The Spleen as an Optimal Site for Islet Transplantation and a Source of Mesenchymal Stem Cells


Abstract
:1. Introduction
2. Islet Transplantation and Its Obstacles
3. Candidate Transplantation Sites for Islets
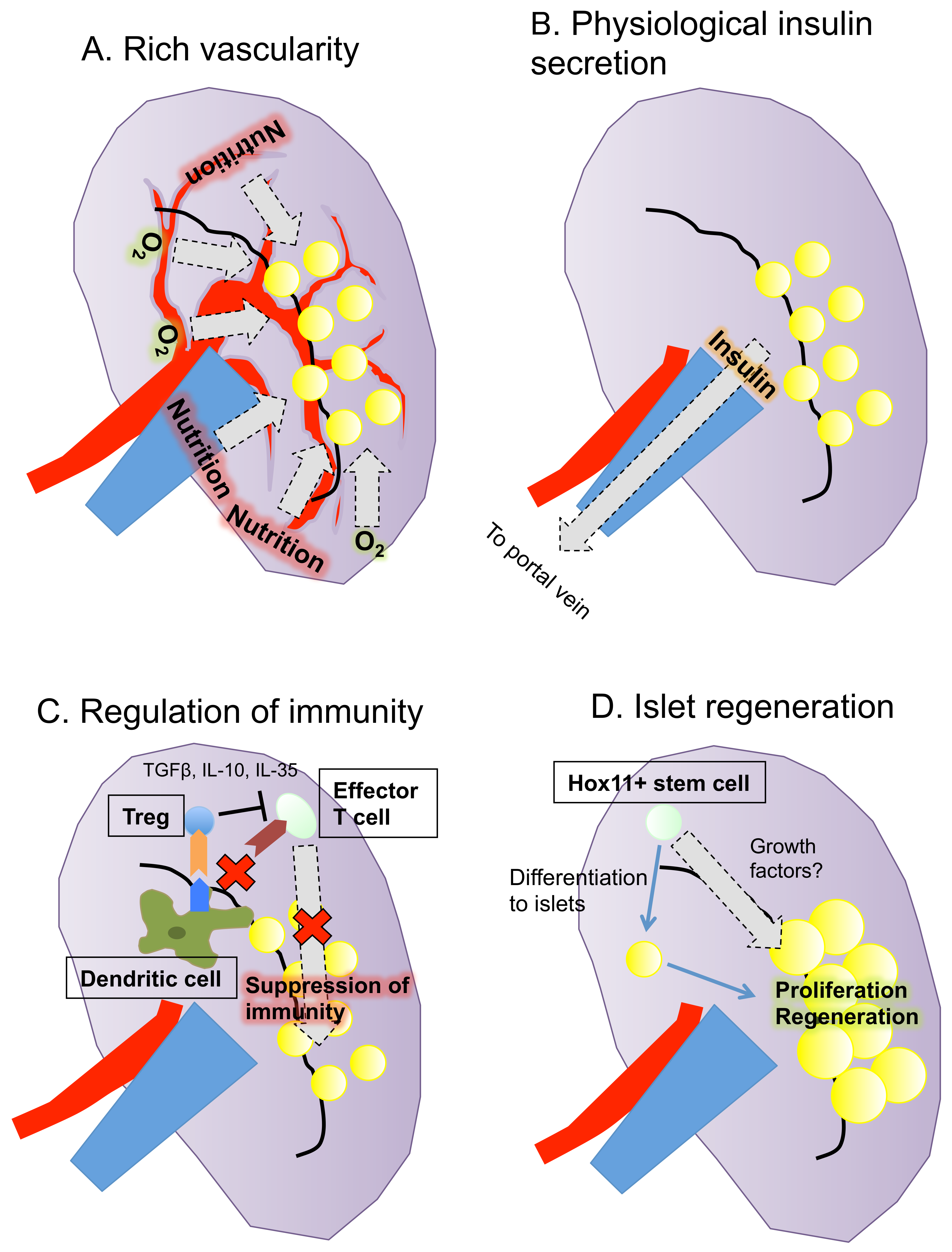
4. Characteristics of the Spleen as an Islet Transplant Site
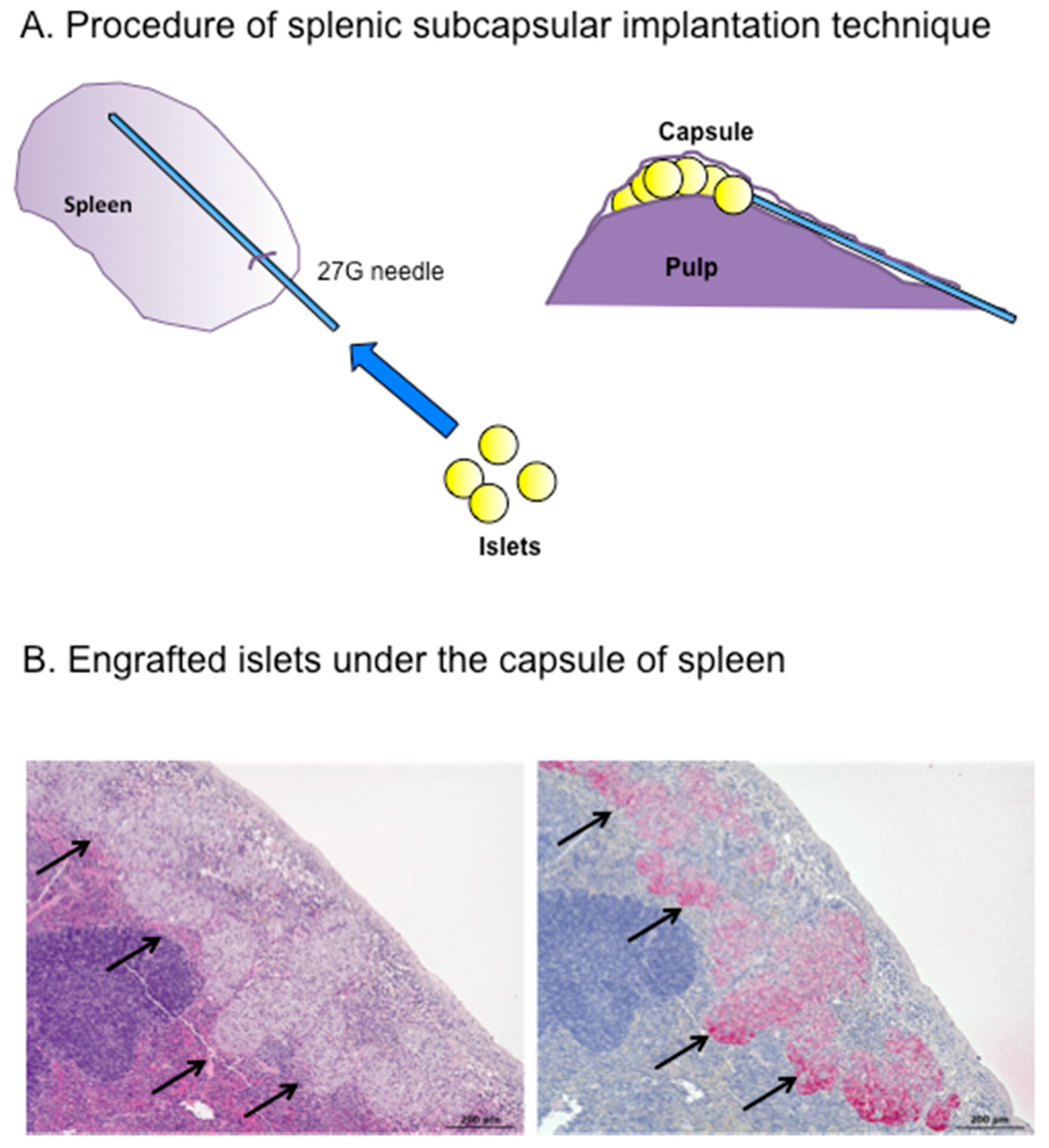
5. Outcomes of Intrasplenic Islet Transplantation
6. Future Clinical Intrasplenic Islet Transplantation
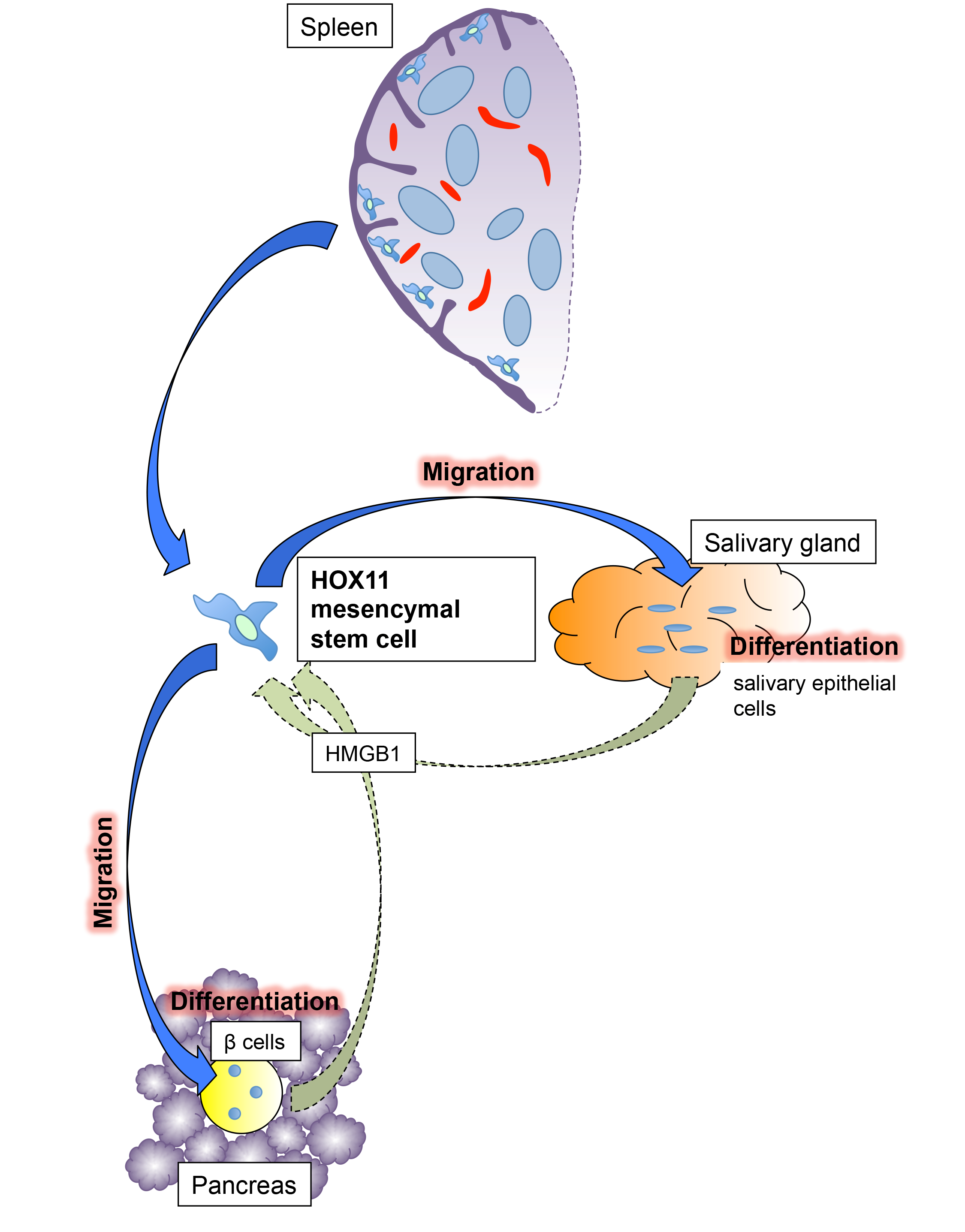
7. Spleen as a Source of Mesenchymal Stem Cells
8. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| HMGB1 | high-mobility group box 1 |
| IBMIR | instant blood-mediated inflammatory reaction |
| IL | interleukin |
| NOD | non-obese |
| Rrm2b | ribonucleoside-diphosphate reductase subunit M2 b |
| T-ALL | T-cell acute lymphoblastic leukemia |
| Tregs | regulatory T cells |
References
- Golub, R.; Tan, J.; Watanabe, T.; Brendolan, A. Origin and Immunological Functions of Spleen Stromal Cells. Trends Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Newland, A.; Provan, D.; Myint, S. Preventing severe infection after splenectomy. BMJ 2005, 331, 417–418. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, G.; Scalera, A.; Finelli, C. Liver-spleen axis: Intersection between immunity, infections and metabolism. World J. Gastroenterol. 2013, 19, 3534–3542. [Google Scholar] [CrossRef] [PubMed]
- Fiorina, P.; Shapiro, A.M.; Ricordi, C.; Secchi, A. The clinical impact of islet transplantation. Am. J. Transplant. 2008, 8, 1990–1997. [Google Scholar] [CrossRef] [PubMed]
- Fiorina, P.; Folli, F.; Bertuzzi, F.; Maffi, P.; Finzi, G.; Venturini, M.; Socci, C.; Davalli, A.; Orsenigo, E.; Monti, L.; et al. Long-term beneficial effect of islet transplantation on diabetic macro-/microangiopathy in type 1 diabetic kidney-transplanted patients. Diabetes Care 2003, 26, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Fiorina, P.; Folli, F.; Zerbini, G.; Maffi, P.; Gremizzi, C.; Di Carlo, V.; Socci, C.; Bertuzzi, F.; Kashgarian, M.; Secchi, A. Islet transplantation is associated with improvement of renal function among uremic patients with type I diabetes mellitus and kidney transplants. J. Am. Soc. Nephrol. 2003, 14, 2150–2158. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.E.; Radosevich, D.M.; Bellin, M.D.; Hering, B.J.; Beilman, G.J.; Dunn, T.B.; Chinnakotla, S.; Vickers, S.M.; Bland, B.; Balamurugan, A.N.; et al. Total pancreatectomy and islet autotransplantation for chronic pancreatitis. J. Am. Coll. Surg. 2012, 214, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Ricordi, C.; Strom, T.B. Clinical islet transplantation: Advances and immunological challenges. Nat. Rev. Immunol. 2004, 4, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Ricordi, C.; Lacy, P.E.; Finke, E.H.; Olack, B.J.; Scharp, D.W. Automated method for isolation of human pancreatic islets. Diabetes 1988, 37, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Scharp, D.W.; Lacy, P.E.; Santiago, J.V.; McCullough, C.S.; Weide, L.G.; Falqui, L.; Marchetti, P.; Gingerich, R.L.; Jaffe, A.S.; Cryer, P.E.; et al. Insulin independence after islet transplantation into type I diabetic patient. Diabetes 1990, 39, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Tzakis, A.G.; Ricordi, C.; Alejandro, R.; Zeng, Y.; Fung, J.J.; Todo, S.; Demetris, A.J.; Mintz, D.H.; Starzl, T.E. Pancreatic islet transplantation after upper abdominal exenteration and liver replacement. Lancet 1990, 336, 402–405. [Google Scholar] [CrossRef] [Green Version]
- Ekmekcioglu, O.; Turkan, S.; Yildiz, S.; Gunes, Z.E. Comparison of tacrolimus with a cyclosporine microemulsion for immunosuppressive therapy in kidney transplantation. Turk. J. Urol. 2013, 39, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, T.K.; Isbel, N.M.; Barraclough, K.A.; Campbell, S.B.; McWhinney, B.C.; Staatz, C.E. Comparison of the influence of cyclosporine and tacrolimus on the pharmacokinetics of prednisolone in adult male kidney transplant recipients. Clin. Drug Investig. 2014, 34, 183–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, A.M.; Lakey, J.R.; Ryan, E.A.; Korbutt, G.S.; Toth, E.; Warnock, G.L.; Kneteman, N.M.; Rajotte, R.V. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N. Engl. J. Med. 2000, 343, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Kahan, B.D. Sirolimus: A comprehensive review. Expert Opin. Pharmacother. 2001, 2, 1903–1917. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Mukherjee, U. A comprehensive review of immunosuppression used for liver transplantation. J. Transplant. 2009, 2009, 701464. [Google Scholar] [CrossRef] [PubMed]
- Milo, R.; Osherov, M. Daclizumab and its use in multiple sclerosis treatment. Drugs Today (Barc) 2017, 53, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Bellin, M.D.; Kandaswamy, R.; Parkey, J.; Zhang, H.J.; Liu, B.; Ihm, S.H.; Ansite, J.D.; Witson, J.; Bansal-Pakala, P.; Balamurugan, A.N.; et al. Prolonged insulin independence after islet allotransplants in recipients with type 1 diabetes. Am. J. Transplant. 2008, 8, 2463–2470. [Google Scholar] [CrossRef] [PubMed]
