Na+-Dependent High-Affinity Nitrate, Phosphate and Amino Acids Transport in Leaf Cells of the Seagrass Posidonia oceanica (L.) Delile




Abstract
:1. Introduction
2. Results
2.1. Effect of Na+ on NO3− and Pi Net Uptake in P. oceanica Leaves
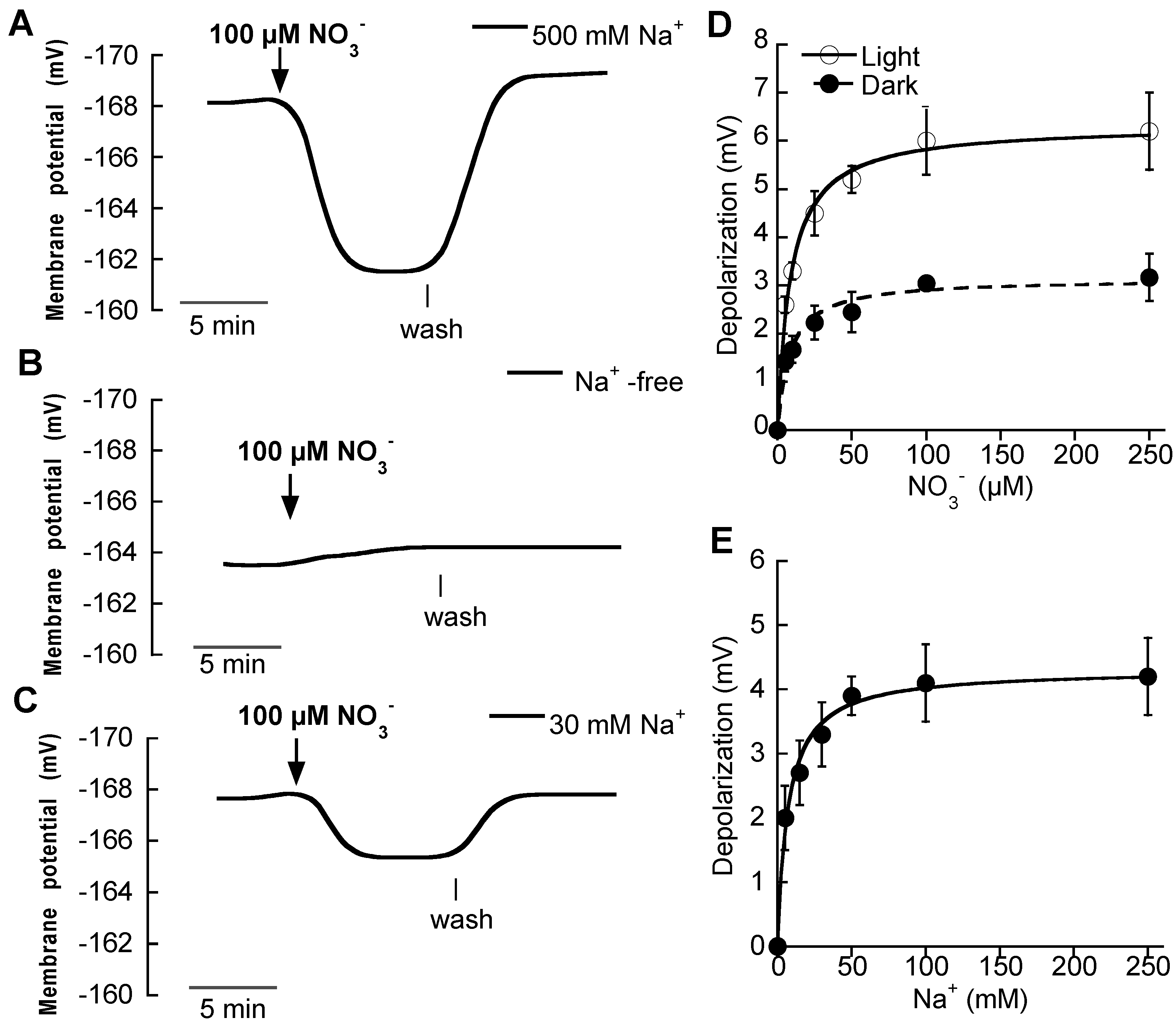
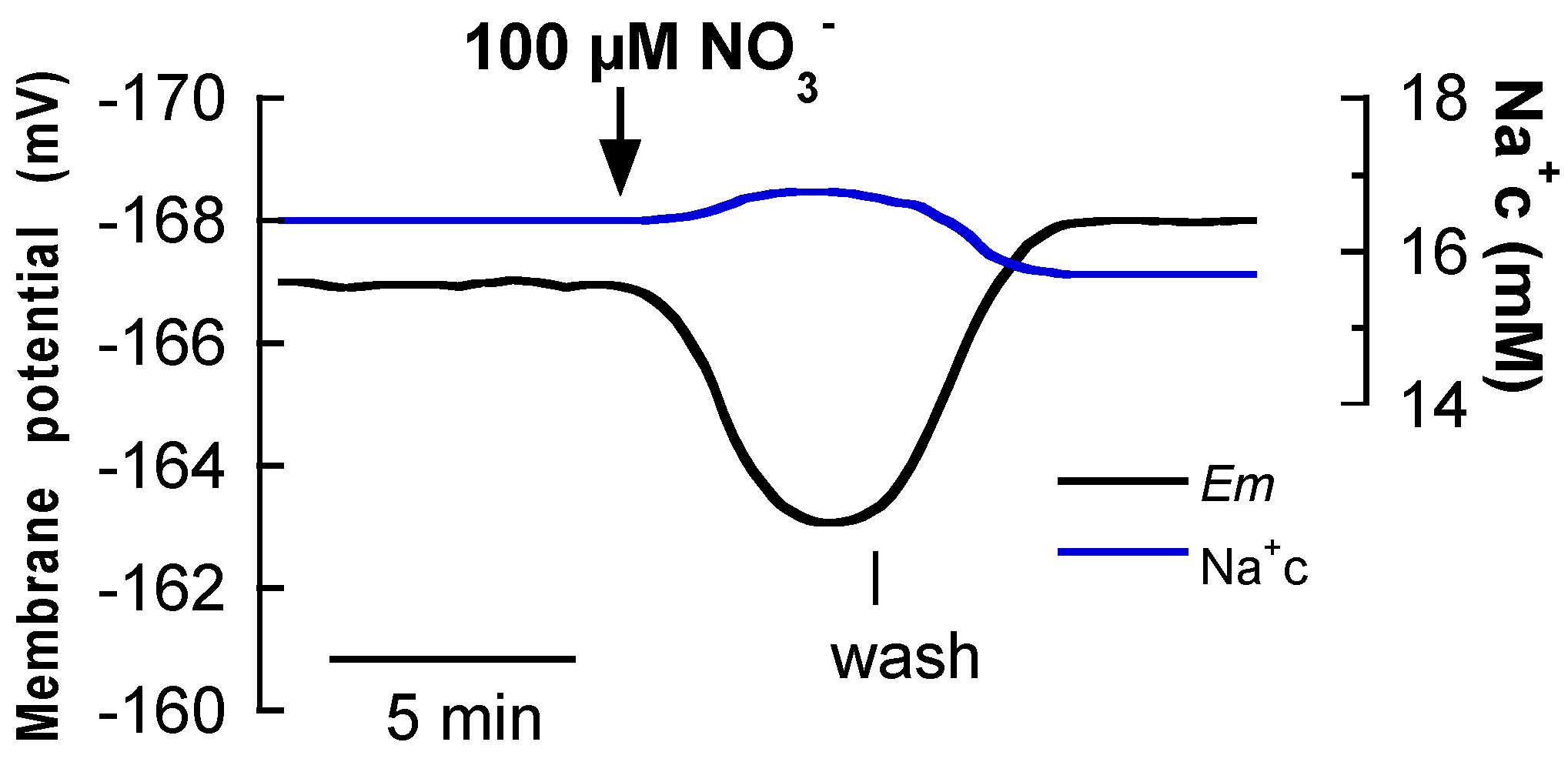
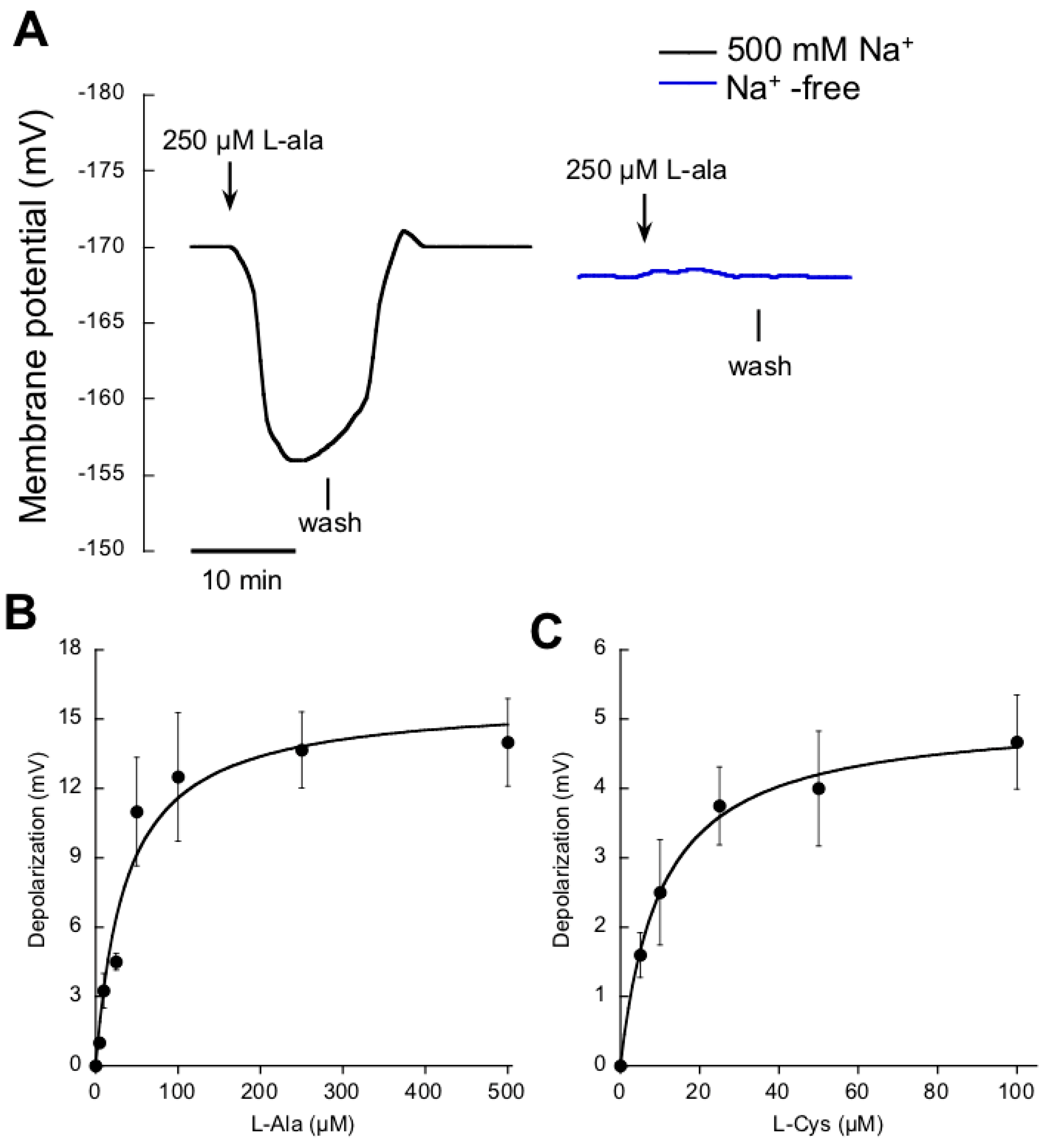
2.2. Na+-Dependent High-Affinity NO3− and Amino Acid Uptake in Single Mesophyll Leaf Cells
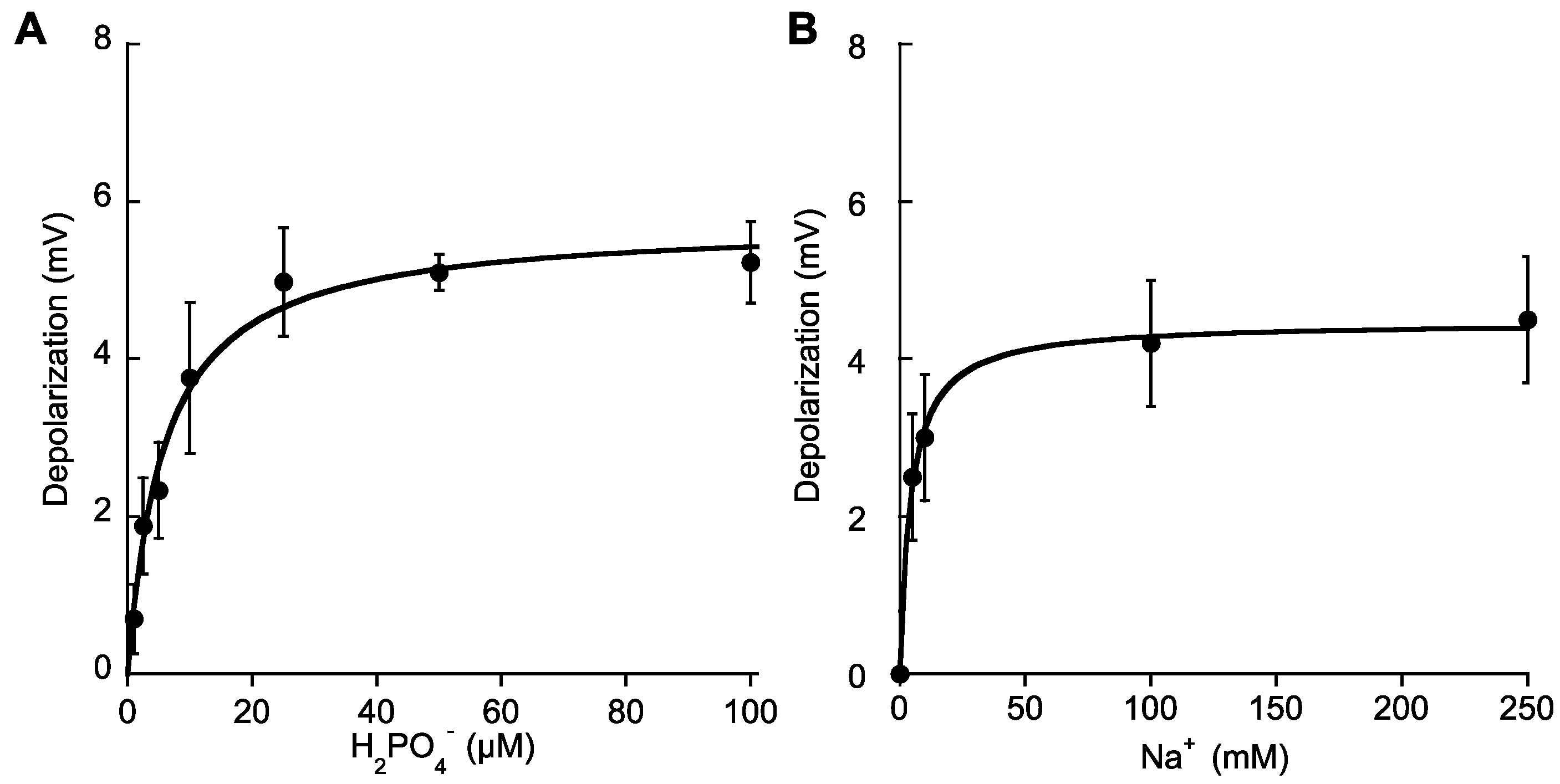
2.3. Na+-Dependent High-Affinity Pi Uptake in Single Mesophyll Leaf Cells
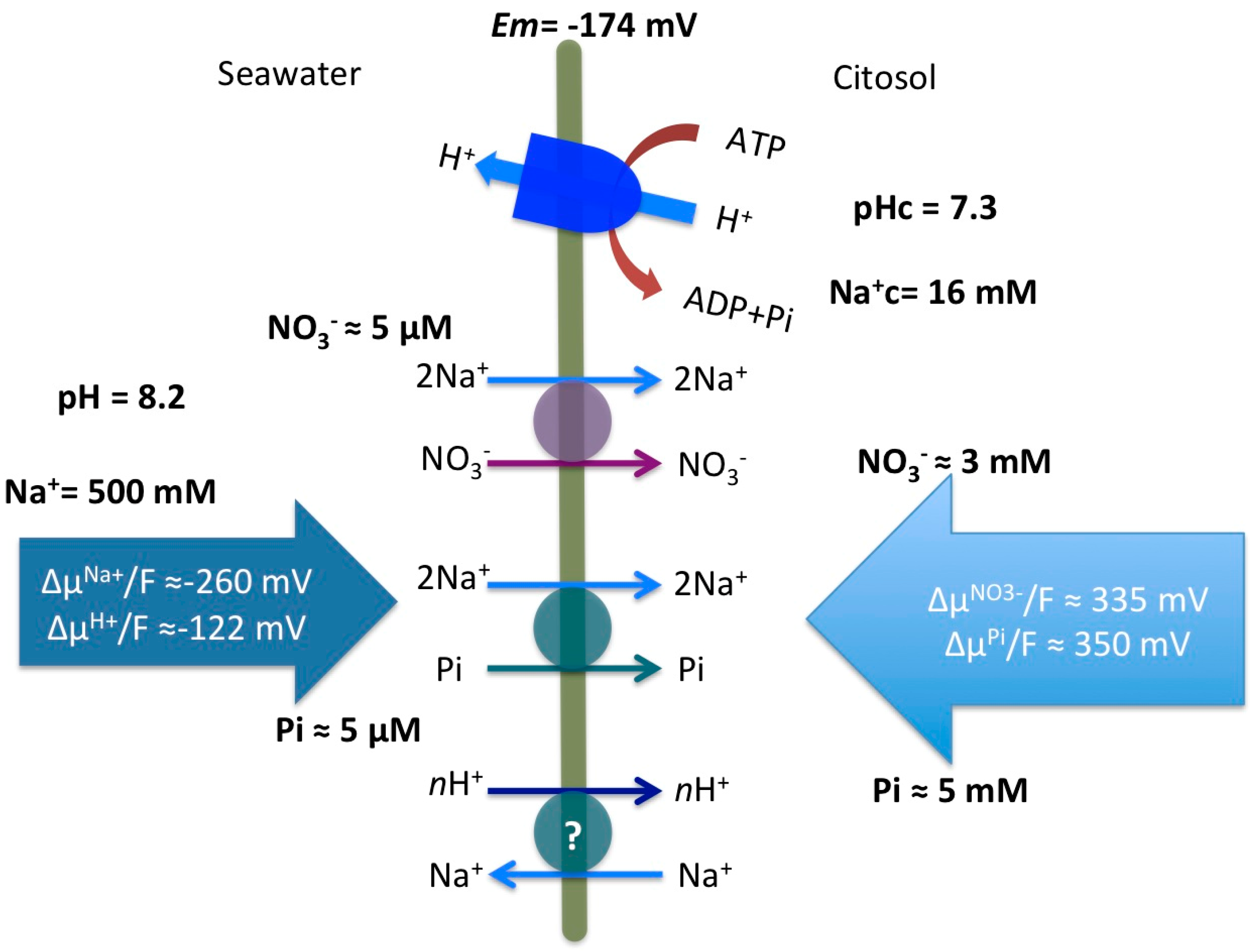
3. Discussion
4. Materials and Methods
4.1. Plant Material and Assay Solutions
4.2. Depletion Experiments
4.3. Electrophysiology
4.4. Data Presentation and Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interests
Abbreviations
| ASW | Artificial Sea Water |
| Dmax | Maximum Depolarization |
| Em | Membrane Potential |
References
- Bethoux, J.P.; El Boukhary, M.S.; Ruiz-Pino, D.; Porin, P.; Copin-Montégut, C. Nutrient, oxygen and carbon ratios, CO2 sequestration and anthropogenic forcing in the Mediterranean Sea. In The Mediterranean Sea. The Handbook of Environmental Chemistry; Saliot, A., Ed.; Spirnger: Belin/Heidelberg, Germany, 2005; pp. 67–87, ISBN-13: 978-3-540-25018-0. [Google Scholar]
- Alcoverro, T.; Romero, J.; Duarte, C.M.; López, N.I. Special and temporal variations in limitation of seagrass Posidonia oceanica growth in the NW Mediterranean. Mar. Ecol. Prog. Ser. 1997, 146, 155–161. [Google Scholar] [CrossRef]
- Pérez, M.; Romero, J.; Duarte, C.M.; Sand-Jensen, K. Phosphorus limitation of Cymodocea nodosa growth. Mar. Biol. 1991, 109, 129–133. [Google Scholar] [CrossRef]
