Overexpression of CBX3 in Pancreatic Adenocarcinoma Promotes Cell Cycle Transition-Associated Tumor Progression

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
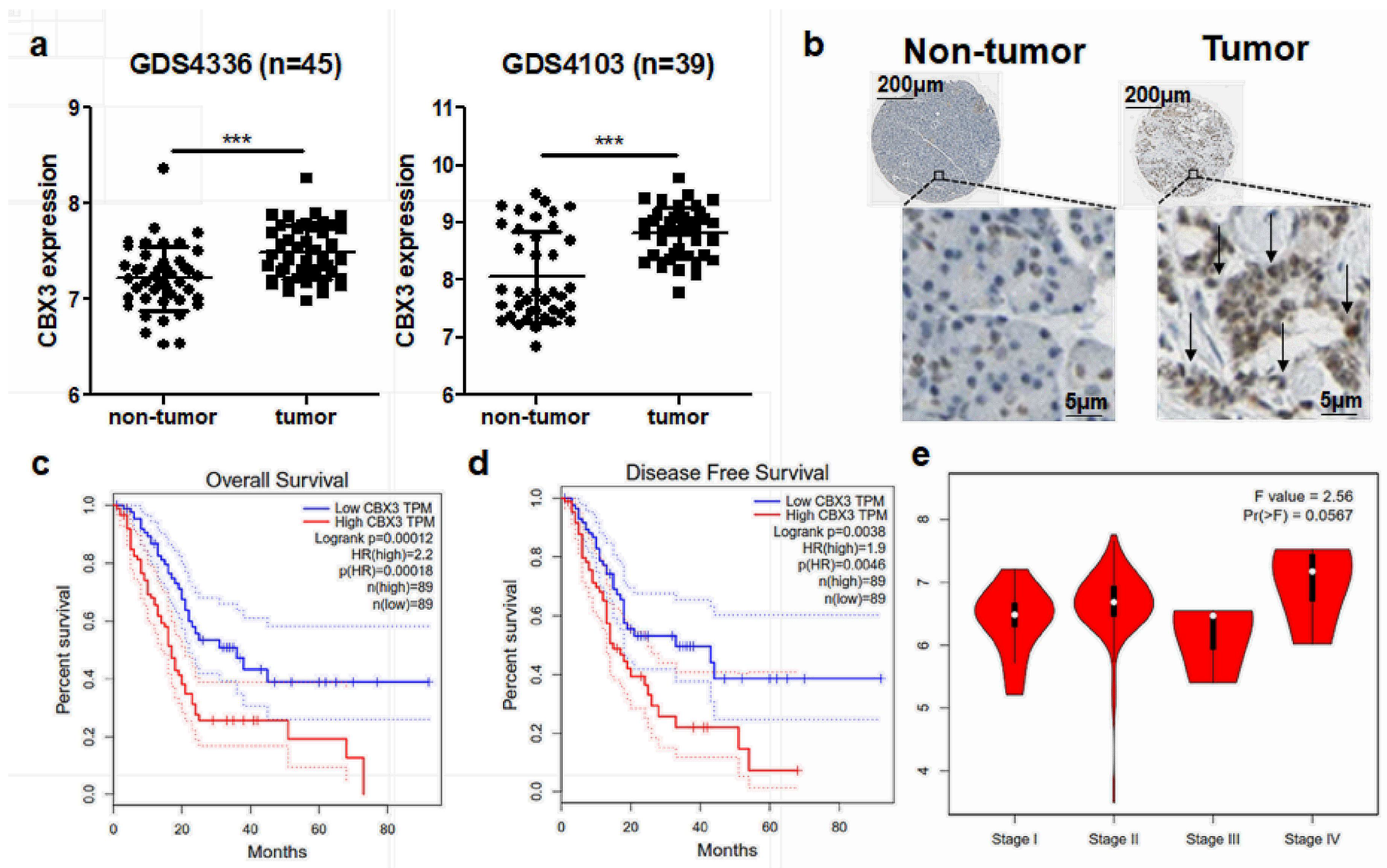
2.1. CBX3 Was Overexpressed in PAAD and Predicted Poor Prognosis of Patients
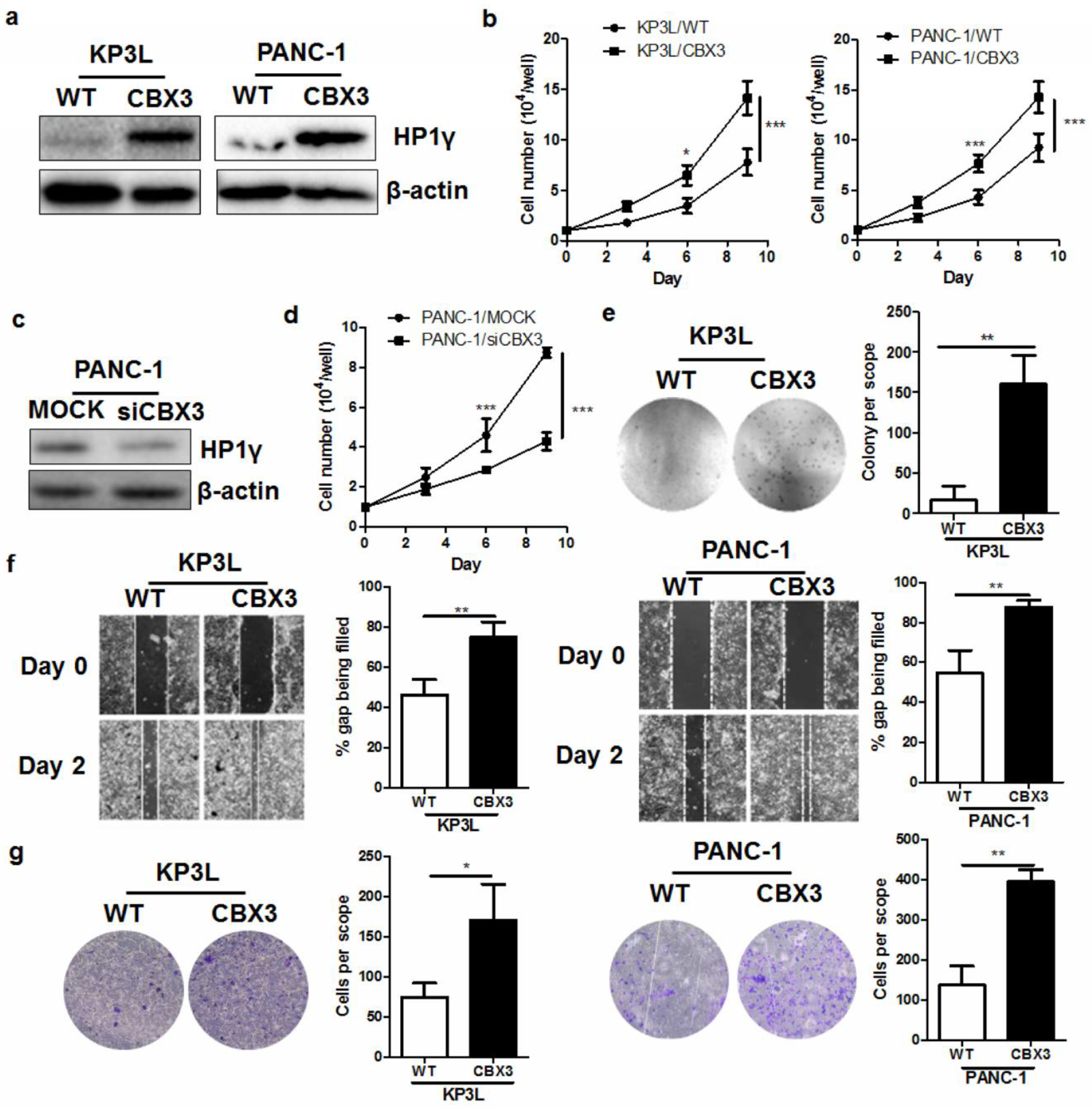
