Transport and Use of Bicarbonate in Plants: Current Knowledge and Challenges Ahead

 ,
,  ,
,  ,
, 
Abstract
:
1. Introduction
2. Bicarbonate Transport
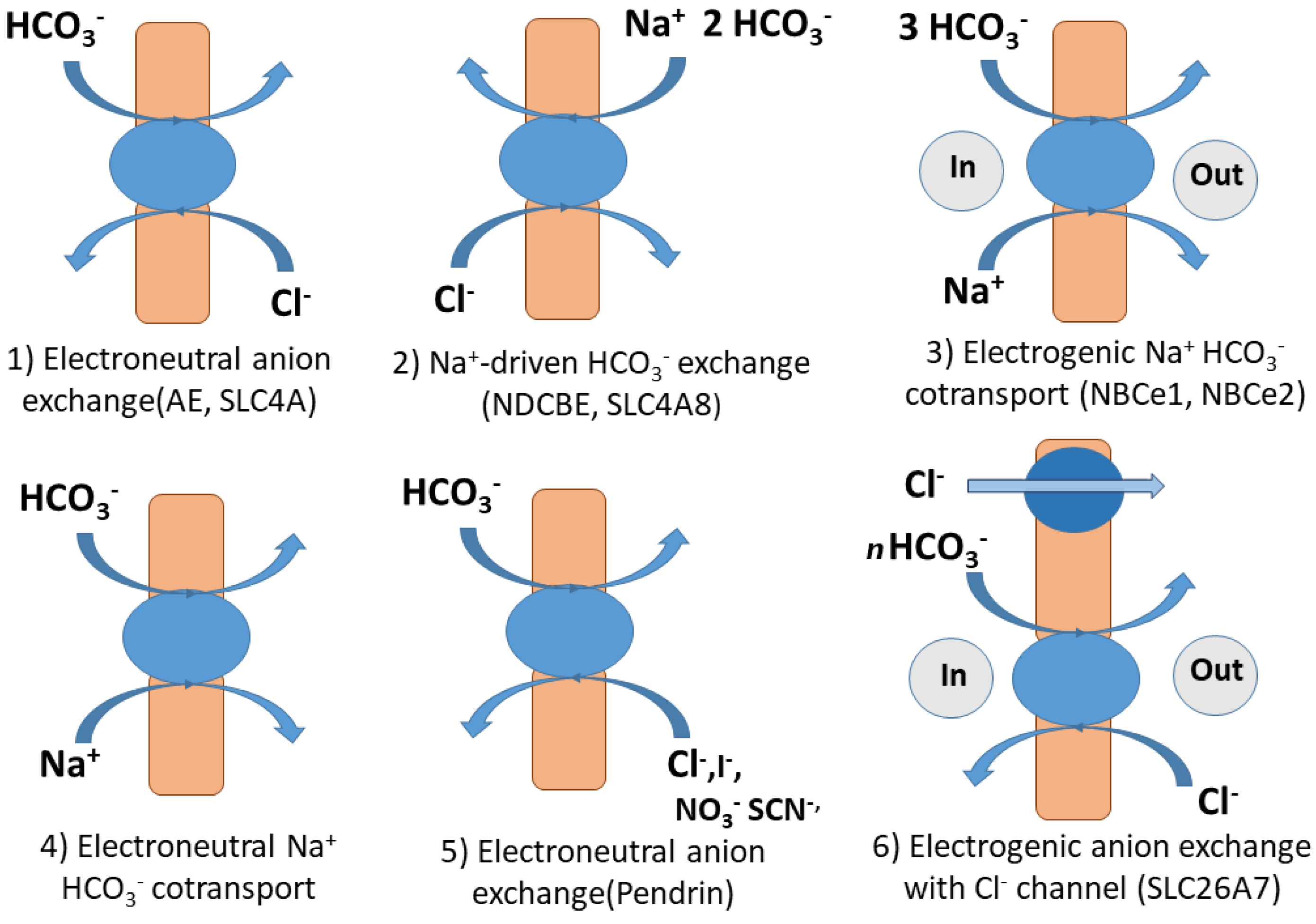
2.1. Bicarbonate Transport by Solute Carriers (SLC) in Humans and Mammals
2.2. Cinorg Transporters in Cyanobacteria
2.3. Cinorg Transport in Microalgae
2.4. Cinorg Transport in Macroalgae
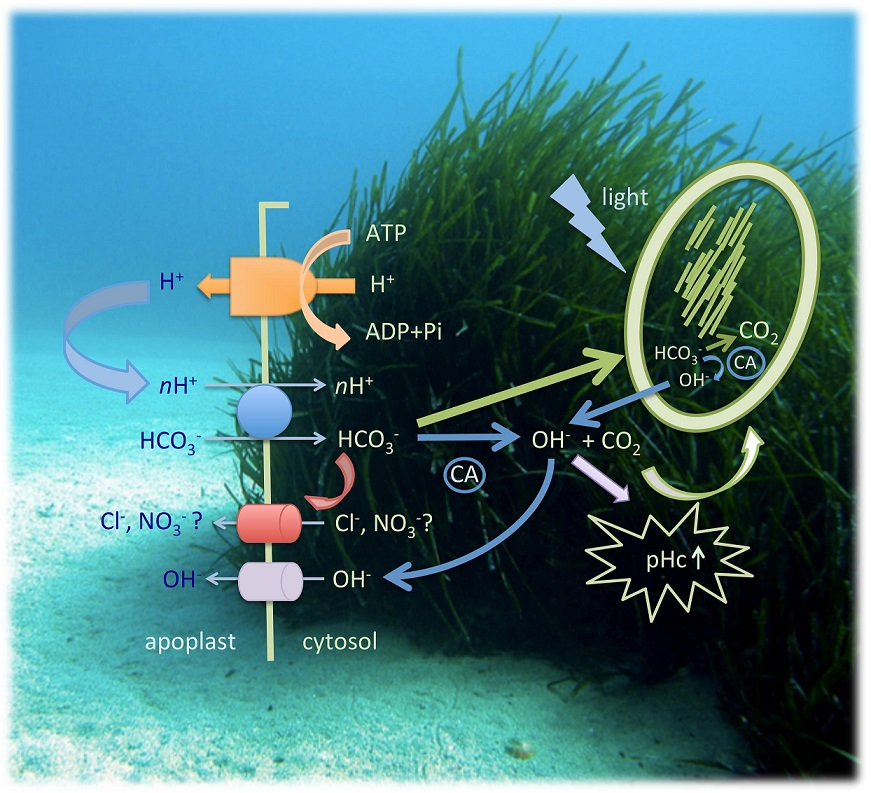
2.5. Cinorg Transport in Seagrasses
2.6. Cinorg Transport in Higher Land Plants
3. Formation and Use of Bicarbonate in Plants
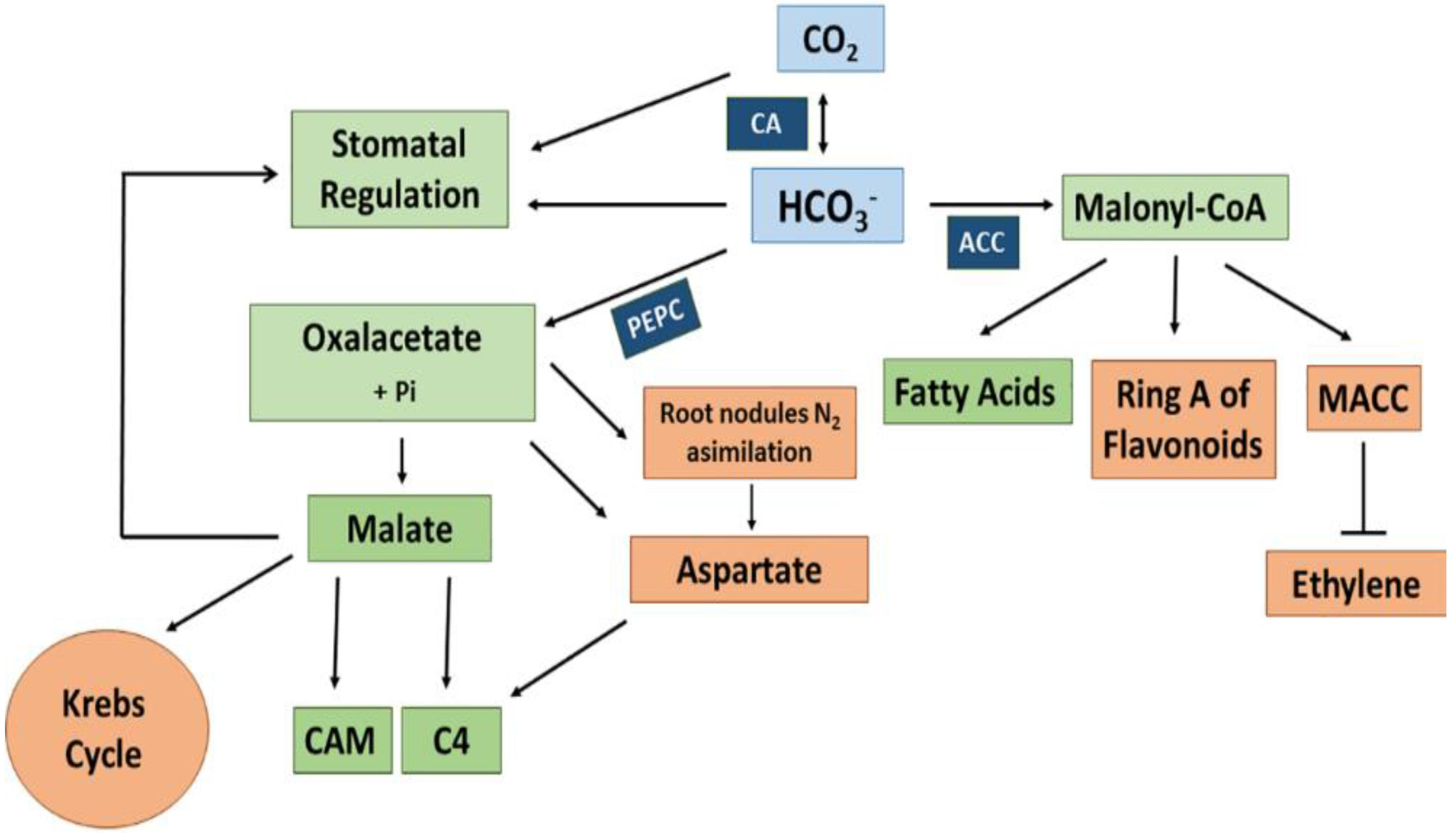
3.1. Plant Carbonic Anhydrase and Phosphoenolpyruvate Carboxylase
3.1.1. Plant α-Carbonic Anhydrases (αCA)
3.1.2. Plant β-Carbonic Anhydrases
3.1.3. Plant γ-Carbonic Anhydrases
3.1.4. Plant PEP Carboxylases
3.2. Carbon Concentration Mechanisms (CCM) in Terrestrial Plants
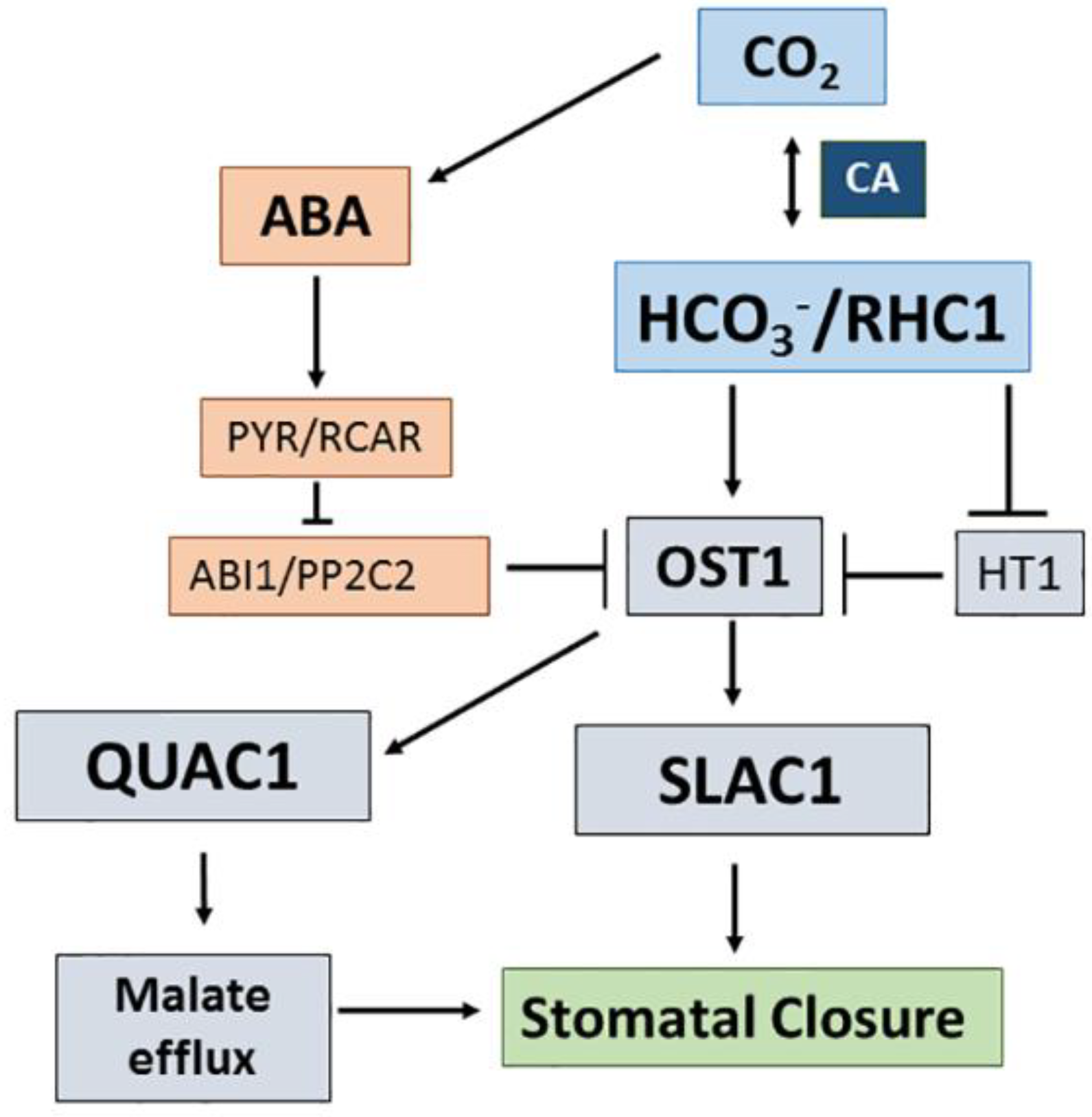
3.3. CO2/Bicarbonate Signalling in Stomatal Guard Cells
4. Plant Response to Bicarbonate-Rich Soils
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| CCM | Carbon concentration mechanism |
| CA | Carbonic anhydrase |
| PEPC | Phosphoenolpyruvate carboxylase |
| SLAC | Slow anion channel |
| SLC | Solute carriers |
| QAC | Quick anion channel |
References
- Madsen, T.V.; San-Jensen, K. Photosynthetic carbon assimilation in aquatic macrophytes. Aquat. Bot. 1991, 41, 5–40. [Google Scholar] [CrossRef]
- Keeley, J.E.; Sandquist, D.R. Carbon: Freshwater plants. Plant Cell Environ. 1992, 15, 1021–1035. [Google Scholar] [CrossRef]
- Falkowsky, P.G.; Raven, J.A. Aquatic Photosynthesis, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2007; Chapter 1; pp. 1–43. ISBN 9780691115511. [Google Scholar]
- Krauskopf, K.B.; Bird, D.K. Introduction to Geochemistry, 3rd ed.; McGraw Hill: New York, NY, USA, 1995; Chapter 3; pp. 61–85. ISBN 0-07-o35820-6. [Google Scholar]
- Bloom, P.R.; Inskeep, W.P. Factors affecting bicarbonate chemistry and iron chlorosis in soils. J. Plant Nutr. 1986, 9, 215–228. [Google Scholar] [CrossRef]
- Sauze, J.; Jones, S.P.; Wingate, L.; Wohl, S.; Ogée, J. The role of soil pH on soil carbonic anhydrase activity. Biogeosciences 2018, 597–612. [Google Scholar] [CrossRef]
- Jones, S.P.; Ogée, J.; Sauze, J.; Wohl, S.; Saavedra, N.; Fernández-Pardo, N.; Maire, J.; Launois, T.; Bosc, A.; Wingate, L. Non-destructive estimates of soil carbonic anhydrase activity and associated soil water oxygen isotope composition. Hydrol. Earth Syst. Sci. 2017, 21, 6363–6377. [Google Scholar] [CrossRef]
- Gao, K.S.; Zou, D.G. Photosynthetic bicarbonate utilization by a terrestrial cyanobacterium Nostoc flagelliforme (Cyanophyceae). J. Appl. Phycol. 2001, 37, 768–771. [Google Scholar] [CrossRef]
- Römheld, V. Different strategies for iron acquisition in higher plants. Physiol. Plant. 1987, 70, 231–234. [Google Scholar] [CrossRef]
- Rashid, A.; Ryan, J. Micronutrient constraints to crop production in soils with Mediterranean-type characteristics: A review. J. Plant Nutr. 2004, 27, 959–975. [Google Scholar] [CrossRef]
- Rao, S.; Wu, Y. Root derived bicarbonate assimilation in response to variable water deficit in Camptotheca acuminate seedlings. Photosynth. Res. 2017, 134, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Bouley, R.; Păunecu, T.G.; Breton, S.; Lu, H.A.J. New insights into the dynamic regulation of water and acid-base balance by renal epithelial cells. Am. J. Physiol. Cell Physiol. 2012, 302, C1421–C1433. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Paine, M.L. Bicarbonate transport during enamel maturation. Calcif. Tissue Int. 2017, 101, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Zoccola, D.; Ganot, P.; Bertucci, A.; Caminiti-Segonds, N.; Techer, N.; Voolstra, C.R.; Aranda, M.; Tambutté, E.; Allemand, D.; Casey, J.R.; Tambutté, S. Bicarbonate transporters in corals point towards a key step in the evolution of cnidarian calcification. Sci. Rep. 2015, 5, 9983. [Google Scholar] [CrossRef] [PubMed]
- Boron, W.F.W.F.; Endeward, V.; Gros, G.; Musa-Aziz, R.; Pohl, P. Intrinsic CO2 permeability of cell membranes and potential biological relevance of CO2 channels. ChemPhysChem 2011, 12, 1017–1019. [Google Scholar] [CrossRef] [PubMed]
- Talbot, K.; Kwong, R.W.M.; Gilmour, K.M.; Perry, S.F. The water channel aquaporin-1a1 facilitates movement of CO2 and ammonia in zebrafish (Danio rerio) larvae. J. Exp. Biol. 2015, 218, 3931–3940. [Google Scholar] [CrossRef] [PubMed]
- Endeward, V.; Arias-Hidalgo, M.; Al-Samir, S.; Gros, G. CO2 permeability of biological membranes and role of CO2 channels. Membranes 2017, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Missner, A.; Kügler, P.; Saparov, S.M.; Sommer, K.; Mathai, J.C.; Zeidel, M.L. Carbon dioxide transport through membranes. J. Biol. Chem. 2008, 283, 25340–25347. [Google Scholar] [CrossRef] [PubMed]
- Uehlein, N.; Kai, L.; Kaldenhoff, R. Plant aquaporins and CO2. In Plant Aquaporins; Signaling and Communications in Plants Series; Chaumont, F., Tyerman, S.D., Eds.; Springer Int. Publ.: Cham, Switzerland, 2017; pp. 255–265. [Google Scholar] [CrossRef]
- Flexas, J.; Ribas-Carbó, M.; Hanson, D.T.; Bota, J.; Otto, B.; Cifre, J.; McDowell, M.C.; Medrano, H.; Kaldenhoff, R. Tobacco aquaporin NTAQP1 is involved in mesophyll conductance to CO2 in vivo. Plant J. 2006, 48, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Groszmann, M.; Osborn, H.L.; Evans, J.R. Carbon dioxide and water transport through aquaporins. Plant Cell Environ. 2017, 40, 938–961. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A.; Beardall, J.; Sánchez-Baracaldo, P. The possible evolution and future of CO2-concentrating mechanisms. J. Exp. Bot. 2017, 68, 3701–3716. [Google Scholar] [CrossRef] [PubMed]
- Price, G.D.; Shelden, M.C.; Howitt, S.M. Membrane topology of the cyanobacterial bicarbonate transporter, SbtA, and identification of potential regulatory loops. Mol. Membr. Biol. 2011, 28, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Duanmu, D.; Miller, A.R.; Horken, K.M.; Weeks, D.P.; Spalding, M.H. Knockdown of limiting-CO2-induced gene HLA3 decreases HCO3− transport and photosynthetic Ci affinity in Chlamydomonas reinhardtii. Proc. Nat. Acad. Sci. USA 2009, 106, 5990–5995. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Tanaka, A.; Matsuda, Y. SLC4 family transporters in a marine diatom directly pump bicarbonate from seawater. Proc. Natl. Acad. Sci. USA 2013, 110, 1767–1772. [Google Scholar] [CrossRef] [PubMed]