- García-Sánchez, M.J.; Jaime, M.P.; Ramos, A.; Sanders, D.; Fernández, J.A. Sodium-dependent nitrate transport at the plasma membrane of leaf cells of the marine higher plant Zostera marina L. Plant Physiol. 2000, 122, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; Linares-Rueda, A.; García-Sánchez, M.J.; Fernández, J.A. Physiological evidence for a sodium-dependent high-affinity phosphate and nitrate transport at the plasma membrane of leaf and root cells of Zostera marina L. J. Exp. Bot. 2005, 56, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.N.S.; Axelsen, K.B.; Harper, J.F.; Palmgren, M.G. Evolution of plant P-Type ATPases. Front. Plant Sci. 2012, 3, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Fan, X.; Orsel, M.; Smith, S.J.; Wells, D.M. Nitrate transport and signalling. J. Exp. Bot. 2007, 58, 2297–2306. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M.; Glass, D.M. Molecular and physiological aspects of nitrate uptake in plants. Trends Plant. Sci. 1998, 3, 389–395. [Google Scholar] [CrossRef]
- Miller, A.J.; Smith, S.J. Nitrate transport and compartmentation in cereal root cells. J. Exp. Bot. 1996, 47, 843–854. [Google Scholar] [CrossRef]
- Lupini, A.; Mercati, F.; Araniti, F.; Miller, A.J.; Sunseri, F.; Abenavoli, M.R. NAR2.1/NRT2.1 functional interaction with NO3‒ and H+ fluxes in high-affinity nitrate transport in maize root regions. Plant Physiol. Biochem. 2016, 102, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Amtmann, A.; Hammond, J.P.; Armengaud, P.; White, P. Nutrient sensing and signalling in plants: Potassium and phosphorus. Adv. Bot. Res. 2006, 43, 209–257. [Google Scholar] [CrossRef]
