Molecular Characterization and Overexpression of VpRPW8s from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici in Nicotiana benthamiana

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
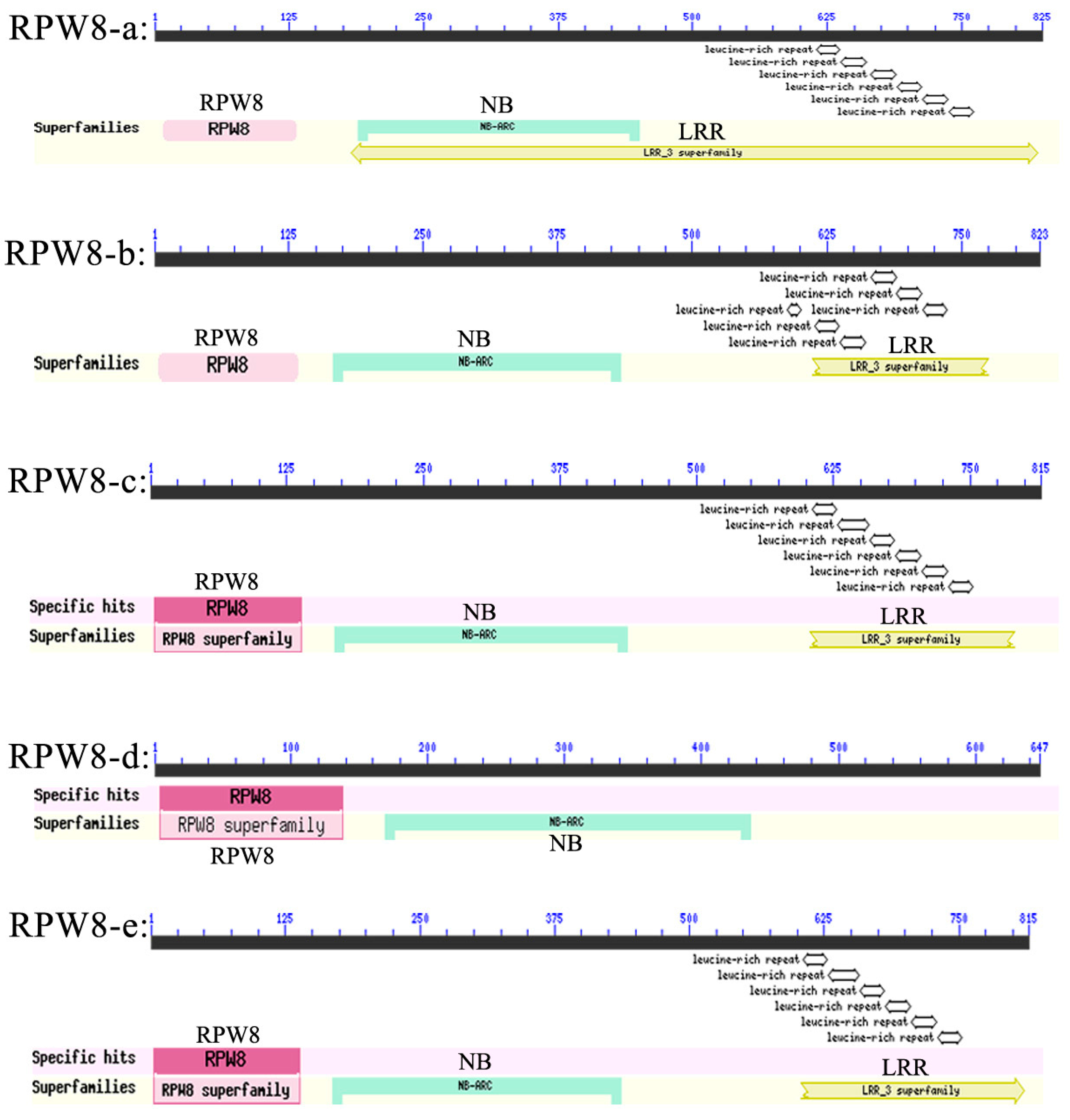
2.1. Isolation, Characterization, and Bioinformatics of VpRPW8s
2.2. Transcriptional Expression Analysis of the RPW8s in V. pseudoreticulata and V. vinifera under P. viticola Stress
2.3. Prokaryotic Expression and Subcellular Localization of VpRPW8s
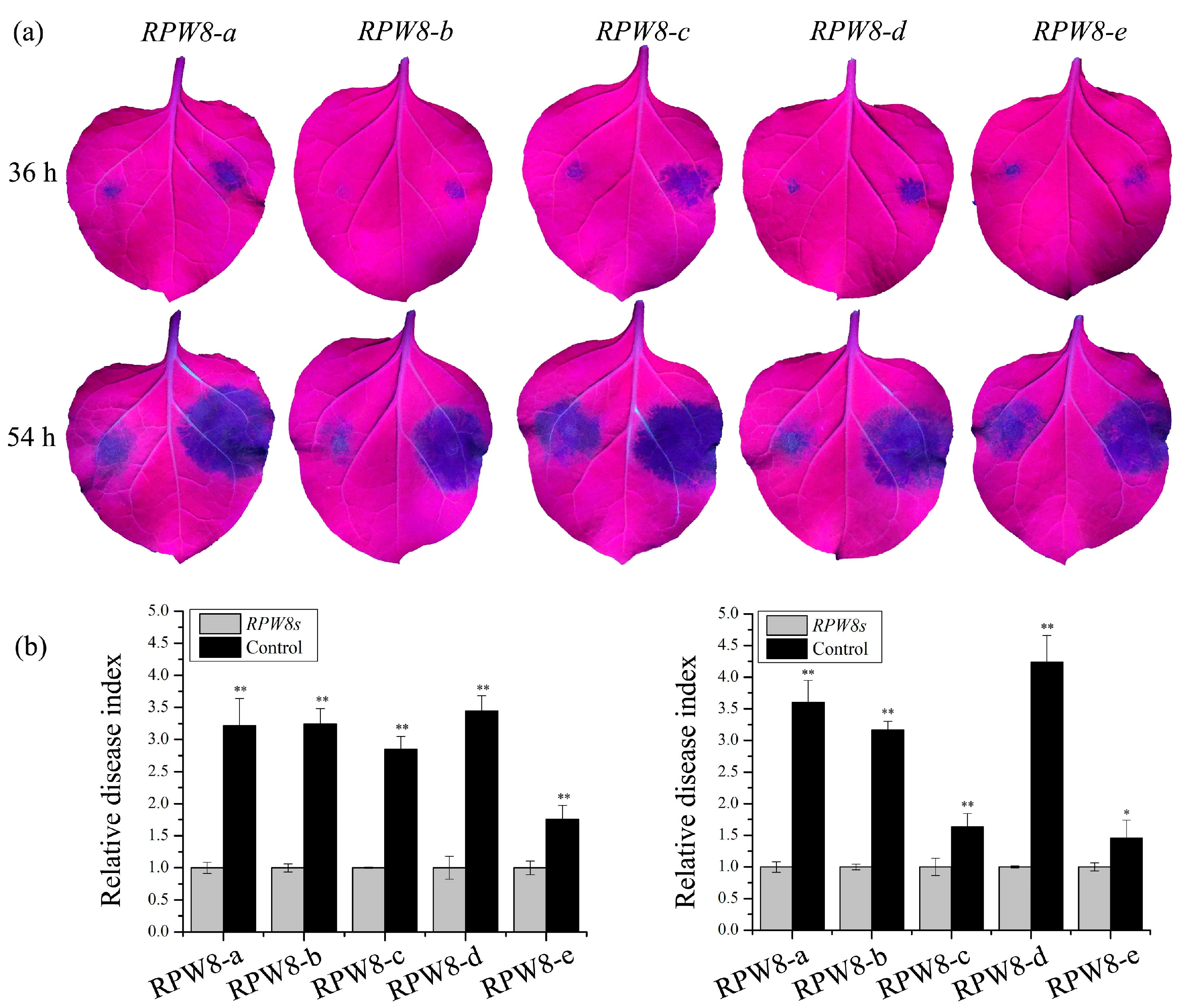
2.4. Transiently Expressing RPW8s Enhanced Resistance to Ph. capsici
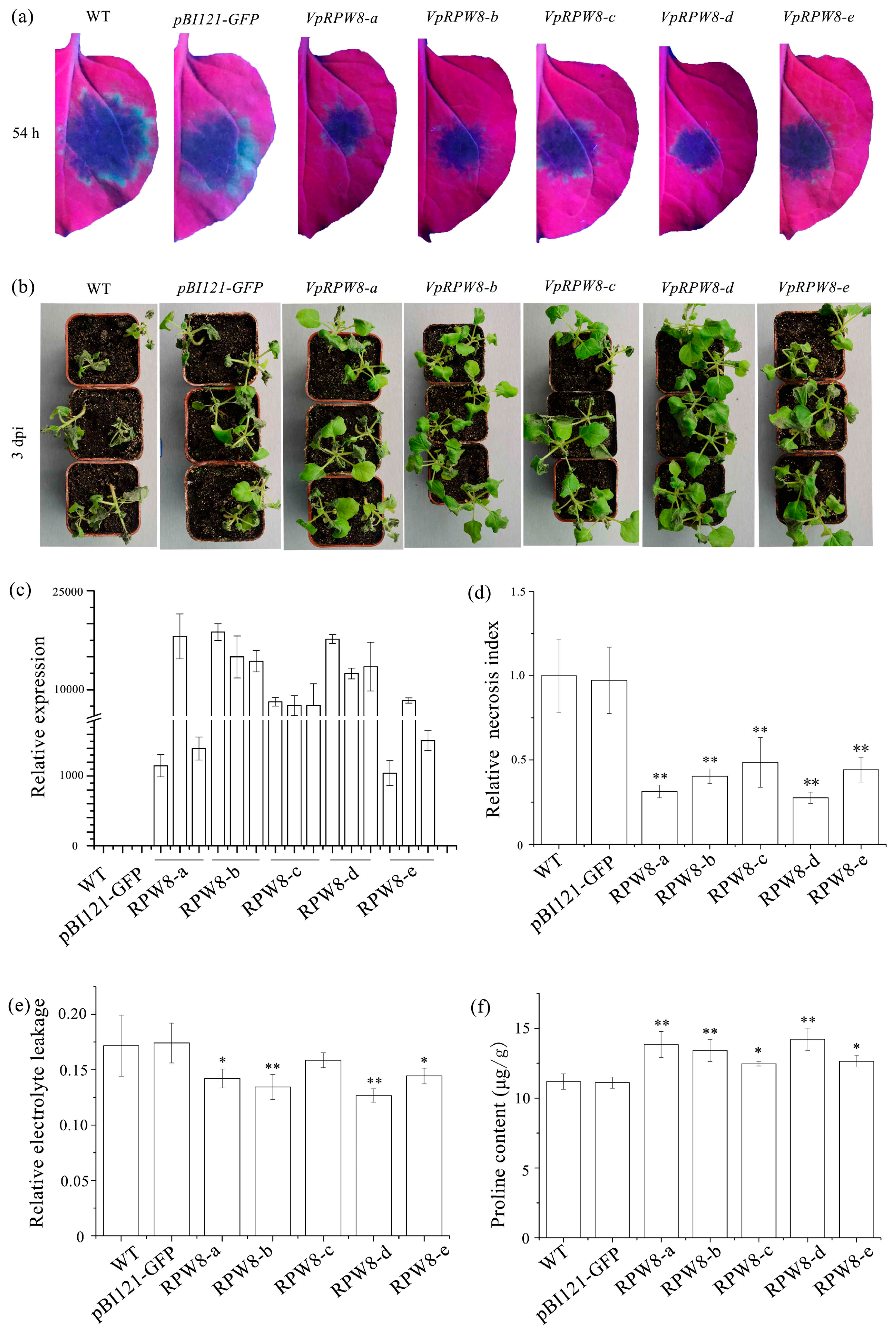
2.5. Stably Expressing RPW8s Enhanced Resistance to Ph. capsici
3. Discussion
3.1. Conserved Domains and Sequence Polymorphism of RPW8s
3.2. VpRPW8s Participate in Resistance to P. viticola
3.3. Broad-Spectrum Disease Resistance of RPW8
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Gene Cloning and Sequence Analysis
4.3. Quantitative Real-Time PCR Analysis
4.4. Prokaryotic Expression and Subcellular Localization Analysis
4.5. Transient Transformation and Resistance to Ph. capsici in N. benthamiana
4.6. Stable Transformation and Resistance to Ph. capsici in N. benthamiana
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| RPW8 | Resistance to Powdery Mildew8 |
| R gene | Resistance Gene |
| NCBI | National Center for Biotechnology Information Search database |
| qRT-PCR | Quantitative Real-Time PCR |
| GFP | Green Fluorescent Protein |
| ORF | Open Reading Frame |
| hpi | Hours Post-Inoculation |
| dpi | Days Post-Inoculation |
References
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.C.; Goheen, A.C. Compendium of Grape Diseases; APS Press: St. Paul, MO, USA, 1988; ISBN 0890540888. [Google Scholar]
- Yu, Y.; Zhang, Y.; Yin, L.; Lu, J. The mode of host resistance to Plasmopara viticola infection of grapevines. Phytopathology 2012, 102, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediter. 2011, 50, 3–44. [Google Scholar] [CrossRef]