- Fernández, P.A.; Hurd, C.L.; Roleda, M.Y. Bicarbonate uptake via an anion exchange protein is the main mechanism of inorganic carbon acquisition by the giant kelp Macrocystis pyrifera (Laminariales, Phaeophyceae) under variable pH. J. Phycol. 2014, 50, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; García, D.; García-Sánchez, M.J.; Niell, F.X.; Felle, H.H.; Fernández, J.A. Direct uptake of HCO3− in the marine angiosperm Posidonia oceanica (L.) Delile driven by a plasma membrane H+ economy. Plant Cell Environ. 2017, 40, 2820–2830. [Google Scholar] [CrossRef] [PubMed]
- Frachisse, J.M.; Thomine, S.; Colcombet, J.; Guern, J.; Barbier-Brygoo, H. Sulfate is both a substrate and an activator of the voltage-dependent anion channel of Arabidopsis hypocotyl cells. Plant Physiol. 1999, 121, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.K. Plasma membrane anion channels in higher plants and their putative functions in roots. New Phytol. 2006, 169, 647–666. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Spalding, M.J. Acclimation to very low CO2: Contribution of limiting CO2 inducible proteins, LCIb and LCIA, to inorganic carbon uptake in Chlamydomonas reinhardtii. Plant Physiol. 2014, 166, 2040–2050. [Google Scholar] [CrossRef] [PubMed]
- Yamano, T.; Sato, E.; Iguchi, H.; Fukuda, Y.; Fukuzawa, H. Characterization of cooperative bicarbonate uptake into chloroplast stroma in the green alga Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2015, 112, 7315–7320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungnick, N.; Ma, Y.; Mukherjee, B.; Cronan, J.C.; Speed, D.J.; Laborde, S.M.; Longstreth, D.J.; Moroney, J.V. The carbon concentrating mechanism in Chlamydomonas reinhardtii: Finding the missing pieces. Photosynth. Res. 2014, 121, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A.; Beardall, J.; Giordano, M. Energy costs of carbon dioxide concentrating mechanisms in aquatic organisms. Photosynth. Res. 2014, 121, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.D.; Boron, W.F. The divergence, actions, roles, and relatives of sodium-coupled bicarbonate transporters. Physiol. Rev. 2013, 93, 803–959. [Google Scholar] [CrossRef] [PubMed]
- Alka, K.; Casey, J.R. Bicarbonate transport in health and disease. Int. Union Biochem. Mol. Biol. 2014, 66, 596–615. [Google Scholar] [CrossRef] [PubMed]
- Perland, E.; Bagchi, S.; Klaesson, A.; Fredriksson, R. Characteristics of 29 novel atypical solute carriers of major facilitator superfamily type: Evolutionary conservation, predicted structure and neuronal co-expression. Open Biol. 2017, 7, 170142. [Google Scholar] [CrossRef] [PubMed]
- Alper, S.L.; Sharmer, A.K. The SLC26 gene family of anion transporters and channels. Mol. Aspects Med. 2013, 34, 494–515. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.F.; Chen, A.-P.; Parler, D.; Boron, W.F. The SLC4 family of bicarbonate (HCO3−) transporters. Mol. Aspects Med. 2013, 34, 159–182. [Google Scholar] [CrossRef] [PubMed]
- Bonar, P.T.; Casey, J.R. Plasma membrane Cl−/HCO3− exchangers: Structure, mechanism and physiology. Channels 2008, 2, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Aalkjaer, C.; Boedtkjer, E.; Choi, I.; Lee, S. Cation-coupled bicarbonate transporters. Compr. Physiol. 2014, 4, 1605–1637. [Google Scholar] [CrossRef] [PubMed]
- Alvadia, C.M.; Sommer, T.; Bjerregaard-Andersen, K.; Damkier, H.H.; Montrasio, M.; Aalkjaer, C.; Morth, J.P. The crystal structure of the regulatory domain of the human sodium-driven chloride/bicarbonate exchanger. Sci. Rep. 2017, 7, 12131. [Google Scholar] [CrossRef] [PubMed]
- Omata, T.; Price, G.D.; Badger, M.R.; Okamura, M.; Gohta, S.; Ogawa, T. Identification of an ATP-binding cassette transporter involved in bicarbonate uptake in the cyanobacterium Synechococcus sp. Strain PCC7942. Proc. Natl. Acad. Sci. USA 1999, 23, 13571–13576. [Google Scholar] [CrossRef]
- Price, G.D.; Badger, M.R.; Woodger, F.J.; Long, B.M. Advances in understanding the cyanobacterial CO2-concentrating mechanism (CCM): Functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. J. Exp. Bot. 2008, 59, 1441–1461. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Price, G.D.; Badger, M.R.; Enomoto, C.; Omata, T. Bicarbonate binding activity of the CmpA protein of the cyanobacterium Synechococcus sp. strain PCC7942 involved in active transport of bicarbonate. J. Biol. Chem. 2000, 275, 20551–20555. [Google Scholar] [CrossRef] [PubMed]
- Koropatkin, N.M.; Koppenaal, D.W.; Pakrasi, H.B.; Smith, T.J. The structure of a cyanobacterial bicarbonate transport protein, CmpA. J. Biol. Chem. 2007, 282, 2606–2614. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.I.; Omata, T. Substrate-binding lipoprotein of the cyanobacterium Synechococcus sp. strain PCC7942 involved in the transport of nitrate and nitrite. J. Biol. Chem. 1997, 272, 3036–3041. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Katoh, H.; Sonoda, M.; Ohkawa, H.; Shimoyama, M.; Fukuzawa, H.; Kaplan, A.; Ogawa, T. Genes essential to sodium-dependent bicarbonate transport in cyanobacteria: Function and phylogenetic analysis. J. Biol. Chem. 2002, 277, 18658–18664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.P.; Battchikova, N.; Jansen, T.; Appel, J.; Ogawa, T.; Aro, E.M. Expression and functional roles of the two distinct NDH-1 complexes and the carbon acquisition complex NdhD3/NdhF3/CupA/Sll1735 in Synechocystis sp. PCC 6803. Plant Cell 2004, 16, 3326–3340. [Google Scholar] [CrossRef] [PubMed]
- Sültemeyer, D.; Klughammer, B.; Badger, M.R.; Price, G.D. Fast induction of high-affinity HCO3− transport in cyanobacteria. Plant Physiol. 1998, 116, 183–192. [Google Scholar] [CrossRef]