- Péret, B.; Clément, M.; Nussaume, L.; Desnos, T. Root developmental adaptation to phosphate starvation: Better safe than sorry. Trends Plant. Sci. 2011, 16, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Péret, B.; Desnos, T.; Jost, R.; Kanno, S.; Berkowitz, O.; Nussaume, L. Root Architecture Responses: In Search of Phosphate. Plant Phys. 2014, 166, 1713–1723. [Google Scholar] [CrossRef] [PubMed]
- Ullrich-Eberius, C.I.; Novacky, A.; Fischer, E.; Lüttge, U. Relationship between energy-dependent phosphate uptake and the electrical membrane potential in Lemna gibba G1. Plant Physiol. 1981, 67, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Rausch, C.; Bucher, M. Molecular mechanisms of phosphate transport in plants. Planta 2002, 21, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Burkholder, J.M.; Tomasko, D.A.; Touchette, B.W. Seagrasses and eutrophication. J. Exp. Mar. Biol. Ecol. 2007, 350, 46–72. [Google Scholar] [CrossRef]
- Lara, C.; Rodríguez, R.; Guerrero, M.G. Sodium-dependent nitrate transport and energetics in cyanobacteria. J. Phycol. 1993, 29, 389–395. [Google Scholar] [CrossRef]
- Shibata, M.; Katoh, H.; Sonoda, M.; Ohkawa, H.; Shimoyama, M.; Fukuzawa, H.; Kaplan, A.; Ogawa, T. Genes essential to sodium-dependent bicarbonate transport in cyanobacteria: Function and phylogenetic analysis. J. Biol. Chem. 2002, 277, 18658–18664. [Google Scholar] [CrossRef] [PubMed]
- Price, G.D.; Woodger, F.J.; Badger, M.R.; Howitt, S.M.; Tucker, L. Identification of a SulP-type bicarbonate transporter in marine cyanobacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 18228–18233. [Google Scholar] [CrossRef] [PubMed]
- Hellebust, J.A. Uptake of organic substrates by Cyclotella cryptica (Bacillariophyceae): Effects of ions, ionophores and metabolic and transport inhibitors. J. Phycol. 1978, 14, 79–83. [Google Scholar] [CrossRef]
