of REG1A and chemosensitivity of cutaneous melanoma. Epigenetics 2013, 8, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Williams, K.C.; Gavin, C.T.; Jardine, E.; Chambers, A.F.; Leong, H.S. Quantification of cancer cell extravasation in vivo. Nat. Protoc. 2016, 11, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. Chicken chorioallantoic membrane angiogenesis model. Methods Mol. Biol. 2012, 843, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Kue, C.S.; Tan, K.Y.; Lam, M.L.; Lee, H.B. Chick embryo chorioallantoic membrane (CAM): An alternative predictive model in acute toxicological studies for anti-cancer drugs. Exp. Anim. 2015, 64, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, M.; Becker, J.; Eberth, S.; Kube, D.; Wilting, J. The chick chorioallantoic membrane as an in vivo xenograft model for Burkitt lymphoma. BMC Cancer 2014, 14, 339. [Google Scholar] [CrossRef] [PubMed]
- Deryugina, E.I.; Quigley, J.P. Chick embryo chorioallantoic membrane model systems to study and visualize human tumor cell metastasis. Histochem. Cell Biol. 2008, 130, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Deryugina, E.I.; Zijlstra, A.; Partridge, J.J.; Kupriyanova, T.A.; Madsen, M.A.; Papagiannakopoulos, T.; Quigley, J.P. Unexpected effect of matrix metalloproteinase down-regulation on vascular intravasation and metastasis of human fibrosarcoma cells selected in vivo for high rates of dissemination. Cancer Res. 2005, 65, 10959–10969. [Google Scholar] [CrossRef] [PubMed]
- Mangieri, D.; Nico, B.; Coluccia, A.M.L.; Vacca, A.; Ponzoni, M.; Ribatti, D. An alternative in vivo system for testing angiogenic potential of human neuroblastoma cells. Cancer Lett. 2009, 277, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zhou, X.; Ming, H.; Zhang, J.; Huang, G.; Zhang, Z.; Li, P. Chick chorioallantoic membrane assay: A 3D animal model for study of human nasopharyngeal carcinoma. PLoS ONE 2015, 10, e0130935. [Google Scholar] [CrossRef] [PubMed]
- Walewska, M.; Dolka, I.; Małek, A.; Wojtalewicz, A.; Wojtkowska, A.; Żbikowski, A.; Lechowski, R.; Zabielska-Koczywąs, K. Experimental tumor growth of canine osteosarcoma cell line on chick embryo chorioallantoic membrane (in vivo studies). Acta Vet. Scand. 2017, 59, 30. [Google Scholar] [CrossRef] [PubMed]
- Zabielska-Koczywąs, K.; Dolka, I.; Król, M.; Żbikowski, A.; Lewandowski, W.; Mieczkowski, J.; Wójcik, M.; Lechowski, R. Doxorubicin Conjugated to Glutathione Stabilized Gold Nanoparticles (Au-GSH-Dox) as an Effective Therapeutic Agent for Feline Injection-Site Sarcomas—Chick Embryo Chorioallantoic Membrane Study. Molecules 2017, 22, 253. [Google Scholar] [CrossRef] [PubMed]
- Zabielska, K.; Lechowski, R.; Krol, M.; Pawlowski, K.M.; Motyl, T.; Dolka, I.; Zbikowski, A. Derivation of feline vaccine-associated fibrosarcoma cell line and its growth on chick embryo chorioallantoic membrane—A new in vivo model for veterinary oncological studies. Vet. Res. Commun. 