- Semenkow, S.; Li, S.; Kahlert, U.D.; Raabe, E.H.; Xu, J.; Arnold, A.; Janowski, M.; Oh, B.C.; Brandacher, G.; Bulte, J.W.M.; et al. An immunocompetent mouse model of human glioblastoma. Oncotarget 2017, 8, 61072–61082. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.A.; Paty, B.W.; Senior, P.A.; Bigam, D.; Alfadhli, E.; Kneteman, N.M.; Lakey, J.R.; Shapiro, A.M. Five-year follow-up after clinical islet transplantation. Diabetes 2005, 54, 2060–2069. [Google Scholar] [CrossRef] [PubMed]
- Barton, F.B.; Rickels, M.R.; Alejandro, R.; Hering, B.J.; Wease, S.; Naziruddin, B.; Oberholzer, J.; Odorico, J.S.; Garfinkel, M.R.; Levy, M.; et al. Improvement in outcomes of clinical islet transplantation: 1999–2010. Diabetes Care 2012, 35, 1436–1445. [Google Scholar] [CrossRef] [PubMed]
- Balamurugan, A.N.; Naziruddin, B.; Lockridge, A.; Tiwari, M.; Loganathan, G.; Takita, M.; Matsumoto, S.; Papas, K.; Trieger, M.; Rainis, H.; et al. Islet product characteristics and factors related to successful human islet transplantation from the Collaborative Islet Transplant Registry (CITR) 1999–2010. Am. J. Transplant. 2014, 14, 2595–2606. [Google Scholar] [CrossRef] [PubMed]
- Mellgren, A.; Schnell Landstrom, A.H.; Petersson, B.; Andersson, A. The renal subcapsular site offers better growth conditions for transplanted mouse pancreatic islet cells than the liver or spleen. Diabetologia 1986, 29, 670–672. [Google Scholar] [CrossRef] [PubMed]
- Hiller, W.F.; Klempnauer, J.; Luck, R.; Steiniger, B. Progressive deterioration of endocrine function after intraportal but not kidney subcapsular rat islet transplantation. Diabetes 1991, 40, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.B.; Knight, M.J.; Scharp, D.W.; Ballinger, W.F.; Lacy, P.E. Effect of transplantation site on the results of pancreatic islet isografts in diabetic rats. Diabetologia 1973, 9, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Yasunami, Y.; Lacy, P.E.; Finke, E.H. A new site for islet transplantation—A peritoneal-omental pouch. Transplantation 1983, 36, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Kin, T.; Korbutt, G.S.; Rajotte, R.V. Survival and metabolic function of syngeneic rat islet grafts transplanted in the omental pouch. Am. J. Transplant. 2003, 3, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Osama Gaber, A.; Chamsuddin, A.; Fraga, D.; Fisher, J.; Lo, A. Insulin independence achieved using the transmesenteric approach to the portal vein for islet transplantation. Transplantation 2004, 77, 309–311. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; McGrath, K.M.; Bottino, R.; Dons, E.M.; Long, C.; Kumar, G.; Ekser, B.; Echeverri, G.J.; Hata, J.; Haruma, K.; et al. Technique of endoscopic biopsy of islet allografts transplanted into the gastric submucosal space in pigs. Cell Transplant. 2013, 22, 2335–2344. [Google Scholar] [CrossRef] [PubMed]
- Sakata, N.; Aoki, T.; Yoshimatsu, G.; Tsuchiya, H.; Hata, T.; Katayose, Y.; Egawa, S.; Unno, M. Strategy for clinical setting in intramuscular and subcutaneous islet transplantation. Diabetes Metab. Res. Rev. 2014, 30, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mojibian, M.; Harder, B.; Hurlburt, A.; Bruin, J.E.; Asadi, A.; Kieffer, T.J. Implanted islets in the anterior chamber of the eye are prone to autoimmune attack in a mouse model of diabetes. Diabetologia 2013, 56, 2213–2221. [Google Scholar] [CrossRef] [PubMed]
- Xin, Z.L.; Ge, S.L.; Wu, X.K.; Jia, Y.J.; Hu, H.T. Intracerebral xenotransplantation of semipermeable membrane- encapsuled pancreatic islets. World J. Gastroenterol. 2005, 11, 5714–5717. [Google Scholar] [CrossRef] [PubMed]
- Bobzien, B.; Yasunami, Y.; Majercik, M.; Lacy, P.E.; Davie, J.M. Intratesticular transplants of islet xenografts (rat to mouse). Diabetes 1983, 32, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.; Scothorne, R.J. Extended survival of pancreatic islet allografts in the testis of guinea-pigs. J. Anat. 1977, 124 Pt 1, 1–8. [Google Scholar] [PubMed]
- Cantarelli, E.; Citro, A.; Pellegrini, S.; Mercalli, A.; Melzi, R.; Dugnani, E.; Jofra, T.; Fousteri, G.; Mondino, A.; Piemonti, L. Transplant Site Influences the Immune Response After Islet Transplantation: Bone Marrow Versus Liver. Transplantation 2017, 101, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.M.; Ketchum, R.J.; Tomaszewski, J.E.; Naji, A.; Barker, C.F.; Brayman, K.L. Intrathymic islet transplantation in the canine: I. Histological and functional evidence of autologous intrathymic islet engraftment and survival in pancreatectomized recipients. Transplantation 2002, 73, 842–852. [Google Scholar] [CrossRef] [PubMed]