- Rees, T.A.; Cresswell, R.C.; Syrett, P.J. Sodium-dependent uptake of nitrate and urea by a marine diatom. Biochim. Biophys. Acta 1980, 596, 141–144. [Google Scholar] [CrossRef]
- Raven, J.A. Energetics and Transport in Aquatic Plants; MBL Lectures in Biology; Alan, R. Liss: New York, NY, USA, 1984; Volume 4, ISBN 0-8451-2203-7. [Google Scholar]
- Fernández, J.A.; García-Sánchez, M.J.; Felle, H. Physiological evidence for a proton pump at the plasma membrane of the marine angiosperm Zostera marina L. J. Exp. Bot. 1999, 50, 1763–1768. [Google Scholar]
- Rubio, L.; Belver, A.; Venema, K.; García-Sánchez, M.J.; Fernández, J.A. Evidence for a sodium efflux mechanism in the leaf cells of the seagrass Zostera marina L. J. Exp. Mar. Biol. Ecol. 2011, 402, 56–64. [Google Scholar] [CrossRef]
- Rubio, L.; García, D.; García-Sánchez, M.J.; Niell, F.X.; Felle, H.H.; Fernández, J.A. Direct uptake of HCO3− in the marine angiosperm Posidonia oceanica (L.) Delile driven by a plasma membrane H+ economy. Plant Cell Environ. 2017, 40, 2820–2830. [Google Scholar] [CrossRef] [PubMed]
- Romero, J.; Lee, K.-S.; Pérez, M.A.; Alcoverro, T. Nutrients dynamics. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 227–254. ISBN 1-4020-2942-X. [Google Scholar]
- Sanders, D.; Hansen, U.-P.; Gradmann, D.; Slayman, C.L. Generalized kinetic analysis of ion-dependent cotransport systems: A unified interpretation of selective ionic effects on Michaelis parameters. J. Membr. Biol. 1984, 77, 123–152. [Google Scholar] [CrossRef] [PubMed]
- Touchette, B.W.; Burkholder, J.M. Review of nitrogen and phosphorus metabolism in seagrasses. J. Exp. Mar. Biol. Ecol. 2000, 250, 133–167. [Google Scholar] [CrossRef]