2012, 36, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.Y.; Blacking, T.M.; Else, R.W.; Sherman, A.; Sang, H.M.; Whitelaw, B.A. Feline mammary carcinoma stem cells are tumorigenic, radioresistant, chemoresistant and defective in activation of the ATM/p53 DNA damage pathway. Vet. J. 2013, 196, 414–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, L.Y.; Gatenby, E.L.; Kamida, A.; Whitelaw, B.A.; Hupp, T.R.; Argyle, D.J. Global gene expression analysis of canine osteosarcoma stem cells reveals a novel role for COX-2 in tumour initiation. PLoS ONE 2014, 9, e83144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabielska-Koczywąs, K.; Wojtkowska, A.; Dolka, I.; Małek, A.; Walewska, M.; Wojtalewicz, A.; Zbikowski, A.; Lechowski, R. 3D chick embryo chorioallantoic membrane model as an in vivo model to study morphological and histopathological features of feline fibrosarcomas. BMC Vet. Res. 2017, 13, 201. [Google Scholar] [CrossRef] [PubMed]
- Corsten, M.F.; Hofstra, L.; Narula, J.; Reutelingsperger, C.P.M. Counting heads in the war against cancer: Defining the role of annexin A5 imaging in cancer treatment and surveillance. Cancer Res. 2006, 66, 1255–1260. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, E.; Kuramitsu, Y.; Okada, F.; Fujimoto, M.; Zhang, X.; Kobayashi, M.; Iizuka, N.; Ueyama, Y.; Nakamura, K. Proteomic profiling for cancer progression: Differential display analysis for the expression of intracellular proteins between regressive and progressive cancer cell lines. Proteomics 2005, 5, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Yang, L.; Xiong, Y.; Guo, H.; Shen, X.; Cheng, Z.; Zhang, Y.; Gao, Z.; Zhu, X. Annexin A5 promotes invasion and chemoresistance to temozolomide in glioblastoma multiforme cells. Tumor Biol. 2014, 35, 12327–12337. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Huang, W.; Zhong, M.; Yin, L.; Jiang, H.; Hou, S.; Gan, P.; Yuan, Y. Identification Keratin 1 as a cDDP-resistant protein in nasopharyngeal carcinoma cell lines. J. Proteom. 2012, 75, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Suzuki, T.; Komiya, T.; Hatashita, E.; Nishio, K.; Kazuhiko, N.; Fukuoka, M. Induction of MRP5 and SMRP mRNA by adriamycin exposure and its overexpression in human lung cancer cells resistant to Adriamycin. Int. J. Cancer 2001, 94, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Liu, X.; Ma, J.; Liu, P.; Hu, C.; Zhang, G. Annexin A5 inhibits diffuse large B-cell lymphoma cell invasion and chemoresistance through phosphatidylinositol 3-kinase signaling. Oncol. Rep. 2014, 32, 2557–2563. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Bai, Y.; Zhang, L.; Gong, L.; Qi, X.; Li, H.; Wang, F.; Chi, X.; Jiang, Y.; Shao, S. Quantitative proteomic profiling the molecular signatures of annexin A5 in lung squamous carcinoma cells. PLoS ONE 2016, 11, e0163622. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Liu, S.; Guo, C.; Hou, Z.; Sun, M.Z. The role of annexin A3 playing in cancers. Clin. Transl. Oncol. 2013, 15, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Yip, K.; Das, P.K.; Suria, D.; Lim, C.; Ng, G.; Liew, C. A case-controlled validation study of a blood- based seven-gene biomarker panel for colorectal cancer in Malaysia. J. Exp. Clin. Cancer Res. 2010, 29, 128. [Google Scholar] [CrossRef] [PubMed]