- White, S.A.; London, N.J.; Johnson, P.R.; Davies, J.E.; Pollard, C.; Contractor, H.H.; Hughes, D.P.; Robertson, G.S.; Musto, P.P.; Dennison, A.R. The risks of total pancreatectomy and splenic islet autotransplantation. Cell Transplant. 2000, 9, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Owen, R.J.; Ryan, E.A.; O’Kelly, K.; Lakey, J.R.; McCarthy, M.C.; Paty, B.W.; Bigam, D.L.; Kneteman, N.M.; Korbutt, G.S.; Rajotte, R.V.; et al. Percutaneous transhepatic pancreatic islet cell transplantation in type 1 diabetes mellitus: Radiologic aspects. Radiology 2003, 229, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Schramm, R.; Scheuer, C.; Yamauchi, J.; Vollmar, B.; Menger, M.D. Heat shock preconditioning impairs revascularization of freely transplanted pancreatic islets. Transplantation 2002, 74, 1544–1550. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, P.O.; Palm, F.; Andersson, A.; Liss, P. Markedly decreased oxygen tension in transplanted rat pancreatic islets irrespective of the implantation site. Diabetes 2001, 50, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Sakata, N.; Hayes, P.; Tan, A.; Chan, N.K.; Mace, J.; Peverini, R.; Sowers, L.; Pearce, W.J.; Chinnock, R.; Obenaus, A.; et al. MRI assessment of ischemic liver after intraportal islet transplantation. Transplantation 2009, 87, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Sakata, N.; Obenaus, A.; Chan, N.; Mace, J.; Chinnock, R.; Hathout, E. Factors affecting islet graft embolization in the liver of diabetic mice. Islets 2009, 1, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Bottino, R.; Fernandez, L.A.; Ricordi, C.; Lehmann, R.; Tsan, M.F.; Oliver, R.; Inverardi, L. Transplantation of allogeneic islets of Langerhans in the rat liver: Effects of macrophage depletion on graft survival and microenvironment activation. Diabetes 1998, 47, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Lu, X.; Yi, S.; Wu, J.; O’Hara, J.M.; Hawthorne, W.J.; Hucker, K.; O’Connell, P.J. Selective rejection of porcine islet xenografts by macrophages. Xenotransplantation 2008, 15, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Ishiyama, K.; Rawson, J.; Omori, K.; Mullen, Y. Liver natural killer cells play a role in the destruction of islets after intraportal transplantation. Transplantation 2011, 91, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Bennet, W.; Sundberg, B.; Groth, C.G.; Brendel, M.D.; Brandhorst, D.; Brandhorst, H.; Bretzel, R.G.; Elgue, G.; Larsson, R.; Nilsson, B.; et al. Incompatibility between human blood and isolated islets of Langerhans: A finding with implications for clinical intraportal islet transplantation? Diabetes 1999, 48, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Johansson, H.; Lukinius, A.; Moberg, L.; Lundgren, T.; Berne, C.; Foss, A.; Felldin, M.; Kallen, R.; Salmela, K.; Tibell, A.; et al. Tissue factor produced by the endocrine cells of the islets of Langerhans is associated with a negative outcome of clinical islet transplantation. Diabetes 2005, 54, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Van der Windt, D.J.; Bottino, R.; Casu, A.; Campanile, N.; Cooper, D.K. Rapid loss of intraportally transplanted islets: An overview of pathophysiology and preventive strategies. Xenotransplantation 2007, 14, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.M.; Goss, J.A.; Deng, S.; Wolf, B.A.; Markmann, E.; Palanjian, M.; Shock, A.P.; Feliciano, S.; Brunicardi, F.C.; Barker, C.F.; et al. Elevated portal vein drug levels of sirolimus and tacrolimus in islet transplant recipients: Local immunosuppression or islet toxicity? Transplantation 2003, 76, 1623–1625. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.A.; Lakey, J.R.; Paty, B.W.; Imes, S.; Korbutt, G.S.; Kneteman, N.M.; Bigam, D.; Rajotte, R.V.; Shapiro, A.M. Successful islet transplantation: Continued insulin reserve provides long-term glycemic control. Diabetes 2002, 51, 2148–2157. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Kin, T.; Shapiro, A.M. A comparison of islet autotransplantation with allotransplantation and factors elevating acute portal pressure in clinical islet transplantation. J. Hepatobiliary Pancreat. Sci. 2012, 19, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.M.; Ahmad, S.A. Management of acute post-operative portal venous thrombosis. J. Gastrointest. Surg. 2010, 14, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.W.; Sutton, R.; McShane, P.; Peters, M.; Morris, P.J. Exocrine contamination impairs implantation of pancreatic islets transplanted beneath the kidney capsule. J. Surg. Res. 1988, 45, 432–442. [Google Scholar] [CrossRef]
- Jindal, R.M.; Sidner, R.A.; McDaniel, H.B.; Johnson, M.S.; Fineberg, S.E. Intraportal vs kidney subcapsular site for human pancreatic islet transplantation. Transplant. Proc. 1998, 30, 398–399. [Google Scholar] [CrossRef]
- Van der Windt, D.J.; Echeverri, G.J.; Ijzermans, J.N.; Cooper, D.K. The choice of anatomical site for islet transplantation. Cell Transplant. 2008, 17, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Ao, Z.; Matayoshi, K.; Lakey, J.R.; Rajotte, R.V.; Warnock, G.L. Survival and function of purified islets in the omental pouch site of outbred dogs. Transplantation 1993, 56, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Berman, D.M.; O’Neil, J.J.; Coffey, L.C.; Chaffanjon, P.C.; Kenyon, N.M.; Ruiz, P., Jr.; Pileggi, A.; Ricordi, C.; Kenyon, N.S. Long-term survival of nonhuman primate islets implanted in an omental pouch on a biodegradable scaffold. Am. J. Transplant. 2009, 9, 91–104. [Google Scholar] [CrossRef] [PubMed]
