New Insight into the Octamer of TYMS Stabilized by Intermolecular Cys43-Disulfide


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
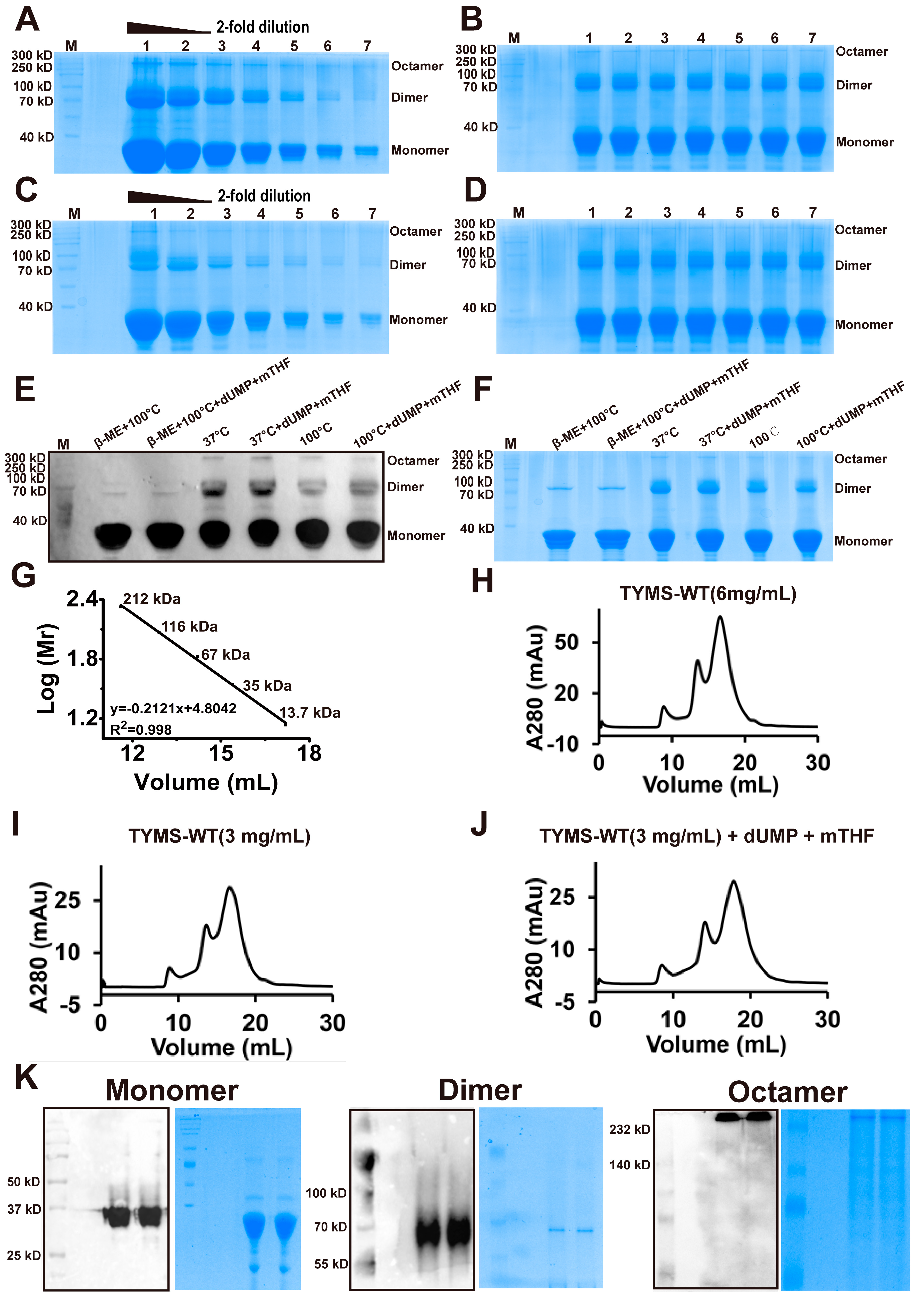
2.1. TYMS Overexpression and Purification
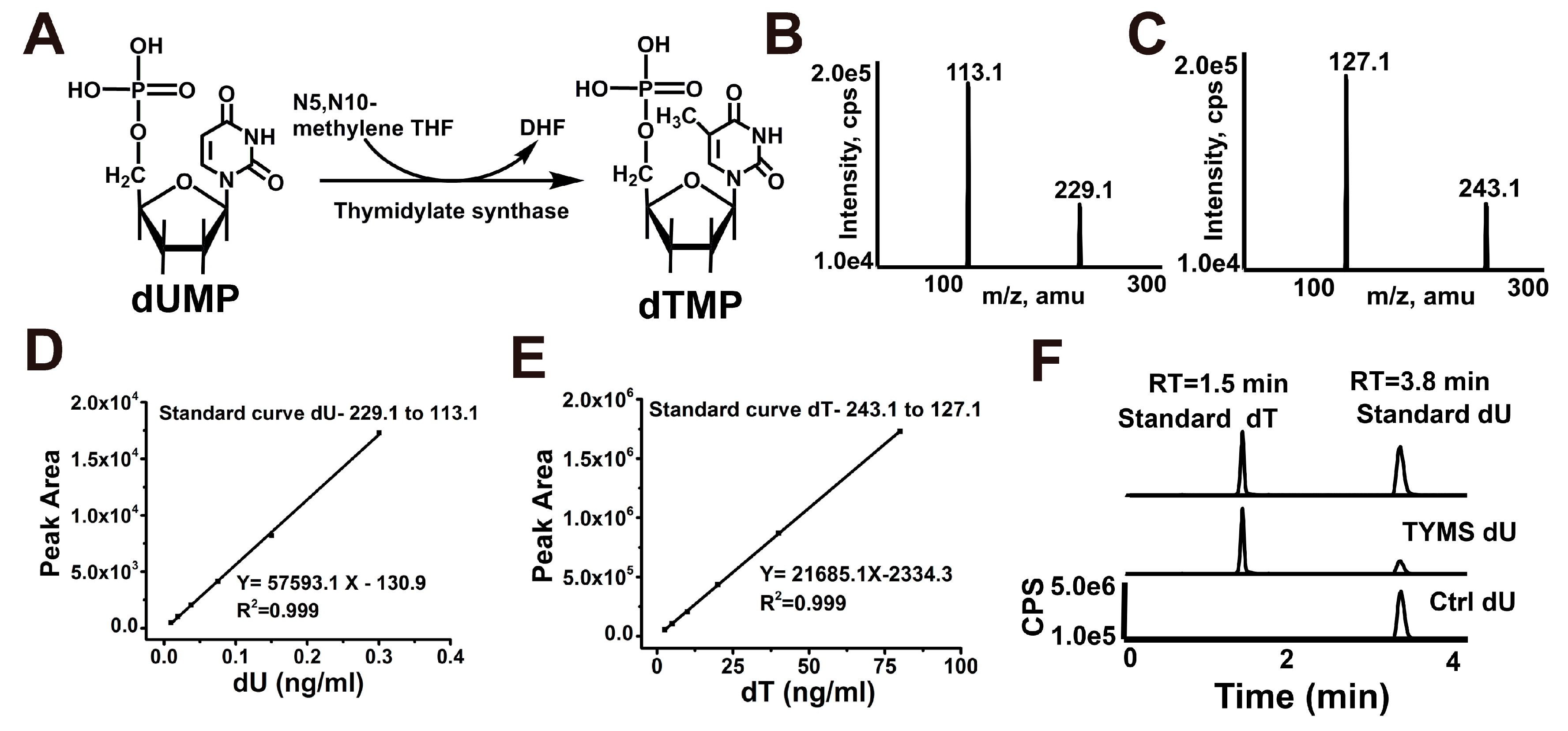
2.2. Reconstitution of TYMS-Mediated dTMP Synthesis
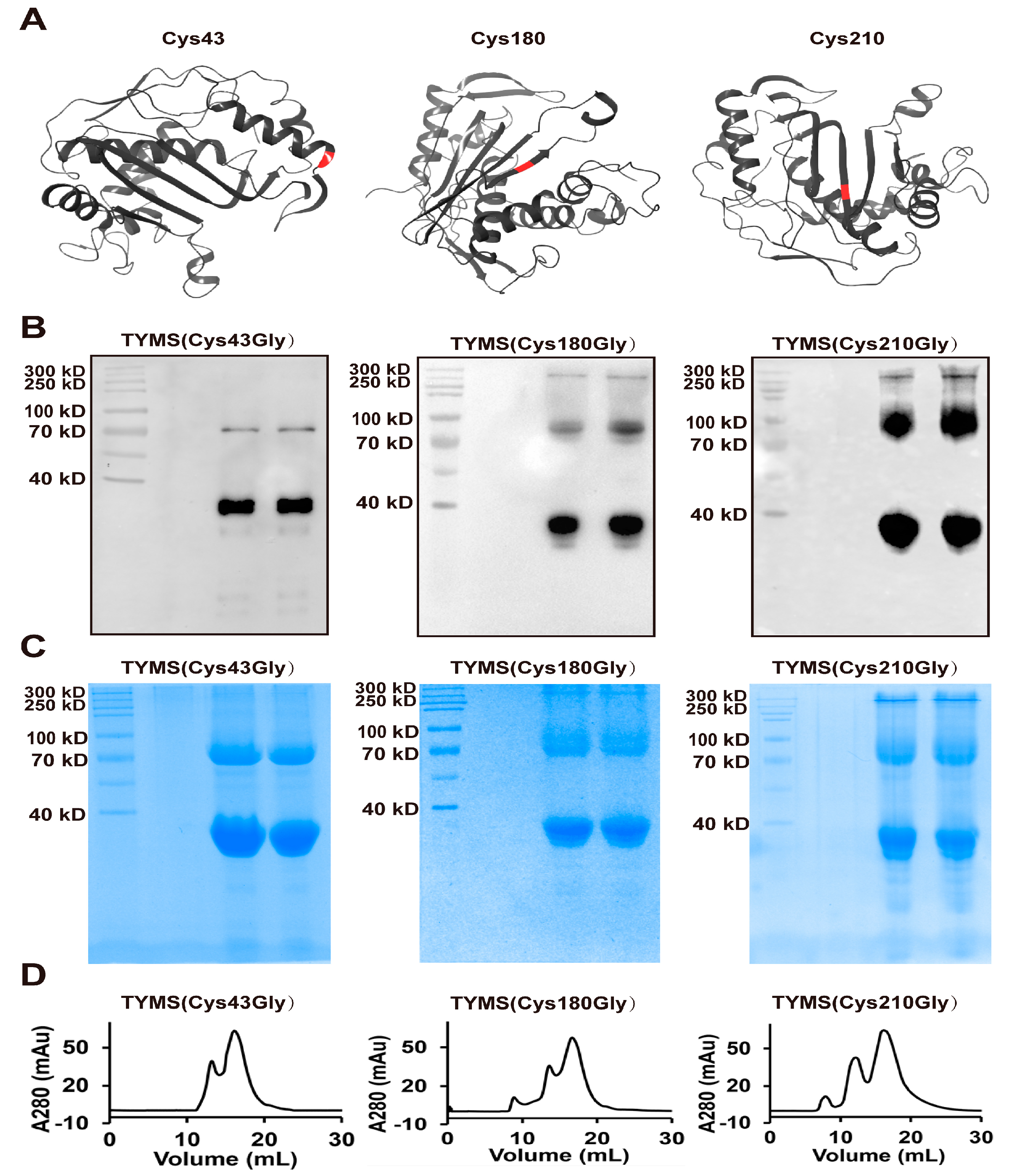
2.3. TYMS Formed Octamer by Intermolecular Cys43-Disulfide
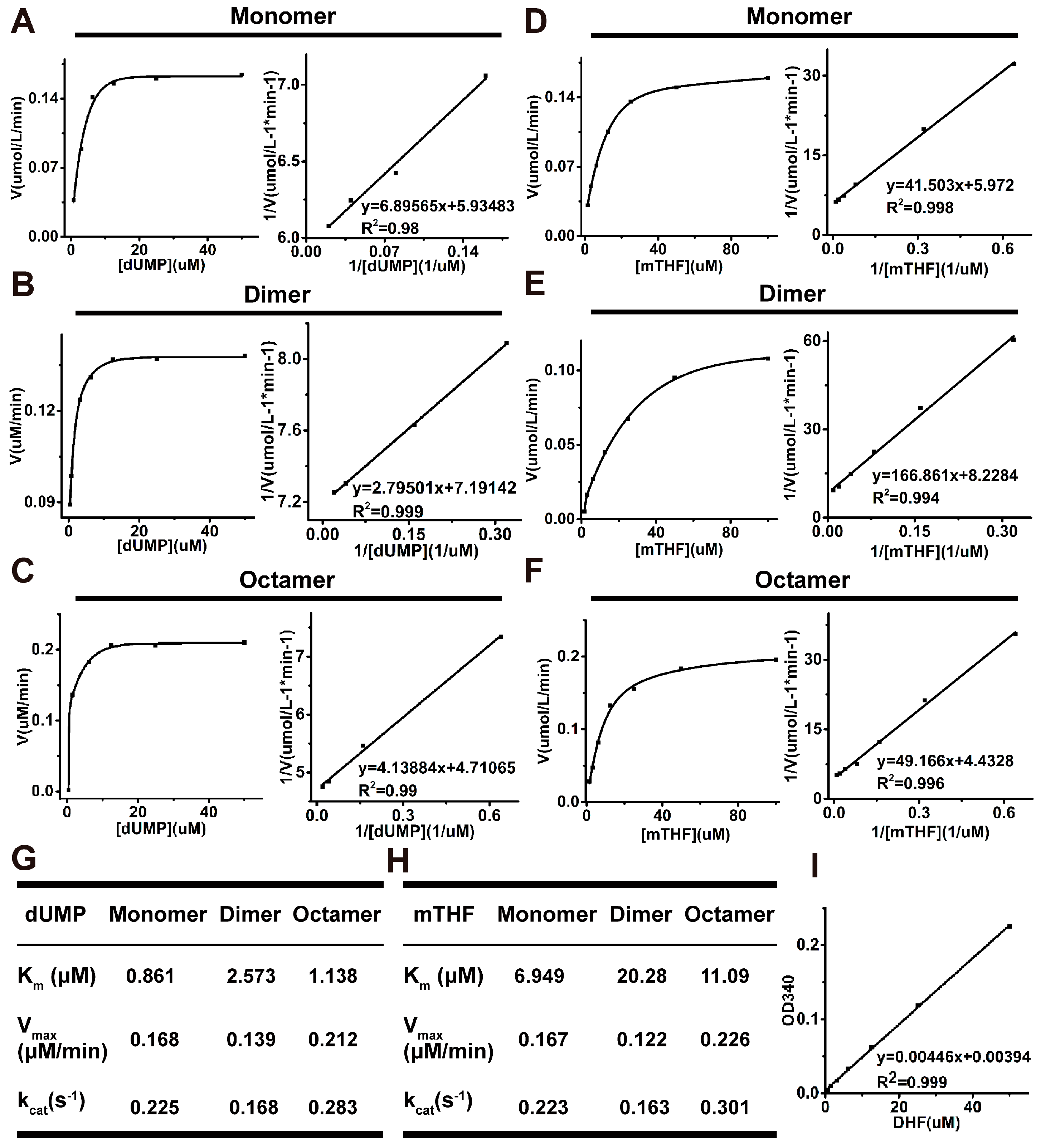
2.4. Analysis of Kinetic Properties of All Oligomeric Forms
3. Discussion
4. Materials and Methods
4.1. Plasmid Construction
4.2. Expression Screening
4.3. Protein Purification
4.4. SDS-PAGE, Western Blot, and Coomassie Brilliant Analysis
4.5. Activity Detection
4.6. Size Exclusion Chromatography
4.7. Reaction Kinetics Detection
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Choi, Y.M.; Yeo, H.K.; Park, Y.W.; Lee, J.Y. Structural Analysis of Thymidylate Synthase from Kaposi’s Sarcoma-Associated Herpesvirus with the Anticancer Drug Raltitrexed. PLoS ONE 2016, 11, e0168019. [Google Scholar] [CrossRef] [PubMed]
- Koehn, E.M.; Fleischmann, T.; Conrad, J.A.; Palfey, B.A.; Lesley, S.A.; Mathews, I.I.; Kohen, A. An unusual mechanism of thymidylate biosynthesis in organisms containing the thyX gene. Nature 2009, 458, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.R.; Guy, H.I. Mammalian pyrimidine biosynthesis: Fresh insights into an ancient pathway. J. Biol. Chem. 2004, 279, 33035–33038. [Google Scholar] [CrossRef] [PubMed]
- Carreras, C.W.; Santi, D.V. The catalytic mechanism and structure of thymidylate synthase. Annu. Rev. Biochem. 1995, 64, 721–762. [Google Scholar] [CrossRef] [PubMed]
