Inhibition of Late and Early Phases of Cancer Metastasis by the NF-κB Inhibitor DHMEQ Derived from Microbial Bioactive Metabolite Epoxyquinomicin: A Review

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
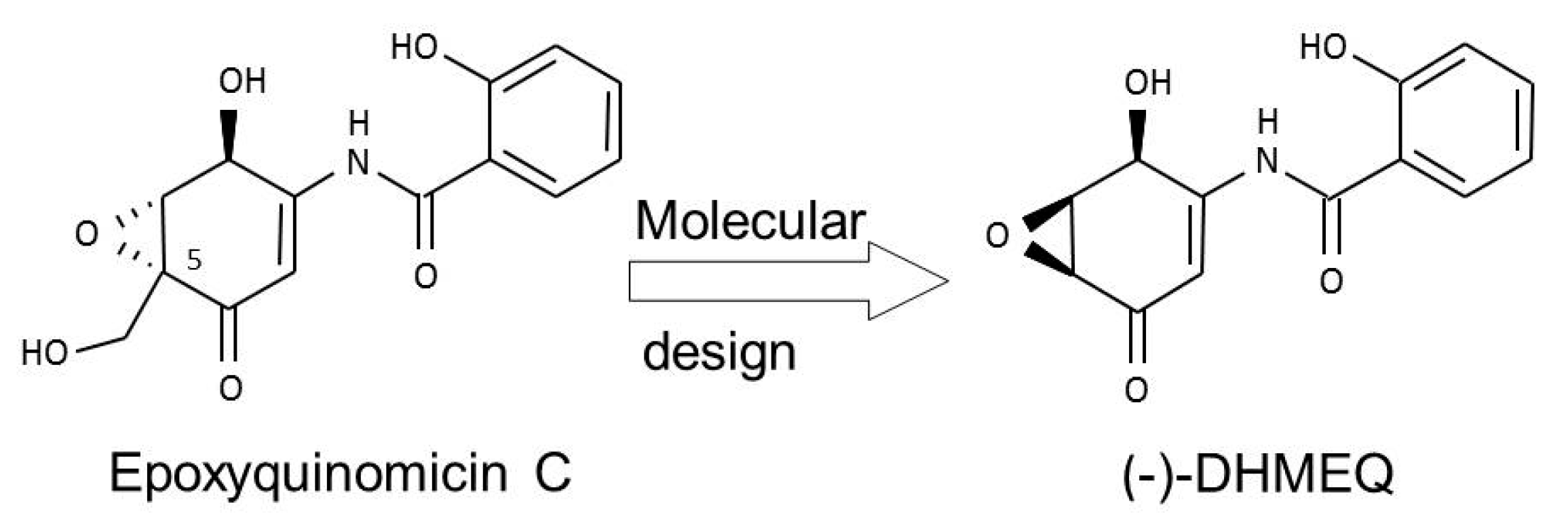
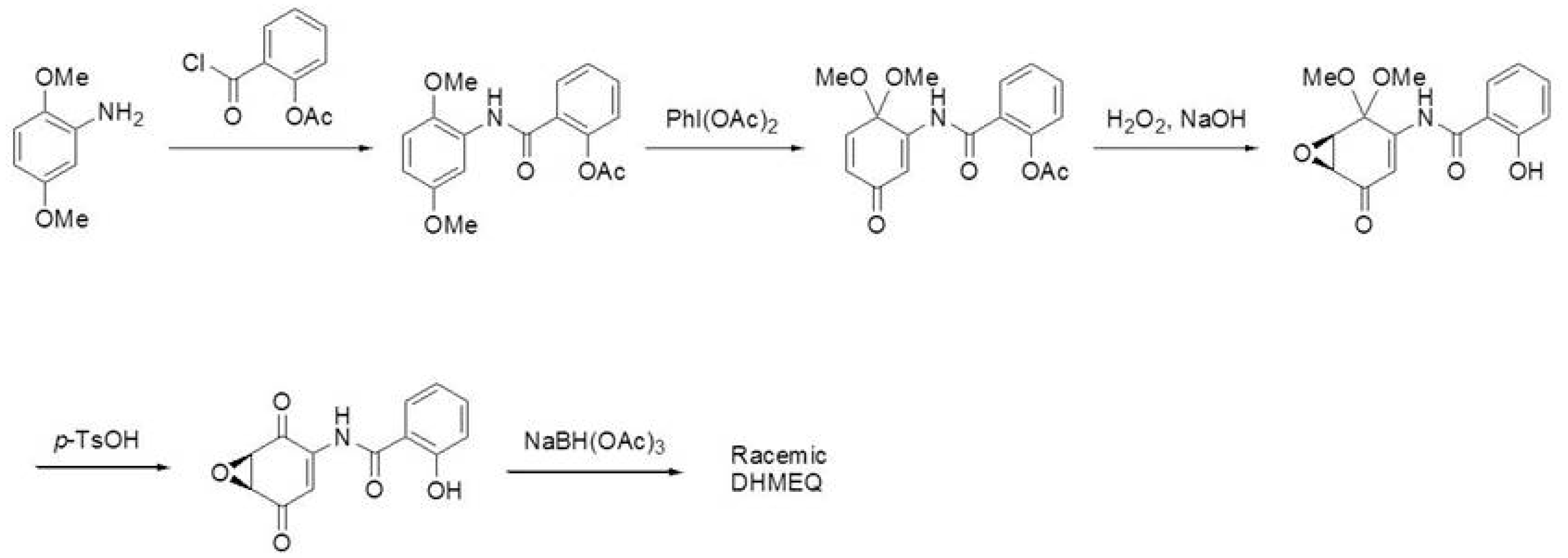
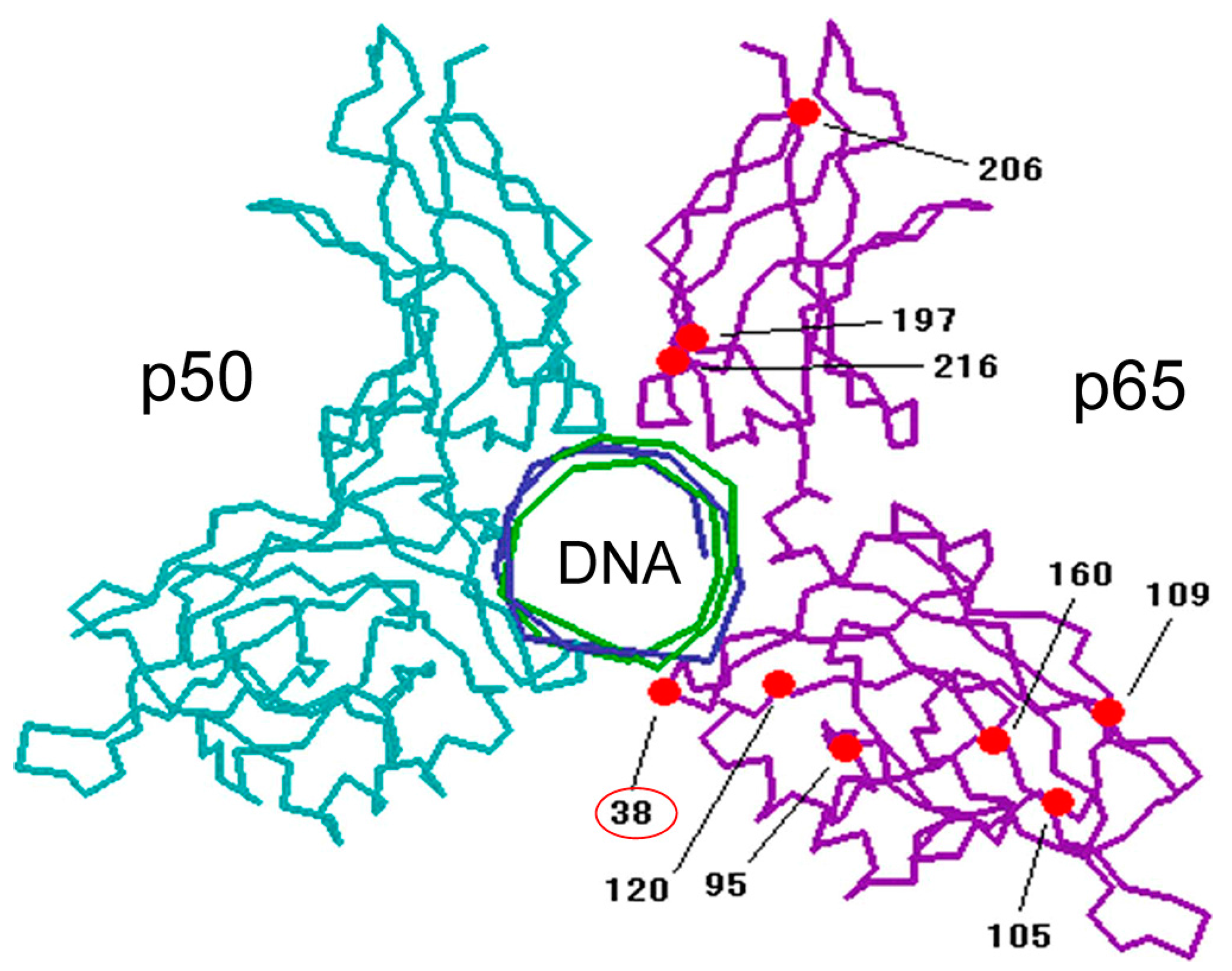
2. Discovery and Mechanism of Action of NF-κB Inhibitor DHMEQ
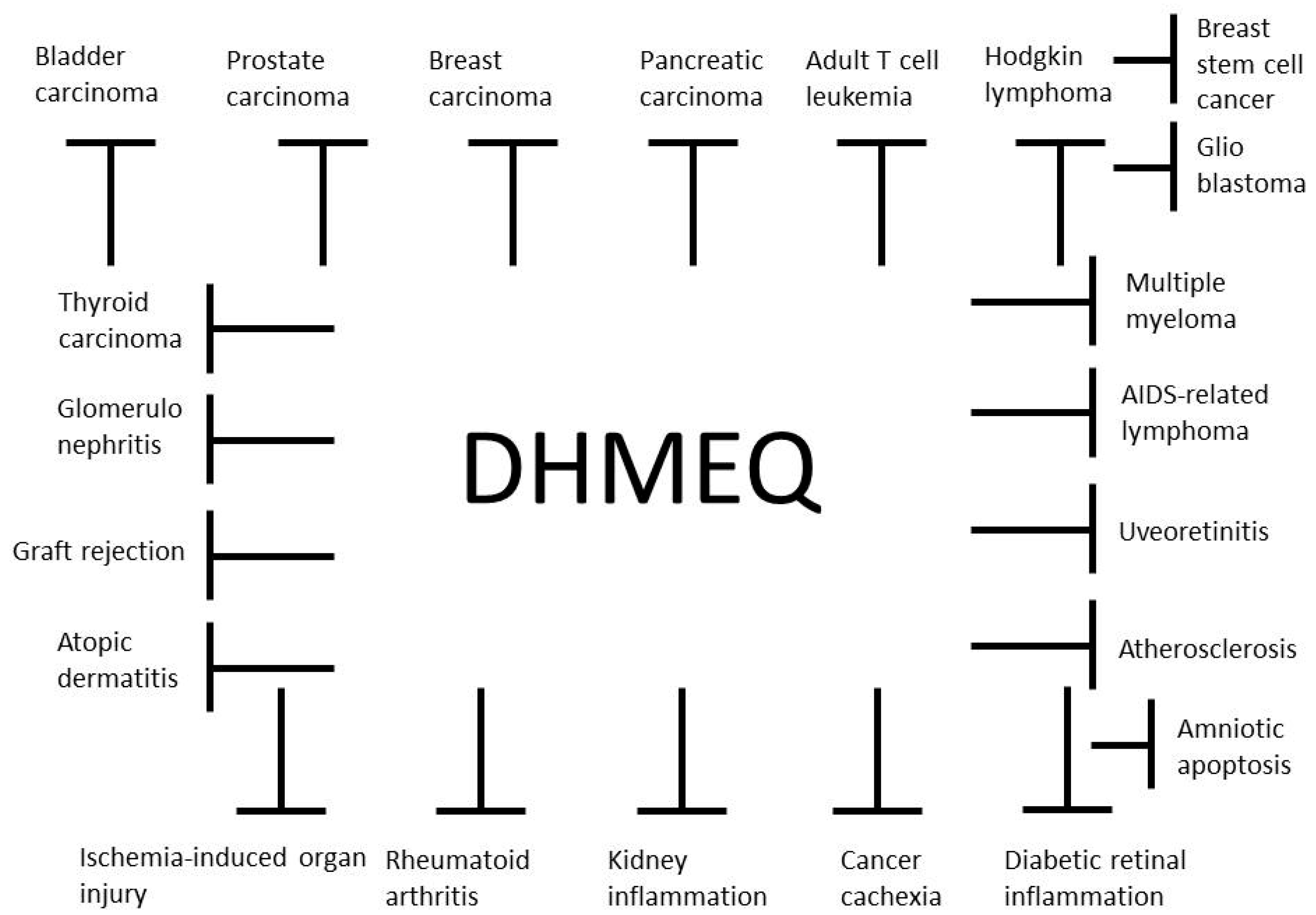
3. Therapeutic Activity of DHMEQ on Inflammatory and Neoplastic Disease Models
4. Contribution of NF-κB to Cancer Metastasis
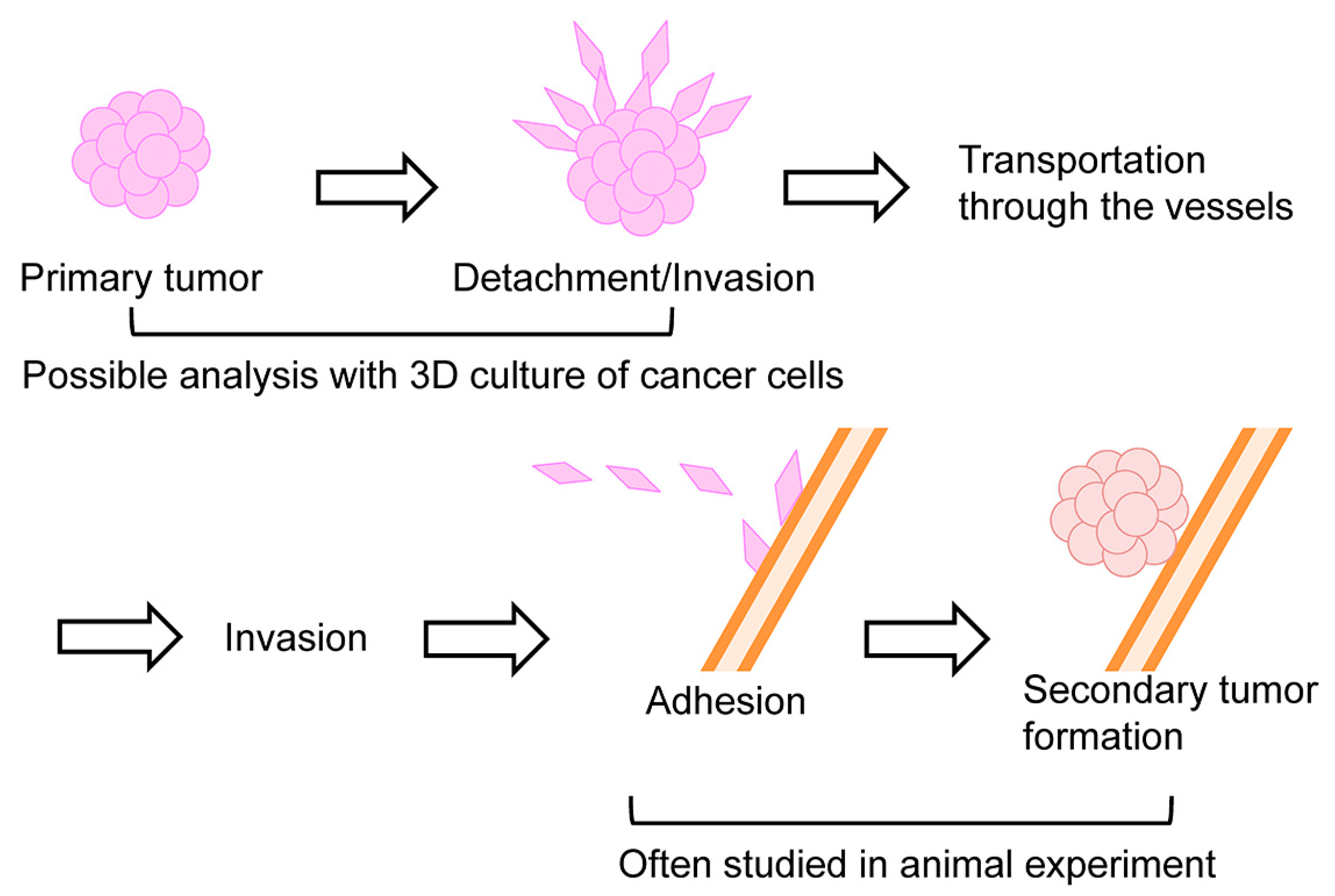
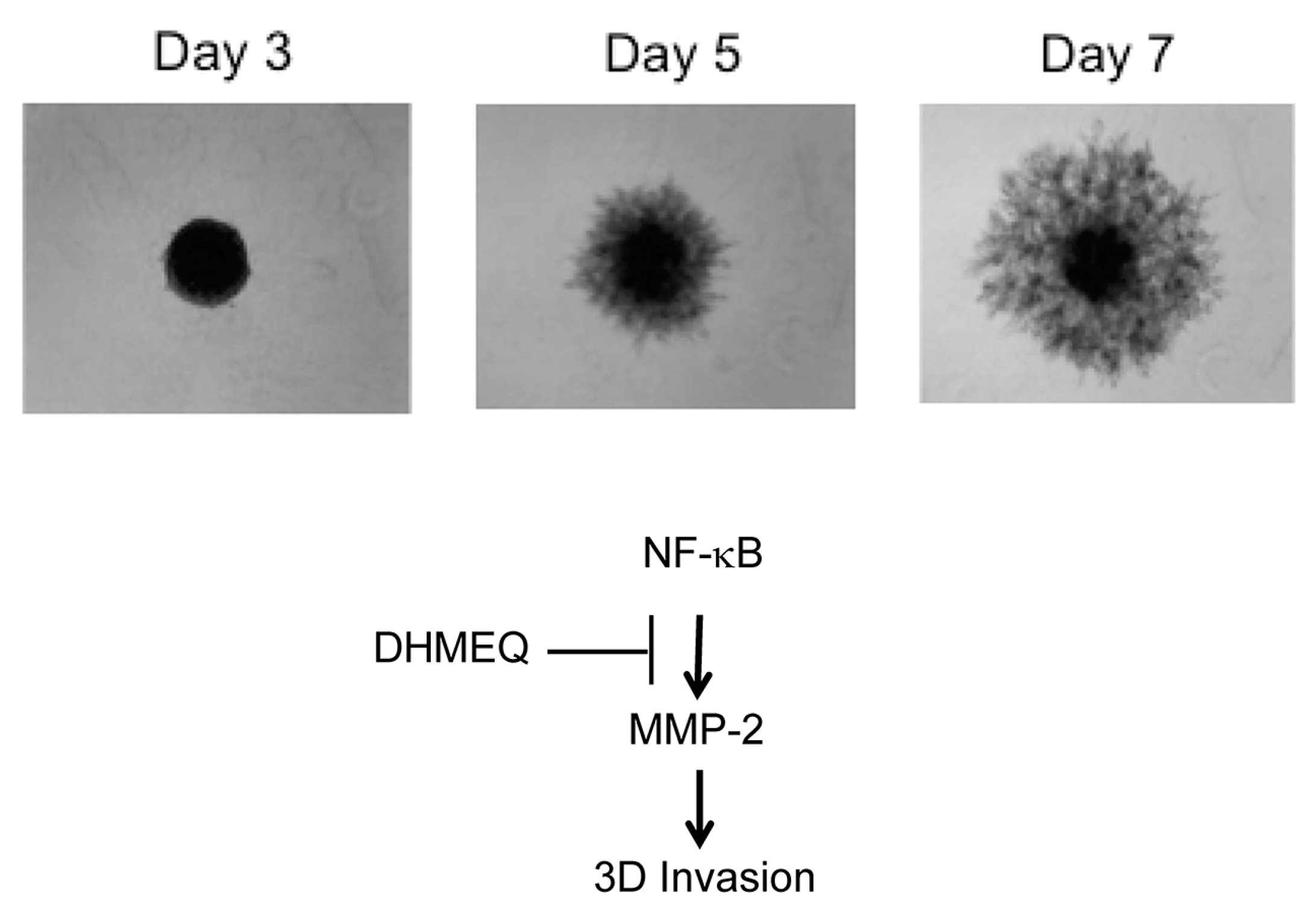
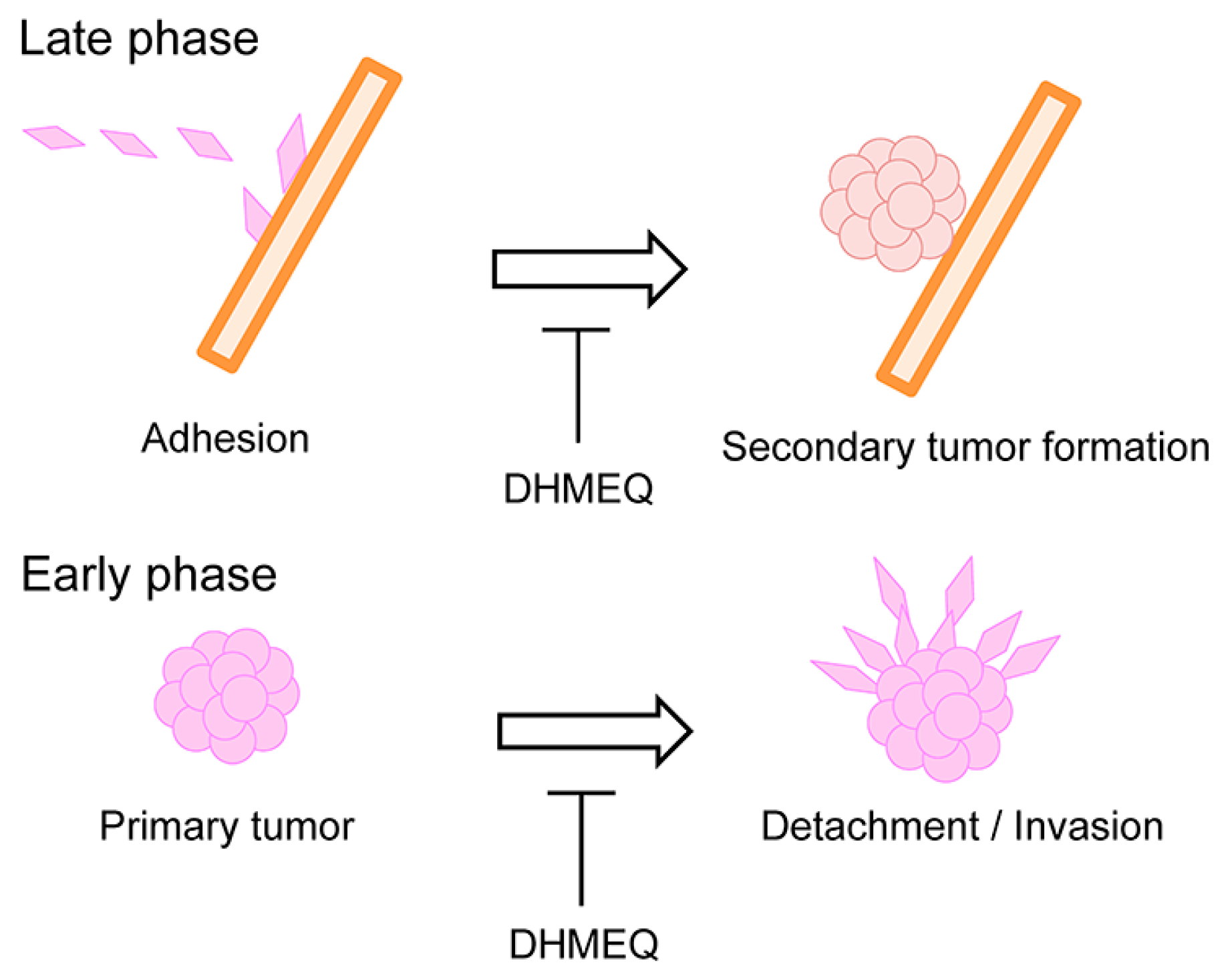
5. Inhibition of Late and Early Phases of Metastasis by DHMEQ
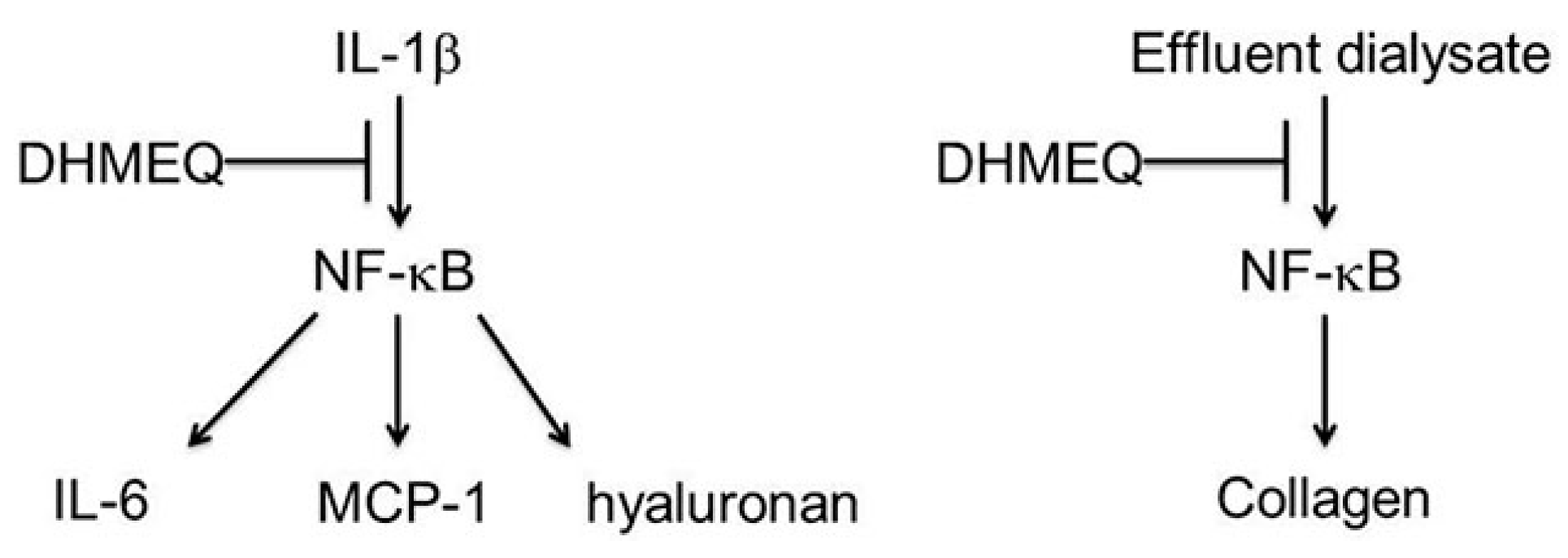
6. Suppression of Peritoneal NF-κB May Inhibit Peripheral Inflammation and Tumor Formation
7. Conclusions and Future Prospective
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Onoda, T.; Iinuma, H.; Sasaki, Y.; Hamada, M.; Isshiki, K.; Naganawa, H.; Takeuchi, T.; Tatsuta, K.; Umezawa, K. Isolation of a novel tyrosine kinase inhibitor, lavendustin A, from Streptomyces griseolavendus. J. Nat. Prod. 1989, 52, 1252–1257. [Google Scholar] [CrossRef] [PubMed]
- Imoto, M.; Kakeya, H.; Sawa, T.; Hayashi, C.; Hamada, M.; Takeuchi, T.; Umezawa, K. Dephostasin, a novel protein tyrosine phosphatase inhibitor produced by Streptomyces. I. Taxonomy, isolation, and characterization. J. Antibiot. 1993, 46, 1342–1346. [Google Scholar] [CrossRef] [PubMed]
- Ohse, T.; Ohba, S.; Yamamoto, T.; Koyano, T.; Umezawa, K. Cyclopentabenzofuran lignan protein synthesis inhibitors from Aglaia odorata. J. Nat. Prod. 1996, 59, 650–652. [Google Scholar] [CrossRef] [PubMed]
- Fukami, A.; Ikeda, Y.; Kondo, S.; Naganawa, H.; Takeuchi, T.; Furuya, S.; Hirabayashi, Y.; Shimoike, K.; Hosaka, S.; Watanabe, Y.; et al. Akaterpin, a novel bioactive triterpene from the marine sponge Callyspongia sp. Terrahedron Lett. 1997, 38, 1201–1202. [Google Scholar] [CrossRef]