- Baine, M.J.; Chakraborty, S.; Smith, L.M.; Mallya, K.; Sasson, A.R.; Brand, R.E.; Batra, S.K. Transcriptional profiling of peripheral blood mononuclear cells in pancreatic cancer patients identifies novel genes with potential diagnostic utility. PLoS ONE 2011, 6, e17014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.F.; Xiao, Z.Q.; Li, M.X.; Li, M.Y.; Zhang, P.F.; Li, C.; Li, F.; Chen, Y.H.; Yi, H.; Yao, H.X.; et al. Quantitative proteome analysis reveals annexin A3 as a novel biomarker in lung adenocarcinoma. J. Pathol. 2009, 217, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.C.; Neo, J.C.; Lo, S.L.; San Tan, G.; Seow, T.K.; Chung, M.C. Proteome database of hepatocellular carcinoma. J. Chromatogr. B 2002, 771, 303–328. [Google Scholar] [CrossRef]
- Kollermann, J.; Schlomm, T.; Bang, H.; Schwall, G.P.; von Eichel-Streiber, C.; Simon, R.; Schostak, M.; Huland, H.; Berg, W.; Sauter, G.; et al. Expression and prognostic relevance of annexin A3 in prostate cancer. Eur. Urol. 2008, 54, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, C.; Bombelli, S.; Raimondo, F.; Torsello, B.; Angeloni, V.; Ferrero, S.; Di Stefano, V.; Chinello, C.; Cifola, I.; Invernizzi, L.; et al. Primary Cell Cultures from Human Renal Cortex and Renal-Cell Carcinoma Evidence a Differential Expression of Two Spliced Isoforms of Annexin A3. Am. J. Pathol. 2010, 176, 1660–1670. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Yan, X.; Yao, X.; Zhang, Y.; Shan, Y.; Mao, N.; Yang, Y.; Pan, L. Secretion of annexin A3 from ovarian cancer cells and its association with platinum resistance in ovarian cancer patients. J. Cell. Mol. Med. 2012, 16, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Inagaki, H.; Naruge, T.; Tabata, S.; Tomida, T.; Yamaguchi, A.; Yoshikuni, M.; Nagahama, Y.; Hotta, Y. cDNA cloning and functional characterization of a meiosis-specific protein (MNS1) with apparent nuclear association. Chromosome Res. 1994, 2, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yang, F.; Leu, N.A.; Wang, P.J. MNS1 is essential for spermiogenesis and motile ciliary functions in mice. PLoS Genet. 2012, 8, e1002516. [Google Scholar] [CrossRef] [PubMed]
- Shakib, K.; Norman, J.T.; Fine, L.G.; Brown, L.R.; Godovac-Zimmermann, J. Proteomics profiling of nuclear proteins for kidney fibroblasts suggests hypoxia, meiosis, and cancer may meet in the nucleus. Proteomics 2005, 5, 2819–2838. [Google Scholar] [CrossRef] [PubMed]
- Türeci, O.; Sahin, U.; Zwick, C.; Koslowski, M.; Seitz, G.; Pfreundschuh, M. Identification of a meiosis-specific protein as a member of the class of cancer/testis antigens. Proc. Natl. Acad. Sci. USA 1998, 95, 5211–5216. [Google Scholar] [CrossRef] [PubMed]
- Straughn, J.M.; Shaw, D.R.; Guerrero, A.; Bhoola, S.M.; Racelis, A.; Wang, Z.; Chiriva-Internati, M.; Grizzle, W.E.; Alvarez, R.D.; Lim, S.H.; et al. Expression of sperm protein 17 (Sp17) in ovarian cancer. Int. J. Cancer 2004, 108, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Mirandola, L.; Pedretti, E.; Figueroa, J.A.; Chiaramonte, R.; Colombo, M.; Chapman, C.; Grizzi, F.; Patrinicola, F.; Kast, W.M.; Nguyen, D.D.; et al. Cancer testis antigen sperm protein 17 as a new target for triple negative breast cancer immunotherapy. Oncotarget 2017, 8, 74378–74390. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.; Allen, W.L.; McLean, E.G.; Wilson, P.M.; McCulla, A.; Moore, S.; Longley, D.B.; Caldas, C.; Johnston, P.G. Pharmacogenomic identification of novel determinants of response to chemotherapy in colon cancer. Cancer Res. 2006, 66, 2765–2777. [Google Scholar] [CrossRef] [PubMed]