- McQuilling, J.P.; Sittadjody, S.; Pareta, R.; Pendergraft, S.; Clark, C.J.; Farney, A.C.; Opara, E.C. Retrieval of Microencapsulated Islet Grafts for Post-transplant Evaluation. Methods Mol. Biol. 2017, 1479, 157–171. [Google Scholar] [PubMed]
- Pareta, R.; McQuilling, J.P.; Sittadjody, S.; Jenkins, R.; Bowden, S.; Orlando, G.; Farney, A.C.; Brey, E.M.; Opara, E.C. Long-term function of islets encapsulated in a redesigned alginate microcapsule construct in omentum pouches of immune-competent diabetic rats. Pancreas 2014, 43, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Aomatsu, Y.; Iwata, H.; Kin, T.; Kanehiro, H.; Hisanga, M.; Ko, S.; Nagao, M.; Harb, G.; Nakajima, Y. Survival of microencapsulated islets at 400 days posttransplantation in the omental pouch of NOD mice. Cell Transplant. 2006, 15, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.D.; Headen, D.M.; Aquart, J.; Johnson, C.T.; Shea, L.D.; Shirwan, H.; Garcia, A.J. Vasculogenic hydrogel enhances islet survival, engraftment, and function in leading extrahepatic sites. Sci. Adv. 2017, 3, e1700184. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Fujita, M.; Bottino, R.; Piganelli, J.D.; McGrath, K.; Li, J.; Lee, W.; Iwase, H.; Wijkstrom, M.; Bertera, S.; et al. Endoscopic biopsy of islet transplants in the gastric submucosal space provides evidence of islet graft rejection in diabetic pigs. Islets 2016, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Streilein, J.W. Unraveling immune privilege. Science 1995, 270, 1158–1159. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Mital, P.; Dufour, J.M. Testisimmune privilege—Assumptions versus facts. Anim. Reprod. 2013, 10, 3–15. [Google Scholar] [PubMed]
- Perez, V.L.; Saeed, A.M.; Tan, Y.; Urbieta, M.; Cruz-Guilloty, F. The eye: A window to the soul of the immune system. J. Autoimmun. 2013, 45, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Pachter, J.S.; de Vries, H.E.; Fabry, Z. The blood-brain barrier and its role in immune privilege in the central nervous system. J. Neuropathol. Exp. Neurol. 2003, 62, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Larocque, D.; Sanderson, N.S.; Bergeron, J.; Curtin, J.F.; Girton, J.; Wibowo, M.; Bondale, N.; Kroeger, K.M.; Yang, J.; Lacayo, L.M.; et al. Exogenous fms-like tyrosine kinase 3 ligand overrides brain immune privilege and facilitates recognition of a neo-antigen without causing autoimmune neuropathology. Proc. Natl. Acad. Sci. USA 2010, 107, 14443–14448. [Google Scholar] [CrossRef] [PubMed]
- Hedger, M.P. Immunophysiology and pathology of inflammation in the testis and epididymis. J. Androl. 2011, 32, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Farooq, S.M.; Elkhatib, W.F.; Ashour, H.M. The in vivo and in vitro induction of anterior chamber associated immune deviation to myelin antigens in C57BL/6 mice. Brain Behav. Immun. 2014, 42, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Polonsky, K.S.; Given, B.D.; Hirsch, L.; Shapiro, E.T.; Tillil, H.; Beebe, C.; Galloway, J.A.; Frank, B.H.; Karrison, T.; Van Cauter, E. Quantitative study of insulin secretion and clearance in normal and obese subjects. J. Clin. Investig. 1988, 81, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Home, P.D. Plasma insulin profiles after subcutaneous injection: How close can we get to physiology in people with diabetes? Diabetes Obes. Metab. 2015, 17, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.S.; Harper, N.; Bevan, D.J.; Staines, N.A. Suppression of collagen induced arthritis by oral administration of type II collagen: Changes in immune and arthritic responses mediated by active peripheral suppression. Autoimmunity 1993, 16, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; al-Sabbagh, A.; Santos, L.M.; Das, M.P.; Weiner, H.L. Epitopes of myelin basic protein that trigger TGF-beta release after oral tolerization are distinct from encephalitogenic epitopes and mediate epitope-driven bystander suppression. J. Immunol. 1993, 151, 7307–7315. [Google Scholar] [PubMed]
- Nagatani, K.; Dohi, M.; To, Y.; Tanaka, R.; Okunishi, K.; Nakagome, K.; Sagawa, K.; Tanno, Y.; Komagata, Y.; Yamamoto, K. Splenic dendritic cells induced by oral antigen administration are important for the transfer of oral tolerance in an experimental model of asthma. J. Immunol. 2006, 176, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, T.T. Suppressor T cells, rebranded as regulatory T cells, emerge from the wilderness bearing surface markers. Gut 2002, 51, 311–312. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Nishikawa, H. Roles of regulatory T cells in cancer immunity. Int. Immunol. 2016, 28, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.J.; Hawthorne, W.J.; Walters, S.N.; Patel, A.T.; O’Connell, P.J.; Chapman, J.R.; Allen, R.D. Induction of allogeneic islet tolerance in a large-animal model. Cell Transplant. 