- Ohno, O.; Ikeda, Y.; Sawa, R.; Igarashi, M.; Kinoshita, N.; Suzuki, Y.; Miyake, K.; Umezawa, K. Isolation of heptadepsin, a novel bacterial cyclic depsipeptide that inhibits lipopolysaccharide activity. Chem. Biol. 2004, 11, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Iinuma, H.; Ikeda, Y.; Igarashi, M.; Hatano, K.; Kinoshita, N.; Ukaji, T.; Simizu, S.; Umezawa, K. Migracins, A and B, new inhibitors of cancer cell migration, produced by Streptomyces sp. J. Antibiot. 2013, 66, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, K.; Kojima, I.; Simizu, S.; Lin, Y.Z.; Fukatsu, H.; Koide, N.; Nakade, Y.; Yoneda, M. Therapeutic a activity of plant-derived alkaloid conophylline on metabolic syndrome and neurodegenerative disease models. Hum. Cell 2017. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, N.; Ariga, A.; To-e, S.; Nakamura, H.; Agata, N.; Hirano, S.; Inoue, J.; Umezawa, K. Synthesis of NF-κB activation inhibitors derived from epoxyquinomicin C. Bioorg. Med. Chem. Lett. 2000, 10, 865–869. [Google Scholar] [CrossRef]
- Erkel, G.; Anke, T.; Sterner, O. Inhibition of NF-κB activation by panepoxydone. Biochem. Biophys. Res. Commun. 1996, 226, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Gehrt, A.; Erkel, G.; Anke, T.; Sterner, O.A. Cycloepoxydon: 1-hydroxy-2-hydroxymethyl-3-pent-1-enylbenzene1-hydroxy-2-hydroxymethyl-3-pent-1,3-dienylbenzene, new inhibitors of eukaryotic signal transduction. J. Antibiot. 1998, 51, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Sugiyama, C.; Ohno, O.; Umezawa, K. Preparation and biological activities of optically active dehydroxymethylepoxyquinomicin, a novel NF-κB inhibitor. Tetrahedron 2004, 60, 7061–7066. [Google Scholar] [CrossRef]