- Kiefer, B.; Riemann, M.; Büche, C.; Kassemeyer, H.H.; Nick, P. The host guides morphogenesis and stomatal targeting in the grapevine pathogen Plasmopara viticola. Planta 2002, 215, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wei, J.; Zou, Y.; Xu, K.; Wang, Y.; Lu, C.; Xu, Y. Molecular characteristics and biochemical functions of VpPR10s from Vitis pseudoreticulata associated with biotic- and abiotic stresses. Int. J. Mol. Sci. 2014, 15, 19162–19182. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; He, P.; Chen, J.; Lamikanra, O.; Lu, J. Evaluation of foliar resistance to Uncinula necator in Chinese wild Vitis species. Vitis 1995, 34, 159–164. [Google Scholar] [CrossRef]
- Xu, W.; Yu, Y.; Zhou, Q.; Ding, J.H.; Dai, L.M.; Xie, X.Q.; Xu, Y.; Zhang, C.H.; Wang, Y.J. Expression pattern, genomic structure, and promoter analysis of the gene encoding stilbene synthase from Chinese wild Vitis pseudoreticulata. J. Exp. Bot. 2011, 62, 2745–2761. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Schwaninger, H.; He, P.; Wang, Y. Comparison of resistance to powdery mildew and downy mildew in Chinese wild grapes. Vitis 2007, 46, 132–136. [Google Scholar]
- Merdinoglu, D.; Wiedemann-Merdinoglu, S.; Coste, P.; Dumas, V.; Haetty, S.; Butterlin, G.; Greif, C.; Adam-Blondon, A.; Bouquet, A.; Pauquet, J. Genetic analysis of downy mildew resistance derived from Muscadinia rotundifolia. Acta Hortic. 2003, 603, 451–456. [Google Scholar] [CrossRef]
- Fischer, B.M.; Salakhutdinov, I.; Akkurt, M.; Eibach, R.; Edwards, K.J.; Töpfer, R.; Zyprian, E.M. Quantitative trait locus analysis of fungal disease resistance factors on a molecular map of grapevine. Theor. Appl. Genet. 2004, 108, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Welter, L.J.; Göktrk-Baydar, N.; Akkurt, M.; Maul, E.; Eibach, R.; Töpfer, R.; Zyprian, E.M. Genetic mapping and localization of quantitative trait loci affecting fungal disease resistance and leaf morphology in grapevine (Vitis vinifera L). Mol. Breed. 2007, 20, 359–374. [Google Scholar] [CrossRef]