2000, 9, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Kodama, S.; Kuhtreiber, W.; Fujimura, S.; Dale, E.A.; Faustman, D.L. Islet regeneration during the reversal of autoimmune diabetes in NOD mice. Science 2003, 302, 1223–1227. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Kodama, S.; Ryu, K.; Schoenfeld, D.A.; Faustman, D.L. Reversal of established autoimmune diabetes by restoration of endogenous beta cell function. J. Clin. Investig. 2001, 108, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Iwahashi, S.; Kanak, M.A.; Shimoda, M.; Takita, M.; Chujo, D.; Tamura, Y.; Rahman, A.M.; Chung, W.Y.; Onaca, N.; et al. Elevation of high-mobility group box 1 after clinical autologous islet transplantation and its inverse correlation with outcomes. Cell Transplant. 2014, 23, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, N.; Itoh, T.; Watarai, H.; Sekine-Kondo, E.; Nagata, N.; Okamoto, K.; Mera, T.; Yamamoto, H.; Yamada, S.; Maruyama, I.; et al. High-mobility group box 1 is involved in the initial events of early loss of transplanted islets in mice. J. Clin. Investig. 2010, 120, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Nitta, T.; Nishinakamura, H.; Kojima, D.; Mera, T.; Ono, J.; Kodama, S.; Yasunami, Y. HMGB1-Mediated Early Loss of Transplanted Islets Is Prevented by Anti-IL-6R Antibody in Mice. Pancreas 2015, 44, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Nishinakamura, H.; Kumano, K.; Takahashi, H.; Kodama, S. The Spleen Is an Ideal Site for Inducing Transplanted Islet Graft Expansion in Mice. PLoS ONE 2017, 12, e0170899. [Google Scholar] [CrossRef] [PubMed]
- Faustman, D.L.; Davis, M. Stem cells in the spleen: Therapeutic potential for Sjogren’s syndrome, type I diabetes, and other disorders. Int. J. Biochem. Cell Biol. 2010, 42, 1576–1579. [Google Scholar] [CrossRef] [PubMed]
- Kodama, S.; Davis, M.; Faustman, D.L. Diabetes and stem cell researchers turn to the lowly spleen. Sci. Aging Knowl. Environ. 2005, 2005, pe2. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.W.; Tan, S.H.; Lee, W.K.; Yap, H.K.; Aw, S.E.; Wong, H.B. Glucose tolerance test and insulin levels in children with transfusion-dependent thalassaemia. Ann. Trop. Paediatr. 1985, 5, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Wohlrab, F.; Schmidt, S.; Kloting, I.; Wilke, B.; Cossel, L. Ducto-insular proliferation of beta-cells after syngeneic islet transplantation into the spleen of streptozotocin-diabetic Lewis rats. Int. J. Pancreatol. 1989, 5, 77–83. [Google Scholar] [PubMed]
- Tanaka, H.; Arakawa, H.; Yamaguchi, T.; Shiraishi, K.; Fukuda, S.; Matsui, K.; Takei, Y.; Nakamura, Y. A ribonucleotide reductase gene involved in a p53-dependent cell-cycle checkpoint for DNA damage. Nature 2000, 404, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Kolb, E.; Ruckert, R.; Largiader, F. Intraportal and intrasplenic autotransplantation of pancreatic islets in the dog. Eur. Surg. Res. 1977, 9, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Feldman, S.D.; Hirshberg, G.E.; Dodi, G.; Raizman, M.E.; Scharp, D.W.; Ballinger, W.F.; Lacy, P.E. Intrasplenic islet isografts. Surgery 1977, 82, 386–394. [Google Scholar] [PubMed]
- Schulak, J.A.; Stuart, F.P.; Reckard, C.R. Physiologic aspects of intrasplenic autotransplantation of pancreatic fragments in the dog after 24 hours of cold storage. J. Surg. Res. 1978, 24, 125–131. [Google Scholar] [CrossRef]
- Gray, B.N.; Caruso, G.; Alford, F.; Chisholm, D. Insulin and glucagon responses of transplanted intrasplenic pancreatic islets. Arch. Surg. 1979, 114, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Merrell, R.C.; Marincola, F.; Maeda, M.; Cobb, L.; Basadonna, G. The metabolic response of intrasplenic islet autografts. Surg. Gynecol. Obstet. 1985, 160, 552–556. [Google Scholar] [PubMed]
- Merrell, R.C.; Maeda, M.; Basadonna, G.; Marincola, F.; Cobb, L. Suppression, stress, and accommodation of transplanted islets of Langerhans. Diabetes 1985, 34, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Gores, P.F.; Boudreaux, J.P.; Hesse, U.J.; Najarian, J.S.; Sutherland, D.E. Canine islet autografts with and without administration of cyclosporine. Surgery 1987, 101, 557–561. [Google Scholar] [PubMed]
- Evans, M.G.; Warnock, G.L.; Rajotte, R.V. Comparison of sites for transplantation of canine pancreatic microfragments. Diabetes Res. 1989, 10, 35–41. [Google Scholar] [PubMed]
- Van der Vliet, J.A.; Kaufman, D.B.; Meloche, R.M.; Heise, J.W.; Field, M.J.; Heil, J.E.; Najarian, J.S.; Sutherland, D.E. A simple method of canine pancreatic islet isolation and intrahepatic transplantation. J. Surg. Res. 1989, 46, 129–134. [Google Scholar] [CrossRef]
- Van der Vliet, J.A.; Kaufman, D.B.; Meloche, R.M.; van der Hem, L.G.; Field, M.J.; Sutherland, D.E. Canine pancreatic islet transplantation: A comparison of two isolation techniques. Eur. Surg. Res. 1990, 22, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Warnock, G.L.; Dabbs, K.D.; Evans, M.G.; Cattral, M.S.; Kneteman, N.M.; Rajotte, R.V. Critical mass of islets that function after implantation in a large mammalian. Horm. Metab. Res. Suppl. 1990, 25, 156–161. [Google Scholar] [PubMed]