- Finer-Moore, J.S.; Santi, D.V.; Stroud, R.M. Lessons and conclusions from dissecting the mechanism of a bisubstrate enzyme: Thymidylate synthase mutagenesis, function, and structure. Biochemistry 2003, 42, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Myllykallio, H.; Lipowski, G.; Leduc, D.; Filee, J.; Forterre, P.; Liebl, U. An alternative flavin-dependent mechanism for thymidylate synthesis. Science 2002, 297, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Mathews, I.I.; Deacon, A.M.; Canaves, J.M.; McMullan, D.; Lesley, S.A.; Agarwalla, S.; Kuhn, P. Functional analysis of substrate and cofactor complex structures of a thymidylate synthase-complementing protein. Structure 2003, 11, 677–690. [Google Scholar] [CrossRef]
- Lesley, S.A.; Kuhn, P.; Godzik, A.; Deacon, A.M.; Mathews, I.; Kreusch, A.; Spraggon, G.; Klock, H.E.; McMullan, D.; Shin, T.; et al. Structural genomics of the Thermotoga maritima proteome implemented in a high-throughput structure determination pipeline. Proc. Natl. Acad. Sci. USA 2002, 99, 11664–11669. [Google Scholar] [CrossRef] [PubMed]
- Leduc, D.; Graziani, S.; Meslet-Cladiere, L.; Sodolescu, A.; Liebl, U.; Myllykallio, H. Two distinct pathways for thymidylate (dTMP) synthesis in (hyper)thermophilic Bacteria and Archaea. Biochem. Soc. Trans. 