- Amoroso, G.; Seimetz, N.; Sultemeyer, D. The dc13 gene upstream of ictB is involved in rapid induction of the high affinity Na+ dependent HCO3− transporter in cyanobacteria. Photosynth. Res. 2003, 77, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Price, G.D.; Woodger, F.J.; Badger, M.R.; Howitt, S.M.; Tucker, L. Identification of a SulP-type bicarbonate transporter in marine cyanobacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 18228–18233. [Google Scholar] [CrossRef] [PubMed]
- Shibagaki, N.; Grossman, A.R. Probing the function of STAS domains of the Arabidopsis sulfate transporters. J. Biol. Chem. 2004, 279, 30791–30799. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Ohkawa, H.; Kaneko, T.; Fukuzawa, H.; Tabata, S.; Kaplan, A.; Ogawa, T. Distinct constitutive and low CO2 induced CO2 uptake systems in cyanobacteria: Genes involved and their phylogenetic relationship with homologous genes in other organisms. Proc. Natl. Acad. Sci. USA 2001, 98, 11789–11794. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Badger, M.R.; Price, G.D. Novel gene products associated with NdhD3/D4-containing NDH-1 complexes are involved in photosynthetic CO2 hydration in the cyanobacterium, Synechococcus sp. PCC7942. Mol. Microbiol. 2002, 43, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Stessman, D.J.; Spalding, M.H. The CO2 concentrating mechanism and photosynthetic carbon assimilation in limiting CO2: How Chlamydomonas works against the gradient. Plant J. 2015, 82, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Burow, M.D.; Chen, Z.Y.; Mouton, T.M.; Moroney, J.V. Isolation of cDNA clones of genes induced upon transfer of Chlamydomonas reinhardtii cells to low CO2. Plant Mol. Biol. 1996, 31, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, N.; Mukherjee, B.; Tsujikawa, T.; Yanase, M.; Nakano, H.; Moroney, J.V.; Fukuzawa, H. Expression of a low CO2-inducible protein, LCI1, increases inorganic carbon uptake in the green alga Chlamydomonas reinhardtii. Plant Cell 2010, 22, 3105–3117. [Google Scholar] [CrossRef] [PubMed]
- Pollock, S.V.; Prout, D.L.; Godfrey, A.C.; Lemaire, S.D.; Moroney, J.V. The Chlamydomonas reinhardtii proteins Ccp1 and Ccp2 are required for long-term growth, but are not necessary for efficient photosynthesis, in a low CO2 environment. Plant Mol. Biol. 2004, 56, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Tortell, P.D.; Reinfelder, J.R.; Morel, F.MM. Active uptake of bicarbonate by diatoms. Nature 1997, 390, 243–244. [Google Scholar] [CrossRef]
- Hopkinson, B.M.; Dupont, C.L.; Matsuda, Y. The physiology and genetics of CO2 concentrating mechanisms in model diatoms. Curr. Opin. Plant Biol. 2016, 31, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Dupont, C.L.; Hopkins, B.M. The diversity of CO2-concetrationg mechanisms in marine diatoms as inferred from their genetic content. J. Exp. Bot. 2017, 68, 3937–3948. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, S.P. The chloroplast endoplasmic reticulum: Structure, function and evolutionary significance. Int. Rev. Cytol. 1981, 72, 49–99. [Google Scholar] [CrossRef]
- Huertas, I.E.; Lubián, L.M. Comparative study of dissolved inorganic carbon and photosynthetic responses in Nannochloris (Chlorophyceae) and Nannochloropsis (Eustigmatophyceae) species. Can. J. Bot. 1997, 76, 1104–1108. [Google Scholar] [CrossRef]
- Huertas, I.E.; Espie, G.S.; Colman, B.; Lubián, L.M. Light-dependent bicarbonate uptake and CO2 efflux in the microalga Nannochloropsis gaditana. Planta 2000, 211, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Huertas, I.E.; Lubián, L.M.; Espie, G.S. Mitochondrial-driven bicarbonate transport supports photosynthesis in a marine microalga. Plant Physiol. 2002, 130, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Herfort, L.; Thake, B.; Roberts, J. Acquisition and use of bicarbonate by Emiliania huxleyi. New Phytol. 2002, 156, 427–436. [Google Scholar] [CrossRef]
- Bruce, L.J.; Robinson, H.C.; Guizouarn, H.; Borgese, F.; Harrison, P.; King, M.J.; Goede, J.S.; Coles, S.E.; Gore, D.M.; Lutz, H.U.; et al. Monovalent cation leaks in human red cells caused by single amino-acid substitutions in the transport domain of the band 3 chloride-bicarbonate exchanger, AE1. Nat. Genet. 2005, 37, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Young, E.B.; Beardall, J.; Giordano, M. Investigation of inorganic carbon acquisition by Dunaliella tertiolecta (Chlorophyta) using inhibitors of putative HCO3− utilization pathways. Eur. J. Phycol 2001, 36, 81–88. [Google Scholar] [CrossRef]
- Walker, N.A.; Smith, F.A.; Cathers, I.R. Bicarbonate assimilation by freshwater charophytes and higher plants. I. Membrane transport of bicarbonate is not proven. J. Membr. Biol. 1980, 12, 241–256. [Google Scholar]
- Price, G.D.; Badger, M.R.; Bassatt, M.E.; Whitecross, M.I. Involvement of plasmalemmasomes and carbonic anhydrase in photosynthetic utilization of bicarbonate in Chara corallina. Aust. J. Plant Physiol. 1985, 12, 241–256. [Google Scholar] [CrossRef]
- Beilby, M.J.; Bisson, M.A. pH banding in charophyte algae. In Plant Electrophysiological Methods and Cell Electrophysiology; Volkov, A.G., Ed.; Springer: Berlin, Germany, 2012; pp. 247–271. ISBN 978-3-642-29118-0. [Google Scholar]
- Lucas, W.J.; Nuccitelli, R. HCO3− and OH− transport across the plasmalemma of Chara. Planta 1980, 150, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Maberly, S.C.; Madsen, T.V. Freshwater agiosperm carbon concentration mechanisms: Processes and patterns. Funct. Plant Biol. 2002, 29, 393–405. [Google Scholar] [CrossRef]
- Maberly, S.C. Exogenous sources of inorganic carbon for photosynthesis by marine macroalgae. J. Phycol. 1990, 26, 439–449. [Google Scholar] [CrossRef]
- Koch, M.; Bowes, G.; Ross, C.; Zhang, X.-H. Climate change and ocean acidification effects on seagrasses and marine microalgae. Glob. Chang. Biol. 2013, 19, 103–132. [Google Scholar] [CrossRef] [PubMed]