- Scharp, D.W.; Marchetti, P.; Swanson, C.; Newton, M.; McCullough, C.S.; Olack, B. The effect of transplantation site and islet mass on long-term survival and metabolic and hormonal function of canine purified islet autografts. Cell Transplant. 1992, 1, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Motojima, K.; Kohara, N.; Maeda, J.; Yamaguchi, M.; Kanematsu, T. A comparison of endocrine and exocrine function after pancreatic fragment autotransplantation into splenic pulp, portal vein, and hepatic parenchyma. Transplantation 1992, 53, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, P.; Olack, B.; Swanson, C.; Newton, M.; Scharp, D.W. Assessment of long-term (1 year) graft survival and metabolic and hormonal changes after intrasplenic canine pancreatic microfragment transplantation. Diabetes Metab. 1993, 19, 17–24. [Google Scholar]
- Toledo-Pereyra, L.H.; Gordon, D.A.; MacKenzie, G.H. Application of cryopreservation techniques to islet cell allotransplantation. Cryobiology 1983, 20, 205–210. [Google Scholar] [CrossRef]
- Sandler, S.; Kojima, Y.; Andersson, A. Cryopreservation of mouse pancreatic islets. Effects of fast cooling on islet B cell function and on the outcome of islet transplantation. Transplantation 1986, 42, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Heise, J.W.; Casanova, D.; Field, M.J.; Munn, S.R.; Najarian, J.S.; Sutherland, D.E. Cold storage preservation of pancreatic tissue prior to and after islet preparation in a dog autotransplantation model. J. Surg. Res. 1989, 47, 30–38. [Google Scholar] [CrossRef]
- Casanova, D.; Heise, J.W.; Field, M.J.; Munn, S.R.; Sutherland, D.E. Successful transplantation of canine islets of Langerhans after 24 hours cold-storage. Diabetes Res. 1989, 10, 31–34. [Google Scholar] [PubMed]
- Merrell, R.C.; Mahoney, M.E.; Basadonna, G.; Cobb, L.F.; Maeda, M. Failure of canine islet allografts and autografts with cyclosporine. Surgery 1985, 98, 324–329. [Google Scholar] [PubMed]
- Basadonna, G.; Kakizaki, K.; Merrell, R.C. Effect of cyclosporine on established islet autografts. J. Surg. Res. 1986, 40, 450–454. [Google Scholar] [CrossRef]
- Kneteman, N.M.; Alderson, D.; Scharp, D.W. Long-term normoglycemia in pancreatectomized dogs following pancreatic islet allotransplantation and cyclosporine immunosuppression. Transplantation 1987, 44, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Kneteman, N.M.; Marchetti, P.; Tordjman, K.; Bier, D.M.; Santiago, J.V.; Swanson, C.J.; Olack, B.J.; Scharp, D.W. Effects of cyclosporine on insulin secretion and insulin sensitivity in dogs with intrasplenic islet autotransplants. Surgery 1992, 111, 430–437. [Google Scholar] [PubMed]
- Hesse, U.J.; Meyer, G.P.; Weyer, J.; Danis, J.; Pichlmaier, H. Islet isolation and autotransplantation in pigs. Zentralbl. Chir. 1994, 119, 653–660. [Google Scholar] [PubMed]
- Gray, D.W. Islet isolation and transplantation techniques in the primate. Surg. Gynecol. Obstet. 1990, 170, 225–232. [Google Scholar] [PubMed]
- Gray, D.W.; Warnock, G.L.; Sutton, R.; Peters, M.; McShane, P.; Morris, P.J. Successful autotransplantation of isolated islets of Langerhans in the cynomolgus monkey. Br. J. Surg. 1986, 73, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Sutton, R.; Gray, D.W.; Burnett, M.; McShane, P.; Turner, R.C.; Morris, P.J. Metabolic function of intraportal and intrasplenic islet autografts in cynomolgus monkeys. Diabetes 1989, 38 (Suppl. 1), 182–184. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, D.F.; Reece-Smith, H.; McShane, P.; Denton, T.; Morris, P.J. Effect of cyclosporin A on allotransplanted pancreatic fragments to the spleen of totally pancreatectomized dogs. Transplantation 1982, 33, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Janney, C.G.; Lacy, P.E.; Finke, E.H.; Davie, J.M. Prolongation of intrasplenic islet xenograft survival. Am. J. Pathol. 1982, 107, 1–5. [Google Scholar] [PubMed]
- Yakimets, W.J.; Lakey, J.R.; Yatscoff, R.W.; Katyal, D.; Ao, Z.; Finegood, D.T.; Rajotte, R.V.; Kneteman, N.M. Prolongation of canine pancreatic islet allograft survival with combined rapamycin and cyclosporine therapy at low doses. Rapamycin efficacy is blood level related. Transplantation 1993, 56, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A. Reversal of hyperglycemia by intrasplenic transplantation of 4-week-cultured allogeneic mouse islets. Diabetes 1982, 31 (Suppl. 4), 55–59. [Google Scholar] [CrossRef] [PubMed]
- Mehigan, D.G.; Zuidema, G.D.; Cameron, J.L. Pancreatic islet transplantation in dogs. Critical factors in technique. Am. J. Surg. 1981, 141, 208–212. [Google Scholar] [CrossRef]