- Glass, A.D.; Shaff, J.E.; Kochian, L.V. Studies of the uptake of nitrate in barley, IV. Electrophysiology. Plant Physiol. 1992, 99, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Kumar, A.; Li, W.; Wang, Y.; Siddiqi, M.Y.; Crawford, N.M.; Glass, A.D.M. High-affinity nitrate transport in roots of Arabidopsis depends on expression of the NAR2-like gene AtNRT3.1. Plant Phys. 2006, 140, 1036–1046. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant. Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Van Engeland, T.; Bouma, T.J.; Morris, E.P.; Brun, F.G.; Peralta, G.; Lara, M.; Hendriks, I.E.; Soetaert, K.; Middelburg, J.J. Potential uptake of dissolved organic matter by seagrasses and macroalgae. Mar. Ecol. Prog. Ser. 2011, 427, 71–81. [Google Scholar] [CrossRef]
- Nafie, Y.A.; Van Engeland, T.; Van Katwikj, M.M.; Bourma, T. Uptake of nitrogen from compound pools by the seagrass Zostera noltii. J. Exp. Mar. Biol. Ecol. 2014, 460, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Vonk, J.A.; Middelburg, J.J.; Stapel, J.; Bouma, T.J. Dissolved organic nitrogen uptake by seagrasses. Limnol. Oceanogr. 2008, 53, 542–548. [Google Scholar] [CrossRef]
- Lee, Y.H.; Foster, J.; Chen, J.; Voll, L.; Weber, A.; Tegeder, M. AAP1 transports uncharged amino acids into roots of Arabidopsis. Plant J. 2007, 50, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Svennerstam, H.; Ganeteg, U.; Näsholm, T. Root uptake of cationic amino acids by Arabidopsis depends on functional expression of amino acid permease 5. New Phytol. 2008, 180, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Svennerstam, H.; Jämtgård, S.; Ahmad, I.; Huss-Danell, K.; Näsholm, T.; Ganeteg, U. Transporters in Arabidopsis roots mediating uptake of amino acids at naturally occurring concentrations. New Phytol. 