- Cornwall, C.E.; Revill, A.T.; Hall-Spencer, J.M.; Milazzo, M.; Raven, J.A.; Hurd, C.L. Inorganic carbon physiology underpins macroalgal responses to elevated CO2. Sci. Rep. 2017, 7, 46297. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.G.; Bidwell, R.G.S. Mechanism of photosynthetic carbon dioxide uptake by the red macroalga, Chondrus crispus. Plant Physiol. 1989, 89, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Mercado, J.M.; Niell, F.X.; Figueroa, F.L. Regulation of the mechanism for HCO3− use by the inorganic carbon level in Porphyra leucosticta in Le Jolis (Rhotophyta). Planta 1997, 201, 319–325. [Google Scholar] [PubMed]
- Moulin, P.; Andría, J.R.; Axelsson, L.; Mercado, J.M. Different mechanisms of inorganic carbon acquisition in red macroalgae (Rhodophyta) revealed by the use of TRIS buffer. Aquat. Bot. 2011, 95, 31–38. [Google Scholar] [CrossRef]
- Flores-Moya, A.; Fernández, J.A. The role of external carbonic anhydrase in the photosynthetic use of inorganic carbon in the deep-water alga Phyllariopsis purpurascens (Laminariales, Phaeophyta). Planta 1998, 207, 115–119. [Google Scholar] [CrossRef]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Ann. Rev. Plant Biol. 2005, 6, 99–131. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A.; Hurd, C.J. Ecophysiology of photosynthesis in macroalgae. Photosynth. Res. 2012, 113, 105–125. [Google Scholar] [CrossRef] [PubMed]
- Larsson, C.; Axelsson, L. Bicarbonate uptake and utilization in marine macroalgae. Eur. J. Phycol. 2010, 34, 79–86. [Google Scholar] [CrossRef]
- Larsson, C.; Axelsson, L.; Ryberg, H.; Beer, S. Photosynthetic carbon utilization by Enteromorpha intestinalis (Chlorophyta) from a Swedish rockpool. Eur. J. Phycol. 1997, 32, 49–54. [Google Scholar] [CrossRef]
- Granbon, M.; Pedersen, M. Carbon acquisition strategies of the red alga Eucheuma denticulatum. Hydrobiologia 1999, 399, 349–354. [Google Scholar] [CrossRef]
- Andría, J.R.; Peérez-Lloréns, J.L.; Vergara, J.J. Mechanisms of inorganic carbon acquisition in Gracilaria gaditana nom. prov. (Rhodophyta). Planta 1999, 208, 564–573. [Google Scholar] [CrossRef]
- Johnston, A.M.; Maberly, S.C.; Raven, J.A. The acquisition of inorganic carbon by four red algae. Oecologia 1992, 92, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Beer, S.; Bjork, M.; Hellblom, F.; Axelsson, L. Inorganic carbon utilization by marine angiosperms (seagrasses). Funct. Plant Biol. 2002, 29, 349–354. [Google Scholar] [CrossRef]
- Borum, J.; Pedersen, O.; Kotula, L.; Fraser, M.W.; Statton, J.; Colmer, T.D.; Kendrick, G.A. Photosynthetic response to globally increasing CO2 of co-occurring temperate seagrass species. Plant Cell Environ. 2016, 39, 1240–1250. [Google Scholar] [CrossRef] [PubMed]
- Larkum, A.W.D.; Drew, E.A.; Ralph, P.J. Photosynthesis and metabolism in seagrasses at the cellular level. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, J.J., Duarte, C.A., Eds.; Springer: Berlin, Germany, 2006; pp. 323–345. ISBN 978-1-4020-2983-7. [Google Scholar]
- Larkum, A.W.D.; Davey, P.A.; Kuo, J.; Ralph, P.J.; Raven, J.A. Carbon-concentrating mechanisms in seagrasses. J. Exp. Bot. 2017, 68, 3773–3784. [Google Scholar] [CrossRef] [PubMed]
- Barbier-Brygoo, H.; Vinauger, M.; Colcombet, J.; Ephritikhine, G.; Frachisse, J.M.; Maurel, C. Anion channels in higher plants: Functional characterization, molecular structure and physiological role. Biochim. Biophy. Acta Biomembr. 2000, 1465, 199–218. [Google Scholar] [CrossRef]
- Beer, S.; Rehnberg, J. The acquisition of inorganic carbon by the seagrass Zostera marina. Aquat. Bot. 1997, 56, 277–283. [Google Scholar] [CrossRef]
- Hellblom, F.; Beer, S.; Björk, M.; Axelsson, L. A buffer sensitive inorganic carbon utilisation system in Zostera marina. Aquat. Bot. 2001, 69, 55–62. [Google Scholar] [CrossRef]
- Mercado, J.M.; Niell, F.X.; Silva, J.; Santos, R. Use of light and inorganic carbon acquisition by two morphotypes of Zostera noltii Hornem. J. Exp. Mar. Biol. Ecol. 2003, 297, 71–81. [Google Scholar] [CrossRef]
- Invers, O.; Perez, M.; Romero, J. Bicarbonate utilization in seagrass photosynthesis: Role of carbonic anhydrase in Posidonia oceanica (L.) Delile and Cymodocea nodosa (Ucria) Ascherson. J. Exp. Mar. Biol. Ecol. 1999, 235, 125–133. [Google Scholar] [CrossRef]
- Hellblom, F.; Axelsson, L. External HCO3− dehydration maintained by acid zones in the plasma membrane is an important component of the photosynthetic carbon uptake in Ruppia cirrhosa. Photosynth. Res. 2003, 77, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Uku, J.; Beer, S.; Björk, S. Buffer sensitivity of photosynthetic carbon utilization in eight tropical seagrasses. Mar. Biol. 2005, 147, 1085–1090. [Google Scholar] [CrossRef]
- Fernández, J.A.; García-Sández, M.J.; Felle, H.H. Physiological evidence for a proton pump and sodium exclusion mechanisms at the plasma membrane of the marine angiosperm Zostera marina L. J. Exp. Bot. 1999, 50, 1763–1768. [Google Scholar] [CrossRef]
- Rubio, L.; Belver, A.; Venema, K.; García-Sánchez, M.J.; Fernández, J.A. Evidence for a sodium efflux mechanism in the leaf cells of the seagrass Zostera marina L. J. Exp. Mar. Biol. Ecol. 2011, 402, 56–64. [Google Scholar] [CrossRef]
- Rubio, L.; Linares-Rueda, A.; García-Sánchez, M.J.; Fernández, J.A. Physiological evidence for a sodium-dependent high-affinity phosphate and nitrate transport at the plasma membrane of leaf and root cells of Zostera marina L. J. Exp. Bot. 2005, 56, 613–622. [Google Scholar] [CrossRef] [PubMed]
- James, P.L.; Larkum, A.W.D. Photosynthetic Ci acquisition of Posidonia australis. Aq. Bot. 1996, 55, 149–157. [Google Scholar]
- Olsen, J.L.; Rouzé, P.; Verhelst, B.; Lin, Y.-C.; Bayer, T.; Collen, J.; Dattolo, E.; de Paoli, E.; Dittami, S.; Maumus, F.; et al. The genome of the seagrass Zostera marina reveals angiosperm adaptation to the sea. Nature 2016, 530, 331–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Thurtle Schmidt, B.; Stroud, R.M. Structure of Bor1 supports an elevator transport mechanism for SLC4 anion exchangers. Proc. Natl. Acad. Sci. USA 2016, 113, 10542–10546. [Google Scholar] [CrossRef] [PubMed]