- Bellin, D.; Peressotti, E.; Merdinoglu, D.; Wiedemann-Merdinoglu, S.; Adam-Blondon, A.-F.; Cipriani, G.; Morgante, M.; Testolin, R.; di Gaspero, G. Resistance to Plasmopara viticola in grapevine ‘Bianca’ is controlled by a major dominant gene causing localized necrosis at the infection site. Theor. Appl. Genet. 2009, 120, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Marguerit, E.; Boury, C.; Manicki, A.; Donnart, M.; Butterlin, G.; Némorin, A.; Wiedemann-Merdinoglu, S.; Merdinoglu, D.; Ollat, N.; Decroocq, S. Genetic dissection of sex determinism, inflorescence morphology and downy mildew resistance in grapevine. Theor. Appl. Genet. 2009, 118, 1261–1278. [Google Scholar] [CrossRef] [PubMed]
- Zyprian, E.; Welter, L.J.; Akkurt, M.; Ebert, S.; Salakhutdinov, I.; Göktürk-Baydar, N.; Eibach, R.; Töpfer, R. Genetic analysis of fungal disease resistance in grapevine. Acta Hortic. 2009, 827, 535–538. [Google Scholar] [CrossRef]
- Moreira, F.M.; Madini, A.; Marino, R.; Zulini, L.; Stefanini, M.; Velasco, R.; Kozma, P.; Grando, M.S. Genetic linkage maps of two interspecific grape crosses (Vitis spp.) used to localize quantitative trait loci for downy mildew resistance. Tree Genet. Genom. 2011, 7, 153–167. [Google Scholar] [CrossRef]
- Schwander, F.; Eibach, R.; Fechter, I.; Hausmann, L.; Zyprian, E.; Töpfer, R. Rpv10: A new locus from the Asian Vitis gene pool for pyramiding downy mildew resistance loci in grapevine. Theor. Appl. Genet. 2012, 124, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Gururani, M.A.; Venkatesh, J.; Upadhyaya, C.P.; Nookaraju, A.; Pandey, S.K.; Park, S.W. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant Pathol. 2012, 78, 51–65. [Google Scholar] [CrossRef]
- Nürnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate immunity in plants and animals: Striking similarities and obvious differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Ron, M.; Avni, A. The receptor for the fungal elicitor ethylene-inducing xylanase is a member of a resistance-like gene family in tomato. Plant Cell 2004, 16, 1604–1615. [Google Scholar] [CrossRef] [PubMed]