2004, 32, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Mishanina, T.V.; Yu, L.; Karunaratne, K.; Mondal, D.; Corcoran, J.M.; Choi, M.A.; Kohen, A. An unprecedented mechanism of nucleotide methylation in organisms containing thyX. Science 2016, 351, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Papamichael, D. The use of thymidylate synthase inhibitors in the treatment of advanced colorectal cancer: Current status. Stem Cells 2000, 18, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Santhekadur, P.K.; Rajasekaran, D.; Siddiq, A.; Gredler, R.; Chen, D.; Schaus, S.E.; Hansen, U.; Fisher, P.B.; Sarkar, D. The transcription factor LSF: A novel oncogene for hepatocellular carcinoma. Am. J. Cancer Res. 2012, 2, 269–285. [Google Scholar] [PubMed]
- Wilson, P.M.; Danenberg, P.V.; Johnston, P.G.; Lenz, H.J.; Ladner, R.D. Standing the test of time: Targeting thymidylate biosynthesis in cancer therapy. Nat. Rev. Clin. Oncol. 2014, 11, 282–298. [Google Scholar] [CrossRef] [PubMed]
- Assaraf, Y.G. Molecular basis of antifolate resistance. Cancer Metastasis Rev. 2007, 26, 153–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Marsh, S.; Cassidy, J.; McLeod, H.L. Pharmacogenomic dissection of resistance to thymidylate synthase inhibitors. Cancer Res. 2001, 61, 5505–5510. [Google Scholar] [PubMed]
- Kitchens, M.E.; Forsthoefel, A.M.; Barbour, K.W.; Spencer, H.T.; Berger, F.G. Mechanisms of acquired resistance to thymidylate synthase inhibitors: The role of enzyme stability. Mol. Pharmacol. 1999, 56, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Phan, J.; Koli, S.; Minor, W.; Dunlap, R.B.; Berger, S.H.; Lebioda, L. Human thymidylate synthase is in the closed conformation when complexed with dUMP and raltitrexed, an antifolate drug. Biochemistry 2001, 40, 1897–1902. [Google Scholar] [CrossRef] [PubMed]