2011, 191, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Hirner, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.; Koch, W. Arabidopsis LHT1 is a high-affinity transporter for cellular amino acid uptake in both root epidermis and leaf mesophyll. Plant Cell 2006, 18, 1931–1946. [Google Scholar] [CrossRef] [PubMed]
- Svennerstam, H.; Ganeteg, U.; Bellini, C.; Näsholm, T. Comprehensive screening of Arabidopsis mutants suggests the lysine histidine transporter 1 to be involved in plant uptake of amino acids. Plant Physiol. 2007, 143, 1853–1860. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Gumy, C.; Blatter, E.; Boeffel, S.; Fricke, W.; Rentsch, D. In planta function of compatible solute transporters of the AtProT family. J. Exp. Bot. 2011, 62, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M. Transporters involved in source to sink partitioning of amino acids and ureides: Opportunities for crop improvement. J. Exp. Bot. 2014, 65, 1865–1878. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Lloréns, J.L.; Niell, F.X. Short-term phosphate uptake kinetics in Zostera noltii Hornem: A comparison between excised leaves and sediment-rooted plants. Hydrobiologia 1995, 297, 17–27. [Google Scholar] [CrossRef]
- Stapel, J.; Aarts, T.L.; van Duynhoven, B.H.M.; de Groot, J.D.; van den Hoogen, P.H.W.; Hemminga, M.A. Nutrient uptake by leaves and roots of the seagrass Thalassia hemprichii in the Spermonde Archipelago, Indonesia. Mar. Ecol. Prog. Ser. 1996, 134, 195–206. [Google Scholar] [CrossRef]