- Schwessinger, B.Z.C. News from the frontline: Recent insights into PAMP-triggered immunity in plants. Curr. Opin. Plant Biol. 2008, 11, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Van der Biezen, E.A.; Jones, J.D. Plant disease-resistance proteins and the gene-for-gene concept. Trend. Biochem. Sci. 1998, 23, 454–456. [Google Scholar] [CrossRef]
- Xiao, S.; Ellwood, S.; Calis, O.; Patrick, E.; Li, T.; Coleman, M.; Turner, J.G. Broad-spectrum mildew resistance in Arabidopsis thaliana mediated by RPW8. Science 2001, 291, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wen, Y.; Berkey, R.; Xiao, S. Specific targeting of the Arabidopsis resistance protein RPW8.2 to the interfacial membrane encasing the fungal haustorium renders broad-spectrum resistance to powdery mildew. Plant Cell 2009, 21, 2898–2913. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. The “Arabidopsis” RPW8 resistance protein is recruited to the extrahaustorial membrane of biotrophic powdery mildew fungi. Plant Cell 2009, 21, 2543. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Devoto, A.; Turner, J.G.; Xiao, S. Expression of the membrane-associated resistance protein RPW8 enhances basal defense against biotrophic pathogens. MPMI 2007, 20, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, J.; Sun, J.L.; Ma, X.F.; Wang, T.T.; Berkey, R.; Yang, H.; Niu, Y.Z.; Fan, J.; Li, Y.; et al. Multiple evolutionary events involved in maintaining homologs of resistance to powdery mildew 8 in Brassica napus. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.L. Polymorphism Analysis and Functional Identification of RPW8 Homologues in Brassica napus. Master’ Thesis, China Agricultural University, Beijing, China, June 2014. [Google Scholar]
- Henanff, G.L.; Heitz, T.; Mestre, P.; Mutterer, J.; Walter, B.; Chong, J. Characterization of Vitis vinifera NPR1 homologs involved in the regulation of pathogenesis-related gene expression. BMC Plant Biol. 2009, 9, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Kortekamp, A. Expression analysis of defence-related genes in grapevine leaves after inoculation with a host and a non-host pathogen. Plant Physiol. Biochem. 