- Hamada, M.; Niitsu, Y.; Hiraoka, C.; Kozawa, I.; Higashi, T.; Shoji, M.; Umezawa, K.; Sugai, T. Chemoenzymatic synthesis of (2S,3S,4S)-form, the physiologically active stereoisomer of dehydroxymethylepoxyquinomicin (DHMEQ), a potent inhibitor on NF-κB. Tetrahedron 2010, 66, 7083–7087. [Google Scholar] [CrossRef]
- Ariga, A.; Namekawa, J.; Matsumoto, N.; Inoue, J.; Umezawa, K. Inhibition of TNF-α-induced nuclear translocation and activation of NF-κB by dehydroxymethyl-epoxyquinomicin. J. Biol. Chem. 2002, 277, 27625–27630. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Horie, R.; Takeiri, M.; Kozawa, I.; Umezawa, K. Inactivation of nuclear factor kappa B components by covalent binding of (−)-dehydroxymethylepoxyquinomicin to specific cysteine residues. J. Med. Chem. 2008, 51, 5780–5788. [Google Scholar] [CrossRef] [PubMed]
- Horie, K.; Ma, J.; Umezawa, K. Inhibition of canonical NF-κB nuclear localization by (−)-DHMEQ via impairment of DNA binding. Oncol. Res. 2015, 22, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Kozawa, I.; Kato, K.; Teruya, T.; Suenaga, K.; Umezawa, K. Unusual intramolecular N→O acyl group migration occurring during conjugation of (−)-DHMEQ with cysteine. Bioorg. Med. Chem. Lett. 2009, 19, 5380–5382. [Google Scholar] [CrossRef] [PubMed]
- Shimada, C.; Ninomiya, Y.; Suzuki, E.; Umezawa, K. Efficient cellular uptake of the novel NF-κB inhibitor (−)-DHMEQ and irreversible inhibition of NF-κB in neoplastic cells. Oncol. Res. 2010, 18, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Takeiri, M.; Horie, K.; Takahashi, D.; Watanabe, M.; Horie, R.; Simizu, S.; Umezawa, K. Involvement of DNA binding domain in the cellular stability and importin affinity of NF-κB component RelB. Org. Biomol. Chem. 2012, 10, 3053–3059. [Google Scholar] [CrossRef] [PubMed]