- Andersson, A.; Eriksson, U.; Petersson, B.; Reibring, L.; Swenne, I. Failure of successful intrasplenic transplantation of islets from lean mice to cure obese-hyperglycaemic mice, despite islet growth. Diabetologia 1981, 20, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Steffes, M.W.; Nielsen, O.; Dyrberg, T.; Baekkeskov, S.; Scott, J.; Lernmark, A. Islet transplantation in mice differing in the I and S subregions of the H-2 complex. Transplantation 1981, 31, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Warnock, G.L.; Rajotte, R.V.; Procyshyn, A.W. Normoglycemia after reflux of islet-containing pancreatic fragments into the splenic vascular bed in dogs. Diabetes 1983, 32, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A. The influence of hyperglycaemia, hyperinsulinaemia and genetic background on the fate of intrasplenically implanted mouse islets. Diabetologia 1983, 25, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Kneteman, N.M.; Rajotte, R.V.; Procyshyn, A.W. Canine pancreatic fragment allotransplantation with cyclosporine A. J. Surg. Res. 1985, 39, 285–293. [Google Scholar] [CrossRef]
- Hayek, A.; Lopez, A.D.; Beattie, G.M. Decrease in the number of neonatal islets required for successful transplantation by strict metabolic control of diabetic rats. Transplantation 1988, 45, 940–942. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, B.; Kohler, E.; Kloting, I.; Besch, W.; Ziegler, M. Survival of islet isografts despite cytotoxicity against pancreatic islets measured in vitro. Exp. Clin. Endocrinol. 1990, 95, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Korsgren, O.; Jansson, L.; Sandler, S.; Andersson, A. Hyperglycemia-induced B cell toxicity. The fate of pancreatic islets transplanted into diabetic mice is dependent on their genetic background. J. Clin. Investig. 1990, 86, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Eizirik, D.L.; Jansson, L.; Flodstrom, M.; Hellerstrom, C.; Andersson, A. Mechanisms of defective glucose-induced insulin release in human pancreatic islets transplanted to diabetic nude mice. J. Clin. Endocrinol. Metab. 1997, 82, 2660–2663. [Google Scholar] [CrossRef] [PubMed]
- Stokes, R.A.; Simond, D.M.; Burns, H.; Patel, A.T.; O’Connell, P.J.; Gunton, J.E.; Hawthorne, W.J. Transplantation sites for porcine islets. Diabetologia 2017, 60, 1972–1976. [Google Scholar] [CrossRef] [PubMed]
- Stokes, R.A.; Cheng, K.; Lalwani, A.; Swarbrick, M.M.; Thomas, H.E.; Loudovaris, T.; Kay, T.W.; Hawthorne, W.J.; O’Connell, P.J.; Gunton, J.E. Transplantation sites for human and murine islets. Diabetologia 2017, 60, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- Gustavson, S.M.; Rajotte, R.V.; Hunkeler, D.; Lakey, J.R.; Edgerton, D.S.; Neal, D.W.; Snead, W.L.; Penaloza, A.R.; Cherrington, A.D. Islet auto-transplantation into an omental or splenic site results in a normal beta cell but abnormal alpha cell response to mild non-insulin-induced hypoglycemia. Am. J. Transplant. 2005, 5, 2368–2377. [Google Scholar] [CrossRef] [PubMed]
- Cesta, M.F. Normal structure, function, and histology of the spleen. Toxicol. Pathol. 2006, 34, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Gaba, R.C.; Jin, B.; Lewandowski, R.J.; Riaz, A.; Memon, K.; Ryu, R.K.; Sato, K.T.; Kulik, L.M.; Mulcahy, M.F.; et al. Perfusion reduction at transcatheter intraarterial perfusion MR imaging: A promising intraprocedural biomarker to predict transplant-free survival during chemoembolization of hepatocellular carcinoma. Radiology 2014, 272, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.S.; Mitchell, D.L.; Vasquez, K.M. High mobility group protein B1 enhances DNA repair and chromatin modification after DNA damage. Proc. Natl. Acad. Sci. USA 2008, 105, 10320–10325. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E.; Manfredi, A.A. High-mobility group box 1 (HMGB1) protein at the crossroads between innate and adaptive immunity. Immunol. Rev. 2007, 220, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Ban, T.; Wang, Z.; Choi, M.K.; Kawamura, T.; Negishi, H.; Nakasato, M.; Lu, Y.; Hangai, S.; Koshiba, R.; et al. HMGB proteins function as universal sentinels for nucleic-acid-mediated innate immune responses. Nature 2009, 462, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Limana, F.; Germani, A.; Zacheo, A.; Kajstura, J.; Di Carlo, A.; Borsellino, G.; Leoni, O.; Palumbo, R.; Battistini, L.; Rastaldo, R.; et al. Exogenous high-mobility group box 1 protein induces myocardial regeneration after infarction via enhanced cardiac C-kit+ cell proliferation and differentiation. Circ. Res. 2005, 97, e73–e83. [Google Scholar] [CrossRef] [PubMed]
- Chadburn, A. The spleen: Anatomy and anatomical function. Semin. Hematol. 2000, 37 (Suppl. 1), 13–21. [Google Scholar] [CrossRef]
- Tran, S.D.; Kodama, S.; Lodde, B.M.; Szalayova, I.; Key, S.; Khalili, S.; Faustman, D.L.; Mezey, E. Reversal of Sjogren’s-like syndrome in non-obese diabetic mice. Ann. Rheum. Dis. 2007, 66, 812–814. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.A.; Rowan-Hull, A.M.; Johnson, P.R. The spleen--a potential source of new islets for transplantation? J. Pediatr. Surg. 2008, 43, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, M.A.; Leiter, E.H. The NOD mouse model of type 1 diabetes: As good as it gets? Nat. Med. 1999, 5, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, D.; Liu, D.; Fan, Z.; Zhang, H.; Liu, O.; Ding, G.; Gao, R.; Zhang, C.; Ding, Y.; et al. Allogeneic mesenchymal stem cell treatment alleviates experimental and clinical Sjogren syndrome. Blood 2012, 120, 3142–3151. [Google Scholar] [CrossRef] [PubMed]