2006, 44, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, Y.; Zhang, H.; Huang, H.; Folta, K.M.; Lu, J. Whole genome wide expression profiles of Vitis amurensis grape responding to downy mildew by using Solexa sequencing technology. BMC Plant Biol. 2010, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.; Yang, B.; Tian, D.; Wu, L.; Wang, D.; Sreekala, C.; Yang, F.; Chu, Z.; Wang, G.L.; White, F.F. R gene expression induced by a type-III effector triggers disease resistance in rice. Nature 2005, 435, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Kortekamp, A.; Welter, L.; Vogt, S.; Knoll, A.; Schwander, F.; Töpfer, R.; Zyprian, E. Identification, isolation and characterization of a CC-NBS-LRR candidate disease resistance gene family in grapevine. Mol. Breed. 2008, 22, 421–432. [Google Scholar] [CrossRef]
- Radwan, O.; Gandhi, S.; Heesacker, A.; Whitaker, B.; Taylor, C.; Plocik, A.; Kesseli, R.; Kozik, A.; Michelmore, R.W.; Knapp, S.J. Genetic diversity and genomic distribution of homologs encoding NBS-LRR disease resistance proteins in sunflower. Mol. Genet. Genom. 2008, 280, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, J.; Yin, L.; Zhang, Y.; Qu, J.; Lu, J. Comparative transcriptome analysis reveals defense-related genes and pathways against downy mildew in Vitis amurensis grapevine. Plant Physiol. Biochem. 2015, 95, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.Y.; Charoenwattana, P.; Holcombe, L.; Turner, J.G. The Arabidopsis genes RPW8.1 and RPW8.2 confer induced resistance to powdery mildew diseases in tobacco. MPMI 2003, 16, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, F.; Sebastiana, M.; Pais, M.S.; Figueiredo, A. Reference gene selection and validation for the early responses to downy mildew infection in susceptible and resistant Vitis vinifera cultivars. PLoS ONE 2013, 8, e72998. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Li, X.; Wu, J.; Yin, L.; Zhang, Y.; Lu, J. Studying the mechanism of Plasmopara viticola RxLR effectors on suppressing plant immunity. Front. Microbiol. 2016, 7, 709. [Google Scholar] [CrossRef] [PubMed]