- Hamasaka, A.; Yoshioka, N.; Abe, R.; Kishino, S.; Umezawa, K.; Ozaki, M.; Todo, S.; Shimizu, H. Topical application of DHMEQ improves allergic inflammation via NF-κB inhibition. J. Allergy Clin. Immunol. 2010, 126, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Noma, N.; Asagiri, M.; Takeiri, M.; Ohmae, S.; Takemoto, K.; Iwaisako, K.; Simizu, S.; Umezawa, K. Inhibition of MMP-2-mediated mast cell invasion by NF-κB inhibitor DHMEQ in mast cells. Int. Achieves Allergy Immunol. 2015, 166, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wei, B.; Lan, Y.; Dai, C.; Gu, Y.; Ma, J.; Liu, X.; Umezawa, K.; Zhang, Y. External application of NF-κB inhibitor DHMEQ suppresses development of atopic dermatitis-like lesions induced with DNCB/OX in BALB/c mice. Immunopharmacol. Immunotoxicol. 2017, 39, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Umezawa, K.; Yasui, M. Apoptosis in mouse amniotic epithelium is induced by activated macrophages through the TNF receptor type 1/TNF pathway. Biol. Reprod. 2011, 84, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Yamanouchi, S.; Adachi, Y.; Shimo, T.; Umezawa, K.; Okigaki, M.; Tsuji, S.; Li, M.; Takaya, J.; Kuge, T.; Ikehara, S.; Kaneko, K. A nuclear factor-κB inhibitor, dehydroxymethylepoxyquinomicin, ameliorates GVHD in allogeneic bone marrow transplantation. Immunobiology 2015, 220, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- El-Salhy, M.; Umezawa, K. Effects of AP-1 and NF-kappa B inhibitors on colonic endocrine cells in rats with TNBS-induced colitis. Mol. Med. Rep. 2016, 14, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- El-Salhy, M.; Umezawa, K. Anti-inflammatory effects of novel AP-1 and NF-κB inhibitors in dextran-sulfate-sodium-induced colitis in rats. Int. J. Mol. Med. 2016, 37, 1457–1464. [Google Scholar] [CrossRef] [PubMed]
- Nishio, H.; Yaguchi, T.; Sugiyama, J.; Sumimoto, H.; Umezawa, K.; Iwata, T.; Susumu, N.; Fujii, T.; Kawamura, N.; Kobayashi, A.; et al. Immunosuppression through constitutively activated NF-κB signaling in human ovarian cancer and its reversal by a NF-κB inhibitor. Br. J. Cancer 2014, 110, 2965–2974. [Google Scholar] [CrossRef] [PubMed]