- Owens, B.M.; Hawley, T.S.; Spain, L.M.; Kerkel, K.A.; Hawley, R.G. TLX1/HOX11-mediated disruption of primary thymocyte differentiation prior to the CD4+CD8+ double-positive stage. Br. J. Haematol. 2006, 132, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, A.A.; Neuberg, D.S.; Dodge, R.K.; Paietta, E.; Larson, R.A.; Wiernik, P.H.; Rowe, J.M.; Caligiuri, M.A.; Bloomfield, C.D.; Look, A.T. Prognostic importance of TLX1 (HOX11) oncogene expression in adults with T-cell acute lymphoblastic leukaemia. Lancet 2004, 363, 535–536. [Google Scholar] [CrossRef]
- Raju, K.; Tang, S.; Dube, I.D.; Kamel-Reid, S.; Bryce, D.M.; Breitman, M.L. Characterization and developmental expression of Tlx-1, the murine homolog of HOX11. Mech. Dev. 1993, 44, 51–64. [Google Scholar] [CrossRef]
- Dear, T.N.; Colledge, W.H.; Carlton, M.B.; Lavenir, I.; Larson, T.; Smith, A.J.; Warren, A.J.; Evans, M.J.; Sofroniew, M.V.; Rabbitts, T.H. The Hox11 gene is essential for cell survival during spleen development. Development 1995, 121, 2909–2915. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transplant Site | Advantages | Disadvantages |
|---|---|---|
| Liver |
|
|
| Kidney |
|
|
| Omental pouch |
|
|
| Mesentery |
|
|
| Gastrointestinal tract |
|
|
| Muscle and subcutaneous tissue |
|
|
| Immune privilege site (brain, testis, eye, thymus) |
|
|
| Authors [Reference] | Publication Year | Transplant Model | Comments |
|---|---|---|---|
| Kolb E et al. [89] | 1977 | Auto (dog) |
|
| Feldman SD et al. [90] | 1977 | Auto (dog) |
|
| Gray BN et al. [92] | 1979 | Auto (dog) |
|
| Mehigan DG et al. [119] | 1981 | Auto (dog) |
|
| Andersson A et al. [120] | 1981 | Iso (mouse) |
|
| Steffes MW et al. [121] | 1981 | Iso, allo (mouse) |
|
| Du Toit DF et al. [115] | 1982 | Allo (dog) |
|
| Janney CG et al. [116] | 1982 | Xeno (rat-to-mouse) |
|
| Andersson A. [118] | 1982 | Allo (mouse) |
|
| Toledo-Pereyra LH et al. [103] | 1983 | Allo (dog) |
|
| Warnock GL et al. [122] | 1983 | Iso (dog) |
|
| Andersson A [123] | 1983 | Iso (mouse) |
|
| Merrell RC et al. [93,94] | 1985 | Auto (dog) |
|
| Kneteman NM et al. [124] | 1985 | Allo (dog) |
|
| Gray DW et al. [113] | 1986 | Auto (monkey) |
|
| Gores PF et al. [95] | 1987 | Auto (dog) |
|
| Kneteman NM et al. [109] | 1987 | Allo (dog) |
|
| Hayek A et al. [125] | 1988 | Iso (rat) |
|
| Sutton R et al. [114] | 1989 | Auto (monkey) |
|
| Evans MG et al. [96] | 1989 | Auto (dog) |
|
| van der Vliet JA et al. [97,98] | 1989 | Auto (dog) |
|
| Warnock GL et al. [99] | 1990 | Auto (dog) |
|
| Ziegler B et al. [126] | 1990 | Iso (rat) |
|
| Korsgren O et al. [127] | 1990 | Iso (mouse) |
|
| Scharp DW et al. [100] | 1992 | Auto (dog) |
|
| Motojima K et al. [101] | 1992 | Auto (dog) |
|
| Marchetti P et al. [102] | 1993 | Auto (dog) |
|
| Ao Z et al. [56] | 1993 | Auto (dog) |
|
| Yakimets WJ et al. [117] | 1993 | Allo (dog) |
|
| Hesse UJ et al. [111] | 1994 | Auto (pig) |
|
| Eizirik DL et al. [128] | 1997 | Xeno, allo (human and mouse-to-nude mouse) |
|
| Horton PJ et al. [77] | 2000 | Allo (dog) |
|
| Authors [Reference] | Publication Year | Transplant model | Comments |
|---|---|---|---|
| Sutton R et al. [114] | 1989 | vs. Liver (auto, monkey) |
|
| Evans MG et al. [96] | 1989 | vs. Liver, kidney (auto, dog) |
|
| van der Vliet JA et al. [97,98] | 1989 | vs. Liver (auto, dog) |
|
| Warnock GL et al. [99] | 1990 | vs. Liver (auto, dog) |
|
| Scharp DW et al. [100] | 1992 | vs. Liver (auto, dog) |
|
| Motojima K et al. [101] | 1992 | vs. Liver (Auto, dog) |
|
| Ao Z et al. [56] | 1993 | vs. Omental pouch (auto, dog) |
|
| Hesse UJ et al. [111] | 1994 | vs. Liver (auto, pig) |
|
| Gustavson SM et al. [131] | 2005 | vs. Omental pouch (auto, dog) |
|
| Stokes RA et al. [129] | 2017 | vs. Liver, kidney (Allo, pig) |
|
| Stokes RA et al. [130] | 2017 | vs. Liver, kidney (iso, mouse) vs. Liver, kidney, portal vein, muscle (xeno, human-to-SCID mouse) |
|
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakata, N.; Yoshimatsu, G.; Kodama, S. The Spleen as an Optimal Site for Islet Transplantation and a Source of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2018, 19, 1391. https://doi.org/10.3390/ijms19051391
Sakata N, Yoshimatsu G, Kodama S. The Spleen as an Optimal Site for Islet Transplantation and a Source of Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2018; 19(5):1391. https://doi.org/10.3390/ijms19051391
Chicago/Turabian StyleSakata, Naoaki, Gumpei Yoshimatsu, and Shohta Kodama. 2018. "The Spleen as an Optimal Site for Islet Transplantation and a Source of Mesenchymal Stem Cells" International Journal of Molecular Sciences 19, no. 5: 1391. https://doi.org/10.3390/ijms19051391
APA StyleSakata, N., Yoshimatsu, G., & Kodama, S. (2018). The Spleen as an Optimal Site for Islet Transplantation and a Source of Mesenchymal Stem Cells. International Journal of Molecular Sciences, 19(5), 1391. https://doi.org/10.3390/ijms19051391