- Takano, J.; Tanaka, M.; Toyoda, A.; Miwa, K.; Kasai, K.; Fuji, K.; Onouchi, H.; Naito, S.; Fujiwara, T. Polar localization and degradation of Arabidopsis boron transporters through distinct trafficking pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 5220–5225. [Google Scholar] [CrossRef] [PubMed]
- Takano, J.; Miwa, K.; Fujiwara, T. Boron transport mechanisms: Collaboration of channels and transporters. Trends Plant Sci. 2008, 13, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Al Mansouri, H.M.; Alhendawi, R.A.M. Effect of increasing concentration of bicarbonate on plant growth and nutrient uptake by maize plants. Am.-Eur. J. Agric. Environ. Sci. 2014, 14, 1–6. [Google Scholar] [CrossRef]
- Meyer, S.; Mumm, P.; Imes, D.; Endler, A.; Weder, B.; Al-Rasheid, K.A.; Geiger, D.; Marten, I.; Martinoia, E.; Hedrich, R. AtALMT12 represents an R-type anion channel required for stomatal movement in Arabidopsis guard cells. Plant J. 2010, 63, 1054–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dryer, I.; Gomez-Porras, J.L.; Riaño-Pachón, D.M.; Hedrich, R.; Geiger, D. Molecular evolution of slow and quick anion channels (SLACs and QUACs/ALMTs). Front. Plant Sci. 2012, 3, 263. [Google Scholar] [CrossRef] [PubMed]
- Machelska, F.; Ahmad, A.; Batool, S.; Müller, H.M.; Ludwig-Müller, J.; Kreuzwieser, J.; Randewig, D.; Hänsch, R.; Mendel, R.R.; Hell, R.; et al. Drought enhanced xylem sap sulfate closes stomata affecting ALMT12 and guard cell ABA synthesis. Plant Physiol. 2017, 174, 798–814. [Google Scholar] [CrossRef]
- Planes, M.D.; Niñoles, R.; Rubio, L.; Bissoli, G.; Bueso, E.; García-Sánchez, M.J.; Alejandro, S.; Gonzalez-Guzmán, M.; Hedrich, R.; Rodriguez, P.L.; et al. A mechanism of growth inhibition by abscisic acid in germinating seeds of Arabidopsis thaliana based on inhibition of plasma membrane H+-ATPase and decreased cytosolic pH, K+, and anions. J. Exp. Bot. 2015, 66, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Canales, J.; Henriquez-Valencia, C.; Brauchi, S. The integration of electrical signals originating in the root of vascular plants. Front. Plant Sci. 2017, 8, 2173. [Google Scholar] [CrossRef] [PubMed]
- Tateda, C.; Watanabe, K.; Kusano, T.; Takahashi, Y. Molecular and genetic characterization of the gene family encoding the voltage-dependent anion channel in Arabidopsis. J. Exp. Bot. 2011, 4773–4785. [Google Scholar] [CrossRef] [PubMed]
- Wandrey, M.; Trevaski, B.; Brewin, N.; Udvardi, M.K. Molecular and cell biology of a family of voltage dependent anion channel porins in Lotus japonicus. Plant Physiol. 2004, 134, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Krammer, E.-M.; Homblé, F.; Prévost, M. Concentration dependent ion selectivity in VDAC: A molecular dynamics simulation study. PLoS ONE 2011, 6, e27994. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.J.; Miklavcic, S.J. A comprehensive biophysical model of ion and water transport in plant roots. I. Clarifying the roles of endodermal barriers in the salt stress response. Front. Plant Sci. 2017, 8, 1326. [Google Scholar] [CrossRef] [PubMed]
- Van der Vliet, L.; Peterson, C.; Hale, B. Cd accumulation in roots and shoots of durum wheat: The roles of transpiration rate and apoplastic bypass. J. Exp. Bot. 2007, 58, 2939–2947. [Google Scholar] [CrossRef] [PubMed]
- Wallace, A.; Mueller, R.T.; Wood, R.A.; Soufi, S.M. Plant uptake of bicarbonate as measured with the 11C isotope. Plant Soil 1979, 51, 431–435. [Google Scholar] [CrossRef]
- Vapaabuori, E.M.; Pelkonen, P. HCO3− uptake through the roots and its effect on the productivity of willow cuttings. Plant Cell Environ. 1985, 8, 531–534. [Google Scholar] [CrossRef]
- Vourinen, A.H.; Vapaavuori, E.M.; Lapinjoki, S. Time course of uptake of dissolved inorganic carbon through willow roots in light and in darkness. Physiol. Plant. 1989, 77, 33–38. [Google Scholar] [CrossRef]
- Overstreet, R.; Ruben, S.; Broyer, T.C. The absorption of bicarbonate ion by barley plants as indicated by studies with radioactive carbon. Proc. Natl. Acad. Sci. USA 1940, 26, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Stolwijk, J.A.J.; Thimann, K.V. On the utake of carbon dioxide and bicarbonate by roots, and its influence on growth. Plant Physiol. 1957, 32, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Stringer, J.W.; Kimmerer, T.W. Refixation of xylem sap CO2 in Populus deltoides. Physiol. Plant. 1993, 89, 243–251. [Google Scholar] [CrossRef]
- Nikolic, M.; Römheld, V. The role of leaf apoplast in iron nutrition of plants. In Plant Nutrition—Food Security and Sustainability of Agro-Ecosystems; Horst, W.J., Schenk, M.K., Bürkert, A., Claassen, N., Flessa, H., Frommer, W.B., Goldbach, H., Olfs, H.-W., Römheld, V., Sattelmacher, B., et al., Eds.; Kluwer Academic Publ.: Dordrecht, The Netherlands, 2001; pp. 274–275. [Google Scholar] [CrossRef]
- Geilfus, C.-M. The pH of the apoplast: Dynamic factor with functional impact under stress. Mol. Plant 2017, 10, 1371–1386. [Google Scholar] [CrossRef] [PubMed]
- Felle, H.H. The H+/Cl− symporter in root hair cells of Sinapis alba (an electrophysiological study using ion-selective microelectrodes). Plant Physiol. 1994, 106, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Barbez, E.; Dünser, K.; GAidora, A.; Lendl, T.; Busch, W. Auxin sterrs root cell expansion via apoplastic pH regulation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2017, 114, E4884–E4893. [Google Scholar] [CrossRef] [PubMed]
- Wegner, L.H.; Zimmermann, U. Bicarbonate-induced alkalinization of the xylem sap in intact maize seedlings as measured in situ with a novel xylem pH probe. Plant Physiol. 2004, 136, 3469–3477. [Google Scholar] [CrossRef] [PubMed]
- Felle, H.H. pH regulation in anoxic plants. Ann. Bot. 2005, 96, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Tolleter, D.; Chochois, V.; Poiré, R.; Price, G.D.; Badger, M.R. Measuring CO2 and HCO3− permeabilities of isolated chloroplasts using MIMS-18O approach. J. Exp. Bot. 2017, 68, 3915–3924. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Nagano, Y. Plant acetyl-CoA carboxylase: Structure, biosynthesis, regulation, and gene manipulation for plant breeding. Biosci. Biotechnol. Biochem. 2004, 68, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Nikolau, B.J.; Ohlrogge, J.B.; Wurtele, E.S. Plant biotin-containing carboxylases. Arch. Biochem. Biophys. 2003, 414, 211–222. [Google Scholar] [CrossRef]