- Seubwai, W.; Kraiklang, R.; Vaeteewoottacharn, K.; Umezawa, K.; Okada, S.; Wongkham, S. Aberrant expression of NF-κB in liver fluke associated cholangiocarcinoma: Implications for targeted therapy. PLoS ONE 2014, 9, e106056. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Kikuchi, E.; Tanaka, N.; Kosaki, T.; Suzuki, E.; Mizuno, R.; Shinojima, T.; Miyajima, A.; Umezawa, K.; Oya, M. Down-regulation of NF kappa B activation is an effective therapeutic modality in acquired platinum-resistant bladder cancer. BMC Cancer 2015, 15, 324. [Google Scholar] [CrossRef] [PubMed]
- Miyake, A.; Dewan, A.; Ishida, T.; Watanabe, M.; Honda, M.; Sata, T.; Yamamoto, N.; Umezawa, K.; Watanabe, T.; Horie, R. Induction of apoptosis in Epstein-Barr virus-infected B-lymphocytes by the NF-κB inhibitor DHMEQ. Microbes Infect. 2008, 10, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Ohsugi, T.; Shoda, M.; Ishida, T.; Aizawa, S.; Maruyama-Nagai, M.; Utsunomiya, A.; Koga, S.; Yamada, Y.; Kamihira, S.; et al. Dual targeting of transformed and untransformed HTLV-1-infected T-cells by DHMEQ, a potent and selective inhibitor of NF-κB, as a strategy for chemoprevention and therapy of adult T cell leukemia. Blood 2005, 106, 2462–2471. [Google Scholar] [CrossRef] [PubMed]
- Saito, D.; Sawamura, M.; Umezawa, K.; Kanai, Y.; Furihata, C.; Matsushima, T.; Sugimura, T. Inhibition of experimental blood-borne lung metastasis by protease inhibitors. Cancer Res. 1980, 40, 2539–2542. [Google Scholar] [PubMed]
- Suzuki, K.; Aiura, K.; Matsuda, S.; Itano, O.; Takeuchi, O.; Umezawa, K.; Kitagawa, Y. Combined effect of dehydroxymethylepoxyquinomicin and gemcitabine in a mouse model of liver metastasis of pancreatic cancer. Clin. Exp. Metastasis 2013, 30, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Nakanishi, K.; Haga, S.; Fujiyoshi, M.; Baba, M.; Mino, K.Y.; Niwa, H.; Yokoo, H.; Umezawa, K.; Ohmiya, Y.; et