- Gallois, P.; Marinho, P. Leaf disk transformation using Agrobacterium tumefaciens-expression of heterologous genes in tobacco. Method Mol. Biol. 1995, 49, 39–48. [Google Scholar]
- Li, X.; Zhang, Y.; Yin, L.; Lu, J. Overexpression of pathogen-induced grapevine TIR-NB-LRR gene VaRGA1, enhances disease resistance and drought and salt tolerance in Nicotiana benthamiana. Protoplasma 2016, 254, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, J. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol. 2001, 42, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Z.; Wang, P.H.; Zhao, H.X. Determination of the content of free proline in wheat leaves. Plant Physiol. Commun. 1990, 4, 62–65. [Google Scholar]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, G.; Fu, P.; Liu, Y.; Xiang, J.; Lu, J. Molecular Characterization and Overexpression of VpRPW8s from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici in Nicotiana benthamiana. Int. J. Mol. Sci. 2018, 19, 839. https://doi.org/10.3390/ijms19030839
Lai G, Fu P, Liu Y, Xiang J, Lu J. Molecular Characterization and Overexpression of VpRPW8s from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici in Nicotiana benthamiana. International Journal of Molecular Sciences. 2018; 19(3):839. https://doi.org/10.3390/ijms19030839
Chicago/Turabian StyleLai, Gongti, Peining Fu, Yunxiao Liu, Jiang Xiang, and Jiang Lu. 2018. "Molecular Characterization and Overexpression of VpRPW8s from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici in Nicotiana benthamiana" International Journal of Molecular Sciences 19, no. 3: 839. https://doi.org/10.3390/ijms19030839
APA StyleLai, G., Fu, P., Liu, Y., Xiang, J., & Lu, J. (2018). Molecular Characterization and Overexpression of VpRPW8s from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici in Nicotiana benthamiana. International Journal of Molecular Sciences, 19(3), 839. https://doi.org/10.3390/ijms19030839