- Honda, K. The biological role of actinin-4 (ACTN4) in malignant phenotypes of cancer. Cell Biosci. 2015, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chu, K.-M. α-Actinin-4 promotes metastasis in gastric cancer. Lab. Investig. 2017, 97, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Tsuda, H.; Honda, K.; Onozato, K.; Takano, M.; Tamai, S.; Imoto, I.; Inazawa, J.; Yamada, T.; Matsubara, O. Actinin-4 gene amplification in ovarian cancer: A candidate oncogene associated with poor patient prognosis and tumor chemoresistance. Mod. Pathol. 2009, 22, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhong, Q.; Rhodes, L.V.; Townley, I.; Bratton, M.R.; Zhang, Q.; Martin, E.C.; Elliott, S.; Collins-Burow, B.M.; Burow, M.E.; et al. Proteomic analysis of acquired tamoxifen resistance in MCF-7 cells reveals expression signatures associated with enhanced migration. Breast Cancer Res. 2012, 14, R45. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Whelan, S.A.; Lu, M.; Shen, D.; Chung, D.U.; Saxton, R.E.; Faull, K.F.; Whitelegge, J.P.; Chang, H.R. Proteomic-Based Biosignatures in Breast Cancer Classification and Prediction of Therapeutic Response. Int. J. Proteom. 2011, 2011, 896476. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, H.; Yu, J.; Yu, H. Chemoresistance to doxorubicin induces epithelial-mesenchymal transition via upregulation of transforming growth factor β signaling in HCT116 colon cancer cells. Mol. Med. Rep. 2015, 12, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Yi, W.; Peng, J.; Zhang, Y.; Fu, F.; Zou, Q.; Tang, Y. Differential protein expressions in breast cancer between drug sensitive tissues and drug resistant tissues. J. Cent. South Univ. (Med. Sci.) 2013, 38, 148–154. [Google Scholar] [CrossRef]
- Baxter, S.S.; Dibble, C.F.; Byrd, W.C.; Carlson, J.; Mack, C.R.; Saldarriaga, I.; Bencharit, S. Role of cytoskeletal proteins in cerebral cavernous malformation signaling pathways: A proteomic analysis. Mol. Biosyst. 2014, 10, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Kuramitsu, Y.; Murakami, K.; Ryozawa, S.; Taba, K.; Kaino, S.; Zhang, X.; Sakaida, I.; Nakamura, K. Proteomic differential display analysis for TS-1-resistant and -sensitive pancreatic cancer cells using two-dimensional gel electrophoresis and mass spectrometry. Anticancer Res. 2011, 31, 2103–2108. [Google Scholar] [PubMed]
- Wójcik, M.; Lewandowski, W.; Król, M.; Pawłowski, K.; Mieczkowski, J.; Lechowski, R.; Zabielska, K. Enhancing anti-tumor efficacy of doxorubicin by non-covalent conjugation to gold nanoparticles—In vitro studies on Feline fibrosarcoma cell lines. PLoS ONE 2015, 10, e0124955. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Obama, T.; Kato, R.; Masuda, Y.; Takahashi, K.; Aiuchi, T.; Itabe, H. Analysis of modified apolipoprotein B-100 structures formed in oxidized low-density lipoprotein using LC-MS/MS. Proteomics 2007, 7, 2132–2141. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| ID | Protein 1 | p Value | Accession Number (UniProtKB) | Species | Score | Match | MW (kDa)2 | pI 2 | Modif. 3 | Seq. Cov. (%) | Ratio I/V4 | RatioIII/V 4 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| U1 | ANXA5 | 0.0145 | P48036 | M. musculus | 194 | 27 | 35.79 | 4.83 | C (C); Ox (M) | 65 | 1.51 | 1.503 |