al. Anoikis induction and inhibition of peritoneal metastasis of pancreatic cancer cells by a nuclear factor-kappa B inhibitor, (−)-DHMEQ. Oncol. Res. 2014, 21, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Ukaji, T.; Lin, Y.Z.; Okada, S.; Umezawa, K. Inhibition of MMP-2-mediated cellular invasion by NF-κB inhibitor DHMEQ in 3D culture of breast carcinoma MDA-MB-231 cells: A model for early phase of metastasis. Biochem. Biophys. Res. Commun. 2017, 485, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, K.; Nanki, T.; Miyasaka, N.; Umezawa, K.; Kubota, T. Effect of a small molecule inhibitor of nuclear factor-κB nuclear translocation in a murine model of arthritis and cultured human synovial cells. Arthritis Res. Ther. 2003, 7, 1348–1359. [Google Scholar] [CrossRef] [PubMed]
- Ohsugi, T.; Horie, R.; Kumasaka, T.; Ishida, A.; Ishida, T.; Yamaguchi, K.; Watanabe, T.; Umezawa, K.; Urano, T. In vivo antitumor activity of the NF-κB inhibitor dehydroxymethyl-epoxyquinomicin in a mouse model of adult T-cell leukemia. Carcinogenesis 2005, 26, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, K. Possible role of peritoneal NF-κB in peripheral inflammation and cancer: Lessons from the inhibitor DHMEQ. Biomed. Pharmacother. 2011, 65, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, K. Peritoneal NF-κB as a Possible molecular target for suppression of various cancers and inflammation. Forum Immunopathol. Dis. Ther. 2013, 4, 63–77. [Google Scholar] [CrossRef]
- Watanabe, E.; Mochizuki, N.; Ajima, H.; Ohno, K.; Shiino, M.; Umezawa, K.; Fukai, M.; Ozaki, M.; Furukawa, H.; Todo, S.; et al. A simple and reliable method for determining plasma concentration of dehydroxymethylepoxyquinomicin by high performance liquid chromatography with mass spectrometry. J. Chromatogr. B 2008, 871, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Suzuki, E.; Takasugi, A.; Obata, R.; Ishikawa, Y.; Umezawa, K.; Nishiyama, S. Efficient synthesis of (±)-parasitenone, a novel inhibitor of NF-κB. Bioorg. Med. Chem. Lett. 2009, 19, 5383–5386. [Google Scholar] [CrossRef] [PubMed]