- Tong, L. Structure and function of biotin-dependent carboxylases. Cell Mol. Life Sci. 2013, 70, 863–891. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Feng, L.; Jeffrey, P.D.; Shi, Y.; Morel, F.M.M. Structure and metal exchange in the cadmium carbonic anhydrase of marine diatoms. Nature 2008, 452, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Alterio, V.; Langella, E.; De Dimone, G.; Monti, S.S. Cadmium-containing carbonic anhydrase CDCA1 in marine diatom Thalassiosira weissflogii. Mar. Drugs 2015, 13, 1688–1697. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, M.; MacKenzie, T.D.B.; Morse, D. An external carbonic anhydrase in a free-living marine dinoflagellate may circumvent diffusion-limited carbon acquisition. Plant Physiol. 2008, 147, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, Q.; Liu, F.C.; Xie, W.F.; Wang, G.D.; Gao, Q.H.; Duan, K. Family-wide expression characterization of Arabidopsis beta-carbonic anhydrase genes using qRT-PCR and Promoter::GUS fusions. Biochimie 2014, 97, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Parisi, G.; Perales, M.; Fornasari, M.S.; Colaneri, A.; González-Schain, N.; Gómez-Casati, D.; Zimmermann, S.; Brennike, A.; Araya, A.; Ferry, J.; et al. Gamma carbonic anhydrases in plant mitochondria. Plant Mol. Biol. 2004, 55, 193–207. [Google Scholar] [CrossRef] [PubMed]
- De Simeone, G.; Du Fiore, A.; Capasso, C.; Supuran, C.T. The zinc coordination pattern in η-carbonic anhydrase from Plasmodium falciparum is different from all other carbonic anhydrase genetic families. Bioorg. Med. Chem Lett. 2015, 25, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; Fisher, G.M.; Andrews, K.T.; Poulsen, S.A.; Capasso, C.; Supuran, C.T. Discovery of a new family of carbonic anhydrases in the malaria pathogen Plasmodium falciparum—The η-carbonic anhydrases. Bioorg. Med. Chem. Lett. 2014, 24, 4389–4396. [Google Scholar] [CrossRef] [PubMed]
- DiMario, R.J.; Machingura, M.C.; Waldrop, G.L.; Moroney, J.V. The many types of carbonic anhydrases in photosynthetic organisms. Plant Sci. 2018, 268, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Kikutani, S.; Nakajima, K.; Nagasato, C.; Tsuji, Y.; Miyatake, A.; Matsuda, Y. Thylakoid luminal θ–carbonic anhydrase critical for growth and photosynthesis in the marine diatom Phaedactylum tricornutum. Proc. Natl. Acad. Sci. USA 2016, 113, 9828–9833. [Google Scholar] [CrossRef] [PubMed]
- Badger, M.R.; Price, G.D. The Role of Carbonic Anhydrase in Photosynthesis. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 1994, 45, 369–392. [Google Scholar] [CrossRef]
- Protoschill-Krebs, G.; Wilhelm, C.; Kesselmeier, J. Consumption of carbonyl sulphide (COS) by higher plant carbonic anhydrase (CA). Atmos. Environ. 1996, 30, 3151–3156. [Google Scholar] [CrossRef]
- Banerjee, S.; Deshpande, P.A. On origin and evolution of carbonic anhydrase isozymes: A phylogenetic analysis from whole-enzyme to active site. Comput. Biol. Chem. 2016, 61, 121–129. [Google Scholar] [CrossRef] [PubMed]
- DiMario, R.J.; Clayton, H.; Mukherjee, A.; Ludwig, M.; Moroney, J.V. Plant carbonic anhydrases: Structures, locations, evolution, and physiological roles. Mol. Plant 2017, 10, 30–46. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J.; Clarke, A.K.; Chen, Z.-Y.; Hugghins, Y.; Moroney, J.V.; Samuelsson, G. A novel α-type carbonic anhydrase associated with the thylakoid membrane in Chlamydomonas reinhardtii is required for growth at ambient CO2. EMBO J. 1998, 17, 1208–1216. [Google Scholar] [CrossRef] [PubMed]
- Fabre, N.; Reiter, I.M.; Becuwe-Linka, N.; Genty, B.; Rumeau, D. Characterization and expression analysis of genes encoding carbonic anhydrases in Arabidopsis. Plant Cell Environ. 2007, 30, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.N.; Ignatova, L.K.; Fedorchuk, T.P.; Ivanov, B.N. Carbonic anhydrases in photosynthetic cells of higher plants. Biochemistry 2015, 80, 674–687. [Google Scholar] [CrossRef] [PubMed]
- Sherstneva, O.N.; Vodeneev, V.A.; Katicheva, L.A.; Surova, L.M.; Sukhov, V.S. Participation if intracellular and extracellular pH changes in photosynthetic response development induced by variation potential in pumpkin seedlings. Biochemistry 2015, 80, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Kader, M.A.; Lindberg, S. Cytosolic calcium and pH signaling in plants under stress. Plant Sign. Behav. 2010, 5, 233–238. [Google Scholar] [CrossRef]
- Hewett-Emmert, D.; Tashian, R.E. Functional diversity, conservation, and convergence in the evolution of the α-, β-, and γ-carbonic anhydrase gene families. Mol. Phylogenet. Evol. 1996, 5, 50–77. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.N.; Fedorchik, T.P.; Vetoshinka, D.V.; Zhurikova, E.M.; Ignatova, L.K.; Ivanov, B.N. Influence of knockout of At4g20990 gene encoding α-CA4 on photosystem II light-harvesting antenna in plants grown under different light intensities and day lengths. Protoplasma. 2018, 255, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Zhurikova, E.M.; Ignatova, L.K.; Rudenko, N.N.; Mudrik, V.A.; Vetoshinka, D.V.; Ivanov, B.N. Participation of two carbonic anhydrases of the lpha family in photosynthetic reactions in Arabidopsis thaliana. Biochemistry 2016, 81, 1182–1187. [Google Scholar] [PubMed]
- Ludwig, M. Carbonic anhydrase and the molecular evolution of C4 photosynthesis. Plant Cell Environ. 2012, 35, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Flemetakis, F.J.; Dimou, M.; Cotzur, D.; Aivalakis, G.; Efrose, R.C.; Kenoutis, C.; Udvardi, M.; Katinakis, P. A Lotus japonicus β-type carbonic anhydrase gene expression pattern suggests distinct physiological roles during nodule development. Biochim. Biophys. Acta Gene Struct. Expr. 2003, 1628, 186–194. [Google Scholar] [CrossRef]