- Genovese, F.; Ferrari, S.; Guaitoli, G.; Caselli, M.; Costi, M.P.; Ponterini, G. Dimer-monomer equilibrium of human thymidylate synthase monitored by fluorescence resonance energy transfer. Protein Sci. 2010, 19, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, C.A.; Clifton, I.J.; Davisson, V.J.; Santi, D.V.; Stroud, R.M. Crystal structure of human thymidylate synthase: A structural mechanism for guiding substrates into the active site. Biochemistry 1995, 34, 16279–16287. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Liu, J.; Maley, F.; Chu, E. Role of cysteine amino acid residues on the RNA binding activity of human thymidylate synthase. Nucleic Acids Res. 2003, 31, 4882–4887. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.H.; Berger, F.G.; Lebioda, L. Effects of ligand binding and conformational switching on intracellular stability of human thymidylate synthase. Biochim. Biophys. Acta 2004, 1696, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Voeller, D.M.; Zajac-Kaye, M.; Fisher, R.J.; Allegra, C.J. The identification of thymidylate synthase peptide domains located in the interface region that bind thymidylate synthase mRNA. Biochem. Biophys. Res. Commun. 2002, 297, 24–31. [Google Scholar] [CrossRef]
- Chu, E.; Voeller, D.; Koeller, D.M.; Drake, J.C.; Takimoto, C.H.; Maley, G.F.; Maley, F.; Allegra, C.J. Identification of an RNA binding site for human thymidylate synthase. Proc. Natl. Acad. Sci. USA 1993, 90, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Chu, E.; Koeller, D.M.; Casey, J.L.; Drake, J.C.; Chabner, B.A.; Elwood, P.C.; Zinn, S.; Allegra, C.J. Autoregulation of human thymidylate synthase messenger RNA translation by thymidylate synthase. Proc. Natl. Acad. Sci. USA 1991, 88, 8977–8981. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; O’Neil, R.H.; Surti, T.S.; Stroud, R.M. Approaches to solving the rigid receptor problem by identifying a minimal set of flexible residues during ligand docking. Chem. Biol. 2001, 8, 445–457. [Google Scholar] [CrossRef]
- Costi, M.P.; Ferrari, S.; Venturelli, A.; Calo, S.; Tondi, D.; Barlocco, D. Thymidylate synthase structure, function and implication in drug discovery. Curr. Med. Chem. 2005, 12, 2241–2258. [Google Scholar] [CrossRef] [PubMed]
- Phan, J.; Mahdavian, E.; Nivens, M.C.; Minor, W.; Berger, S.; Spencer, H.T.; Dunlap, R.B.; Lebioda, L. Catalytic cysteine of thymidylate synthase is activated upon substrate binding. Biochemistry 2000, 39, 6969–6978. [Google Scholar] [CrossRef] [PubMed]
- Almog, R.; Waddling, C.A.; Maley, F.; Maley, G.F.; Van Roey, P. Crystal structure of a deletion mutant of human thymidylate synthase Delta (7–29) and its ternary complex with Tomudex and dUMP. Protein Sci. 2001, 10, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Phan, J.; Steadman, D.J.; Koli, S.; Ding, W.C.; Minor, W.; Dunlap, R.B.; Berger, S.H.; Lebioda, L. Structure of human thymidylate synthase suggests advantages of chemotherapy with noncompetitive inhibitors. J. Biol. Chem. 2001, 276, 14170–14177. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; McCammon, J.A. Substrate channeling between the human dihydrofolate reductase and thymidylate synthase. Protein Sci. 2016, 25, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Jansson, A.; Sim, D.; Larsson, A.; Nordlund, P. Structural analyses of human thymidylate synthase reveal a site that may control conformational switching between active and inactive states. J. Biol. Chem. 2017, 292, 13449–13458. [Google Scholar] [CrossRef] [PubMed]
- Salo-Ahen, O.M.; Tochowicz, A.; Pozzi, C.; Cardinale, D.; Ferrari, S.; Boum, Y.; Mangani, S.; Stroud, R.M.; Saxena, P.; Myllykallio, H.; et al. Hotspots in an obligate homodimeric anticancer target. Structural and functional effects of interfacial mutations in human thymidylate synthase. J. Med. Chem. 2015, 58, 3572–3581. [Google Scholar] [CrossRef] [PubMed]
- Cella, R.; Carbonera, D.; Orsi, R.; Ferri, G.; Iadarola, P. Proteolytic and partial sequencing studies of the bifunctional dihydrofolate reductase-thymidylate synthase from Daucus carota. Plant Mol. Biol. 1991, 16, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Dowiercial, A.; Wilk, P.; Rypniewski, W.; Rode, W.; Jarmula, A. Crystal structure of mouse thymidylate synthase in tertiary complex with dUMP and raltitrexed reveals N-terminus architecture and two different active site conformations. Biomed. Res. Int. 2014, 2014, 945803. [Google Scholar] [CrossRef] [PubMed]
- Cha-Aim, K.; Hoshida, H.; Fukunaga, T.; Akada, R. Fusion PCR via novel overlap sequences. Methods Mol. Biol. 2012, 852, 97–110. [Google Scholar] [PubMed]