- Gras, A.F.; Marguerite, S.K.; Madden, C.J. Phosphorus uptake kinetics of a dominant tropical seagrass Thalassia testudinum. Aquat. Bot. 2003, 76, 299–315. [Google Scholar] [CrossRef]
- Rae, A.L.; Cybinski, D.H.; Jarmey, J.M.; Smith, F.W. Characterization of two phosphate transporters from barley: Evidence for diverse function and kinetic properties among members of the Pht1 family. Plant Mol. Biol. 2003, 53, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, M.G. Plant plasma membrane H+-ATPases: Powerhouses for nutrient uptake. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 817–845. [Google Scholar] [CrossRef] [PubMed]
- Falhof, J.; Pedersen, J.T.; Fuglsang, A.T.; Palmgren, M. Plasma Membrane H+-ATPase Regulation in the Center of Plant Physiology. Mol. Plant 2016, 9, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Munns, R.; Colmer, T.D. Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot. 2015, 115, 419–431. [Google Scholar] [CrossRef] [PubMed]
- García-Robledo, E.; Corzo, E.; Papaspyrou, S. A fast and direct spectrophotometric method for the sequential determination of nitrate and nitrite at low concentrations in small volumes. Mar. Chem. 2014, 162, 30–36. [Google Scholar] [CrossRef]
- Fernández, J.A.; Niell, F.X.; Lucena, J. A rapid and sensitive automated determination of phosphate in natural waters. Limnol. Oceanogr. 1985, 30, 227–230. [Google Scholar] [CrossRef]
- Felle, H. A study of the current-voltage relationships of electrogenic active and passive membrane elements in Riccia fluitans. Biochim. Biophys. Acta 1981, 646, 151–160. [Google Scholar] [CrossRef]
- Rubio, L.; Linares-Rueda, A.; García-Sánchez, M.J.; Fernández, J.A. Ammonium uptake kineticts in root and leaf cells of Zostera marina L. J. Exp. Mar. Biol. Ecol. 2007, 352, 271–279. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Net Uptake Rate | ASW | Na+-Free ASW |