- Floryszak-Wieczorek, J.; Arasimowicz-Jelonek, M. The multifunctional face of plant carbonic anhydrase. Plant Physiol. Biochem. 2017, 112, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Slaymaker, D.H.; Navarre, D.A.; Clark, A.; del Pozo, O.; Martin, G.B.; Klessig, D.F. The tobacco salicylic acid-binding protein 3 (SABP3) is the chloroplast carbonic anhydrase, which exhibits antioxidant activity and plays a role in the hypersensitive response. Proc. Natl. Acad. Sci. USA 2002, 99, 11640–11645. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.J.; Guo, C.; Coleman, J.R. Reduction of plastid localized carbonic anhydrase activity results in reduced Arabidopsis seedling survivorship. Plant Physiol. 2018, 147, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Fromm, S.; Senkler, J.; Eubel, H.; Peterhänsel, C.; Braun, H.P. Life without complex I: Proteome analysis of an Arabidopsis mutant lacking the mitochondrial NADH dehydrogenase complex. J. Exp. Bot. 2016, 67, 3079–3093. [Google Scholar] [CrossRef] [PubMed]
- Arroyave, C.; Tolrà, R.; Chaves, L.; de Souza, M.; Barceló, J.; Poschenrieder, C. A proteomic approach to the mechanisms underlying activation of aluminium resistance in roots of Urochloa decumbens. J. Inorg Biochem. 2018, 181, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Lepiniec, L.; Vidal, J.; Chollet, R.; Gadal, P.; Cretin, C. Phosphoenolpyruvate carboxylase: Structure, regulation and volution. Plant Sci. 1994, 99, 11–124. [Google Scholar] [CrossRef]
- Rao, S.K.; Noël, C.; Magnin, C.; Reiskind, J.B.; Bowes, G. Photosynthetic and other phosphoenolpyruvate carboxylase isoforms in the single-cell, facultative C4 system of Hydrilla verticillata. Plant Physiol. 2002, 130, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Silvera, K.; Winter, K.; Rodriguez, B.L.; Albion, R.L.; Cushman, J.C. Multiple isoforms of phosphoenolpyruvate carboxylase in the Orchidaceae (subtribe Oncidiinae): Implications for the evolution of crassulacean acid metabolism. J. Exp. Bot. 2014, 65, 3623–3636. [Google Scholar] [CrossRef] [PubMed]
- Nimmo, H.G. Control of the phosphorylation of phosphoenolpyruvate carboxylase in higher plants. Arch. Biochem. Biophys. 2003, 414, 189–196. [Google Scholar] [CrossRef]
- Ruiz-Ballesta, I.; feria, A.B.; She, Y.M.; Plaxton, W.C.; Echevarría, C. In vivo monoubiquitination of anaplerotic phosphoenolpyruvate carboxylase at Lys624 in germinating sorghum seeds. J. Exp. Bot. 2014, 65, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, R.; Cejudo, F.J. Identification and expression analysis of a gene encoding a bacterial-type phosphoenolpyruvate carboxylase from Arabidopsis in rice. Plant Physiol. 2003, 132, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Gennidakis, S.; Rao, S.; Greenham, K.; Uhrig, R.G.; O’Leary, B.; Snedden, W.A.; Lu, C.; Plaxton, W.C. Bacterial- and plant-type phosphoenolpyruvate carboxylase polypeptides interact in the heterooligomeric Class-2 PEPC complex of developing castor oil seeds. Plant J. 2007, 52, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; Hill, A.T.; Pyc, M.; Anderson, E.M.; Snedden, W.A.; Mullen, R.T.; She, Y.-M.; Plaxton, W.C. Regulatory phosphorylation of bacterial-type PEP carboxylase by the Ca2+- dependent protein kinase RcCDPK1 in developing castor oil seeds. Plant Physiol. 2017, 174, 1012–1027. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Khuu, N.; Howard, A.S.M.; Mullen, R.T.; Plaxton, W.C. Bacterial and plant-type phosphoenolpyruvate carboxylase isozymes from developing castor oil seeds interact in vivo and associate with the surface of mitochondria. Plant J. 2012, 71, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Plaxton, W.C.; O’Leary, B. The central role of phosphoenolpyruvate metabolism in developing castor oil seeds. In Seed Development. Omics. Technologies Toward Improvement of Seed Quality and Crop Yield; Agrawal, G.K., Rakwal, R., Eds.; Springer: Berlin, Germany, 2012; pp. 279–301. [Google Scholar] [CrossRef]
- Fischinger, S.A.; Schulze, J. The importance of nodule CO2 fixation for the efficiency of symbiotic nitrogen fixation in pea at vegetative growth and during pot formation. J. Exp. Bot. 2010, 61, 2281–2291. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; Kaiser, E.; Yin, X.; Harbinson, J.; Molenaar, J.; Driever, S.; Struik, P.C. Dynamic modelling of limitations on improving leaf CO2 assimilation under fluctuating irradiance. Plant Cell Environ. 2018, 41, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Keeley, J. Aquatic CAM photosynthesis: A brief history of its discovery. Aquat. Bot. 2014, 118, 38–44. [Google Scholar] [CrossRef]
- Niyogi, K.K.; Wolosiuk, R.A.; Malkin, R. Photosynthesis. In Biochemistry & Molecular Biology of Plants, 2nd ed.; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; Wiley & Sons: Oxford, UK, 2015; pp. 508–566. ISBN 9780470714218. [Google Scholar]
- DiMario, R.J.; Quebedeaux, J.C.; Longstreth, D.J.; Dassanayake, M.; Hartman, M.M.; Moroney, J.V. The cytoplasmic carbonic anhydrases βCA2 and βCA4 are required for optimal plant growth at low CO2. Plant Physiol. 2016, 171, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.F.; Sage, T.L.; Kocacinar, F. Photorespiration and the evolution of C4 photosynthesis. Annu. Rev. Plant Biol. 2012, 63, 19–47. [Google Scholar] [CrossRef] [PubMed]
- Ueno, U. Environmental regulation of C3 and C4 differentiation in the amphibious sedge Eleocharis vivipara. Plant Phsyiol. 2001, 127, 1524–1532. [Google Scholar] [CrossRef]
- Lüttge, U. Ecophysiology of Crassulacean acid metabolism (CAM). Ann. Bot. 2004, 93, 629–652. [Google Scholar] [CrossRef] [PubMed]
- Mackinder, L.C.M. The Chlamydomonas CO2-concentrating mechanism and its potential for engineering photosynthesis in plants. New Phytol. 2018, 217, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Hu, H.; Ries, A.; Merilo, E.; Kollist, H.; Schroeder, J.I. Central functions of bicarbonate in S-type anion channel activation and OST1 protein kinase in CO2 signal transduction guard cell. EMBO J. 2011, 30, 1645–1658. [Google Scholar] [CrossRef] [PubMed]