- Wang, L.; Quan, C.; Liu, B.; Xu, Y.; Zhao, P.; Xiong, W.; Fan, S. Green fluorescent protein (GFP)-based overexpression screening and characterization of AgrC, a Receptor protein of quorum sensing in Staphylococcus aureus. Int. J. Mol. Sci. 2013, 14, 18470–18487. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhou, G.; Yu, X.; Xu, Q.; Wang, K.; Xie, D.; Yang, Q.; Wang, L. LC-MS-MS quantitative analysis reveals the association between FTO and DNA methylation. PLoS ONE 2017, 12, e0175849. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, R.B.; Harding, N.G.; Huennekens, F.M. Thymidylate synthetase from amethopterin-resistant Lactobacillus casei. Biochemistry 1971, 10, 88–97. [Google Scholar] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, D.; Wang, L.; Xiao, Q.; Wu, X.; Zhang, L.; Yang, Q.; Wang, L. New Insight into the Octamer of TYMS Stabilized by Intermolecular Cys43-Disulfide. Int. J. Mol. Sci. 2018, 19, 1393. https://doi.org/10.3390/ijms19051393
Xie D, Wang L, Xiao Q, Wu X, Zhang L, Yang Q, Wang L. New Insight into the Octamer of TYMS Stabilized by Intermolecular Cys43-Disulfide. International Journal of Molecular Sciences. 2018; 19(5):1393. https://doi.org/10.3390/ijms19051393
Chicago/Turabian StyleXie, Dan, Lulu Wang, Qi Xiao, Xiaoyan Wu, Lin Zhang, Qingkai Yang, and Lina Wang. 2018. "New Insight into the Octamer of TYMS Stabilized by Intermolecular Cys43-Disulfide" International Journal of Molecular Sciences 19, no. 5: 1393. https://doi.org/10.3390/ijms19051393
APA StyleXie, D., Wang, L., Xiao, Q., Wu, X., Zhang, L., Yang, Q., & Wang, L. (2018). New Insight into the Octamer of TYMS Stabilized by Intermolecular Cys43-Disulfide. International Journal of Molecular Sciences, 19(5), 1393. https://doi.org/10.3390/ijms19051393