|---|---|---|
| μmol NO3−·gFW−1·h−1 | 0.34 ± 0.01 | 0.09 ± 0.01 * |
| μmol Pi·gFW−1·h−1 | 0.04 ± 0.003 | 0.01 ± 0.002 * |
| Nutrient | Km (μM) | Dmax (mV) |
|---|---|---|
| NO3− (Light) | 8.7 ± 1 (7.2 ± 1.1 mM Na+) 1 | 6.3 ± 0.2 |
| NO3− (Dark) | 8.2 ± 1.7 | 3.1 ± 0.1 |
| l-ala | 37 ± 11 | 16 ± 1.2 |
| l-cys | 10 ± 1 | 5 ± 0.2 |
| Pi | 5.8 ± 1 (4.3 ± 0.5 mM Na+) 1 | 5.7 ± 0.2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio, L.; García-Pérez, D.; García-Sánchez, M.J.; Fernández, J.A. Na+-Dependent High-Affinity Nitrate, Phosphate and Amino Acids Transport in Leaf Cells of the Seagrass Posidonia oceanica (L.) Delile. Int. J. Mol. Sci. 2018, 19, 1570. https://doi.org/10.3390/ijms19061570
Rubio L, García-Pérez D, García-Sánchez MJ, Fernández JA. Na+-Dependent High-Affinity Nitrate, Phosphate and Amino Acids Transport in Leaf Cells of the Seagrass Posidonia oceanica (L.) Delile. International Journal of Molecular Sciences. 2018; 19(6):1570. https://doi.org/10.3390/ijms19061570
Chicago/Turabian StyleRubio, Lourdes, Delia García-Pérez, María Jesús García-Sánchez, and José A. Fernández. 2018. "Na+-Dependent High-Affinity Nitrate, Phosphate and Amino Acids Transport in Leaf Cells of the Seagrass Posidonia oceanica (L.) Delile" International Journal of Molecular Sciences 19, no. 6: 1570. https://doi.org/10.3390/ijms19061570
APA StyleRubio, L., García-Pérez, D., García-Sánchez, M. J., & Fernández, J. A. (2018). Na+-Dependent High-Affinity Nitrate, Phosphate and Amino Acids Transport in Leaf Cells of the Seagrass Posidonia oceanica (L.) Delile. International Journal of Molecular Sciences, 19(6), 1570. https://doi.org/10.3390/ijms19061570