- Engineer, C.B.; Hashimoto-Sugimoto, M.; Negi, J.; Israelsson-Nordsröm, M.; Azoulay-Shemer, T.; Rappel, W.-J.; Iba, K.; Schroeder, J.I. CO2 sensing and CO2 regulation of stomatal conductance: Advances and open questions. Trends Plant Sci. 2016, 21, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Hou, C.; Ren, Z.; Pan, J.J.; Zhang, H.; Bai, F.; Zhang, P.; Zhu, H.; He, Y.; Luo, S.; et al. A molecular pathway for CO2 response in Arabidopsis guard cells. Nat. Commun. 2015, 6, 6057. [Google Scholar] [CrossRef] [PubMed]
- Kinzel, H. Influence of limestone, silicates and soil pH on vegetation. In Encyclopedia of Plant Physiology, New Series; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Heidelberg, Germany, 1983; Volume 12, pp. 201–244. ISBN 3-540-10907-2. [Google Scholar]
- Kruckeberg, A.R. Geology and Plant Life. In The Effects of Landforms and Rock Types on Plants; University of Washington Press: Seattle, WA, USA, 2002; Chapter 5; pp. 103–228. ISBN 0-295-98203-9. [Google Scholar]
- Lee, J.A.; Woolhouse, H.W. A comparative study of bicarbonate inhibition of root growth of certain grasses. New Phytol. 1969, 68, 1–11. [Google Scholar] [CrossRef]
- Terés, J.; Busoms, S.; Pérez, L.; Villaroya, A.; Álvarez-Fernández, A.; Tolrà, R.; Salt, D.; Poschenrieder, C. Soil carbonate drives local adaptation in Arabidopsis thaliana. New Phytol. 2018. under review. [Google Scholar]
- Yang, X.; Römheld, V.; Marschner, H. Effect of bicarbonate on root growth and accumulation of organic acids in Zn-inefficient and Zn-efficient rice cultivars (Oryza sativa L.). Plant Soil 1994, 164, 1–7. [Google Scholar] [CrossRef]
- Raven, L.A.; Smith, F.A. Nitrogen assimilation and transport in vascular land plants in relation to intracellular pH regulation. New Phytol. 1976, 76, 415–431. [Google Scholar] [CrossRef]
- Marschner, H.; Römheld, V. In vivo measurements of root induced pH changes at the soil-root interface: Effect of plant species and nitrogen source. Z. Pflanzenphysiol. 1983, 111, 241–251. [Google Scholar] [CrossRef]
- Veen, B.W. Influence of oxygen deficiency on growth and function of plant roots. Plant Soil 1988, 111, 259–266. [Google Scholar] [CrossRef]
- Lucena, J.J. Effects of bicarbonate, nitrate and other environmental factors on iron deficiency chlorosis: A review. J. Plant Nutr. 2000, 23, 1561–1606. [Google Scholar] [CrossRef]
- Wanek, W.; Popp, M. Effects of rhizospheric bicarbonate on net nitrate uptake and partitioning between the main nitrate utilising processes in Populus canescens and Sambucus nigra. Plant Soil 2000, 221, 13–24. [Google Scholar] [CrossRef]
- Alhendawi, R.A.; Römheld, V.; Kirkby, E.A.; Marschner, H. Influence of increasing bicarbonate concentrations on plant growth, organic acid accumulation in roots and iron uptake by barley, sorghum, and maize. J. Plant Nutr. 2008, 20, 1731–1753. [Google Scholar] [CrossRef]
- Martínez-Cuenca, M.R.; Legaz, F.; Forner Giner, M.A.; Iglesias, D.J. Bicarbonate blocks iron translocation from cotyledons inducing iron stress responses in Citrus roots. J. Plant Physiol. 2013, 170, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Lucena, C.; Rolera, F.J.; Rojas, C.L.; García, M.J.; Alcántara, E.; Pérez-Vicente, R. Bicarbonate blocks the expression of several genes involved in the physiological responses to Fe deficiency of strategy 1 plants. Funct. Plant Biol. 2007, 34, 1002–1009. [Google Scholar] [CrossRef]
- Sisó-Terraza, P.; Luis-Villaroya, A.; Fourcroy, P.; Briat, J.-F.; Abadía, A.; Gaymard, F.; Abadía, J.; Álvarez-Fernández, A. Accumulation and secretion of coumarinolignans and other coumarins in Arabidopsis thaliana roots in response to iron deficiency at high pH. Front. Plant Sci. 2016, 23, 1711. [Google Scholar] [CrossRef] [PubMed]
- Donnini, S.; De Nisi, P.; Gabotti, D.; Tato, L.; Zocchi, G. Adaptive strategies of Parietaria diffusa (M.& K.) to calcareous habitat with limited iron availability. Plant Cell Environ. 2012, 35, 1171–1184. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cinorg Uptake Mechanism | AZ | TRIS | Seagrass Species | References |
|---|---|---|---|---|
| Apoplastic dehydration of HCO3− catalysed by CA | + | − | Zostera marina | [93] |
| Cymodocea nodosa | [96] | |||
| Halophyla ovalis | [98] | |||
| Cymodocea serrulata | [98] | |||
| Cymodocea rotundata | [98] | |||
| Synringodium isoetifolium | [98] | |||
| Halodule wrightii | [98] | |||
| Thalassia hemprichii | [98] | |||
| Thalassodendron ciliatum | [98] | |||
| Enhalus acoroides | [98] | |||
| Posidonia australis | [102] | |||
| Apoplastic dehydration of HCO3− in acid regions | + | + | Halophila stipulacea | [88] |
| Rupia maritima | [88] | |||
| Cymodocea nodosa (?) | [96] | |||
| Cymodocea rotundata | [98] | |||
| Synringodium isoetifolium | [98] | |||
| Halodule wrightii | [98] | |||
| Thalassia hemprichii | [98] | |||
| Thalassodendron ciliatum | [98] | |||
| Enhalus acoroides | [98] | |||
| Plasma membrane HCO3−/H+ symport | − | + | Posidonia oceanica | [27] 1 |
| Zostera marina | [94] | |||
| Halophyla stipulacea | [88] | |||
| Rupia maritima | [88] | |||
| Cymodocea nodosa (?) | [96] | |||
| Halophila ovalis | [98] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poschenrieder, C.; Fernández, J.A.; Rubio, L.; Pérez, L.; Terés, J.; Barceló, J. Transport and Use of Bicarbonate in Plants: Current Knowledge and Challenges Ahead. Int. J. Mol. Sci. 2018, 19, 1352. https://doi.org/10.3390/ijms19051352
Poschenrieder C, Fernández JA, Rubio L, Pérez L, Terés J, Barceló J. Transport and Use of Bicarbonate in Plants: Current Knowledge and Challenges Ahead. International Journal of Molecular Sciences. 2018; 19(5):1352. https://doi.org/10.3390/ijms19051352
Chicago/Turabian StylePoschenrieder, Charlotte, José Antonio Fernández, Lourdes Rubio, Laura Pérez, Joana Terés, and Juan Barceló. 2018. "Transport and Use of Bicarbonate in Plants: Current Knowledge and Challenges Ahead" International Journal of Molecular Sciences 19, no. 5: 1352. https://doi.org/10.3390/ijms19051352
APA StylePoschenrieder, C., Fernández, J. A., Rubio, L., Pérez, L., Terés, J., & Barceló, J. (2018). Transport and Use of Bicarbonate in Plants: Current Knowledge and Challenges Ahead. International Journal of Molecular Sciences, 19(5), 1352. https://doi.org/10.3390/ijms19051352