| U2 | ANXA3 | 0.0119 | O35639 | M. musculus | 78 | 16 | 36.53 | 5.50 | C (C); Ox (M) | 52 | 1.66 | 1.74 |
| D1 | MNS1 | 0.0167 | Q61884 | M. musculus | 105 | 14 | 60.26 | 6.32 | Ac (N-term); C (C) Ox (M); Ac (P N-term) | 20 | 0.29 | 0.36 |
| ID | Protein 1 | p Value | Accession Number (UniProtKB) | Species | Score | Match | MW (kDa) 1 | pI 2 | Modif. 3 | Seq. Cov. (%) | Ratio I/V 4 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| U3 | MRP5 | 0.024 | Q9R1X5 | M. musculus | 70 | 16 | 162.05 | 8.80 | C (C); Ox (M); Ac (P N-term); Diox (M) | 8 | 2.23 |
| U4 | SYDC | 0.0289 | Q922B2 | M. musculus | 62 | 12 | 57.57 | 6.07 | C (C); Ox (M); Ac (P N-term) | 32 | 1.88 |
| U5 | SAP18 | 0.0237 | O00422 | H. sapiens | 101 | 12 | 17.61 | 9.38 | C (C); Ox (M); Ac (P N-term); Ph (ST) | 47 | 2.83 |
| U6 | CBR3 | 0.0495 | Q8K354 | M. musculus | 80 | 12 | 31.33 | 6.15 | C (C); Ox (M); Diox (M) | 44 | 2.2 |
| U7 | 1A1L1 | 0.0125 | Q96QU6 | H. sapiens | 86 | 13 | 57.86 | 6.01 | C (C); Ox (M); Ac (P N-term) | 26 | 4.2 |
| D2 | ACTA1 | 0.05 | P68134 | M. musculus | 84 | 26 | 103.63 | 5.23 | C (C); Ox (M) | 29 | 0.09 |
| D3 | ACTN4 | 0.0283 | O43707 | H. sapiens | 120 | 27 | 105.37 | 5.25 | C (C); Ox (M); Ac (P N-term) | 30 | 0.37 |
| D4 | VIME | 0.0152 | P20152 | M. musculus | 175 | 29 | 53.71 | 5.06 | C (C); Ox (M); Ac (P N-term) | 52 | 0.61 |
| D5 | TCPB | 0.0344 | P80314 | M. musculus | 114 | 12 | 57.78 | 5.97 | C (C); Ox (M) | 29 | 0.20 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zabielska-Koczywąs, K.; Michalak, K.; Wojtalewicz, A.; Winiarczyk, M.; Adaszek, Ł.; Winiarczyk, S.; Lechowski, R. Proteomic Differences in Feline Fibrosarcomas Grown Using Doxorubicin-Sensitive and -Resistant Cell Lines in the Chick Embryo Model. Int. J. Mol. Sci. 2018, 19, 576. https://doi.org/10.3390/ijms19020576
Zabielska-Koczywąs K, Michalak K, Wojtalewicz A, Winiarczyk M, Adaszek Ł, Winiarczyk S, Lechowski R. Proteomic Differences in Feline Fibrosarcomas Grown Using Doxorubicin-Sensitive and -Resistant Cell Lines in the Chick Embryo Model. International Journal of Molecular Sciences. 2018; 19(2):576. https://doi.org/10.3390/ijms19020576
Chicago/Turabian StyleZabielska-Koczywąs, Katarzyna, Katarzyna Michalak, Anna Wojtalewicz, Mateusz Winiarczyk, Łukasz Adaszek, Stanisław Winiarczyk, and Roman Lechowski. 2018. "Proteomic Differences in Feline Fibrosarcomas Grown Using Doxorubicin-Sensitive and -Resistant Cell Lines in the Chick Embryo Model" International Journal of Molecular Sciences 19, no. 2: 576. https://doi.org/10.3390/ijms19020576
APA StyleZabielska-Koczywąs, K., Michalak, K., Wojtalewicz, A., Winiarczyk, M., Adaszek, Ł., Winiarczyk, S., & Lechowski, R. (2018). Proteomic Differences in Feline Fibrosarcomas Grown Using Doxorubicin-Sensitive and -Resistant Cell Lines in the Chick Embryo Model. International Journal of Molecular Sciences, 19(2), 576. https://doi.org/10.3390/ijms19020576