- Park, B.K.; Zhang, H.; Zeng, Q.; Dai, J.; Keller, E.T.; Giordano, T.; Gu, K.; Shah, V.; Pei, L.; Zarbo, R.J.; et al. NF-κB in breast cancer cells promotes osteolytic bone metastasis by inducing osteoclastogenesis via GM-CSF. Nat. Med. 2007, 13, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Zins, K.; Abraham, D.; Sioud, M.; Aharinejad, S. Colon cancer cell–derived tumor necrosis factor-α mediates the tumor growth–promoting response in macrophages by up-regulating the colony-stimulating factor-1 pathway. Cancer Res. 2007, 67, 1038–1045. [Google Scholar] [CrossRef] [PubMed]
- Ukaji, T.; Umezawa, K. Novel approaches to target NF-κB and other signaling pathways in cancer stem cells. Adv. Biol. Regul. 2014, 56, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Oike, Y.; Izumi-Nagai, K.; Koto, T.; Satofuka, S.; Ozawa, Y.; Yamashiro, K.; Inoue, M.; Tsubota, K.; Umezawa, K.; et al. Suppression of diabetes-induced retinal inflammation by blocking angiotensin II type 1 receptor or its downstream NF-κB pathway. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4342–4350. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, T.; Kawaguchi, M.; Yorita, K.; Tanaka, H.; Umezawa, K.; Kataoka, H. Antitumor effect of dehydroxymethylepoxyquinomicin (DHMEQ), a small molecule inhibitor of nuclear factor-κB, on glioblastoma. Neuro-Oncology 2012, 14, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Horiguchi, Y.; Nakashima, J.; Kikuchi, E.; Kanao, K.; Miyajima, A.; Ohigashi, T.; Umezawa, K.; Murai, M. Prevention of cancer cachexia by a novel nuclear factor κB inhibitor in prostate cancer. Clin. Cancer Res. 2005, 11, 5590–5594. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, E.; Sugiyama, C.; Umezawa, K. Inhibition of inflammatory mediator secretion by (−)-DHMEQ in mouse bone marrow-derived macrophages. Biomed. Pharmacother. 2009, 63, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Sosinnska, P.; Macckowiak, B.; Staniszewski, R.; Umezawa, K.; Breborowicz, A. Inhibition of NF-κB with dehydroxyepoxiquinomicin modifies function of human peritoneal mesothelial cells. Am. J. Transl. Res. 2016, 8, 5756–5765. [Google Scholar]
- Vernon-Roberts, B. Lymphocyte to macrophage transformation in the peritoneal cavity preceding the mobilization of peritoneal macrophages to inflamed areas. Nature 1969, 222, 1286–1288. [Google Scholar] [CrossRef] [PubMed]










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.; Ukaji, T.; Koide, N.; Umezawa, K. Inhibition of Late and Early Phases of Cancer Metastasis by the NF-κB Inhibitor DHMEQ Derived from Microbial Bioactive Metabolite Epoxyquinomicin: A Review. Int. J. Mol. Sci. 2018, 19, 729. https://doi.org/10.3390/ijms19030729
Lin Y, Ukaji T, Koide N, Umezawa K. Inhibition of Late and Early Phases of Cancer Metastasis by the NF-κB Inhibitor DHMEQ Derived from Microbial Bioactive Metabolite Epoxyquinomicin: A Review. International Journal of Molecular Sciences. 2018; 19(3):729. https://doi.org/10.3390/ijms19030729
Chicago/Turabian StyleLin, Yinzhi, Tamami Ukaji, Naoki Koide, and Kazuo Umezawa. 2018. "Inhibition of Late and Early Phases of Cancer Metastasis by the NF-κB Inhibitor DHMEQ Derived from Microbial Bioactive Metabolite Epoxyquinomicin: A Review" International Journal of Molecular Sciences 19, no. 3: 729. https://doi.org/10.3390/ijms19030729
APA StyleLin, Y., Ukaji, T., Koide, N., & Umezawa, K. (2018). Inhibition of Late and Early Phases of Cancer Metastasis by the NF-κB Inhibitor DHMEQ Derived from Microbial Bioactive Metabolite Epoxyquinomicin: A Review. International Journal of Molecular Sciences, 19(3), 729. https://doi.org/10.3390/ijms19030729