2.2. CBX3 Promoted the In Vitro Proliferation and Invasiveness of PAAD Cells
2.3. CBX3 Overexpression Accelerated In Vivo Tumor Progression of PAAD
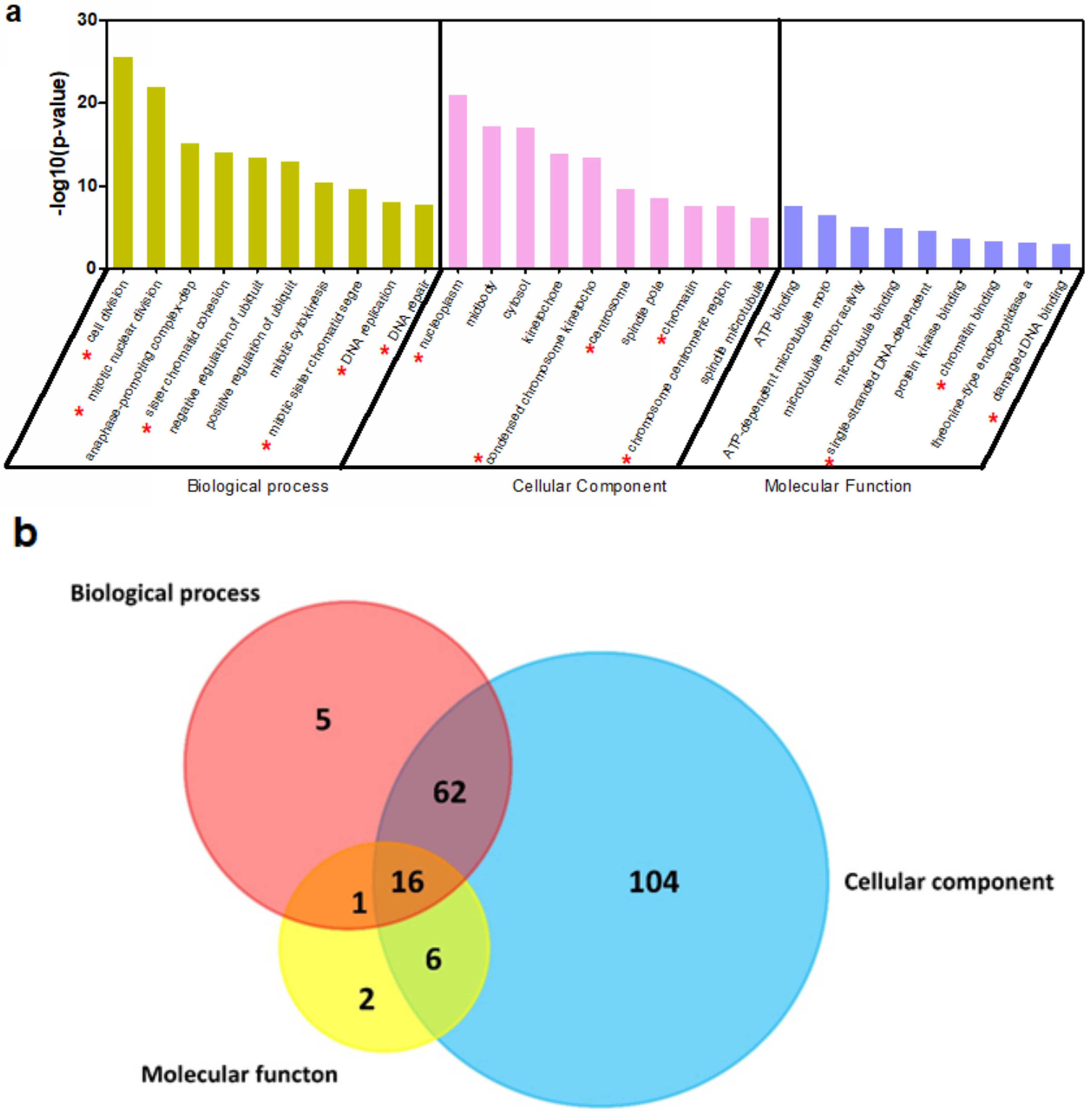
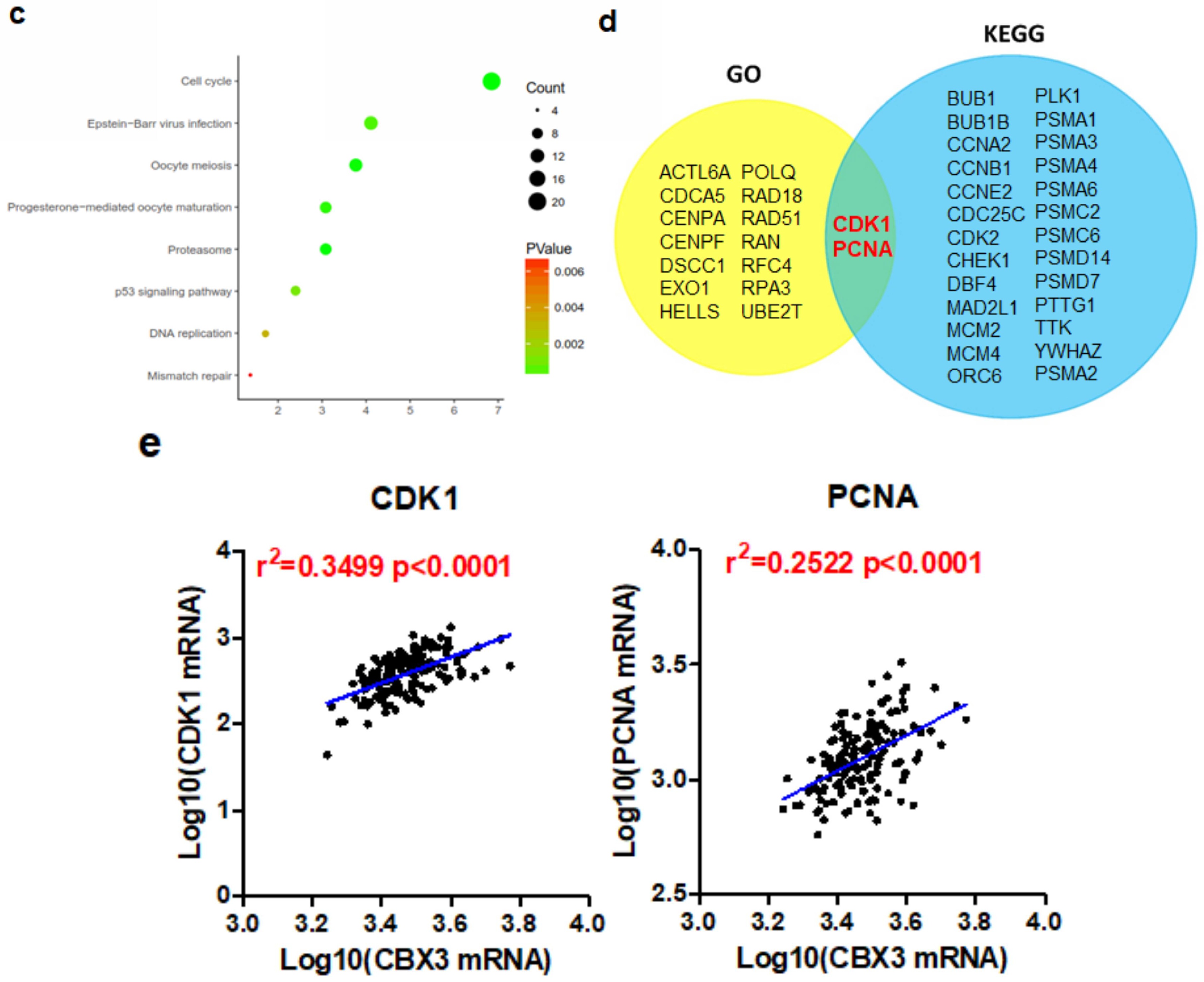
2.4. CBX3 May Be Involved in the Regulation of Cell Cycle Progression of PAAD
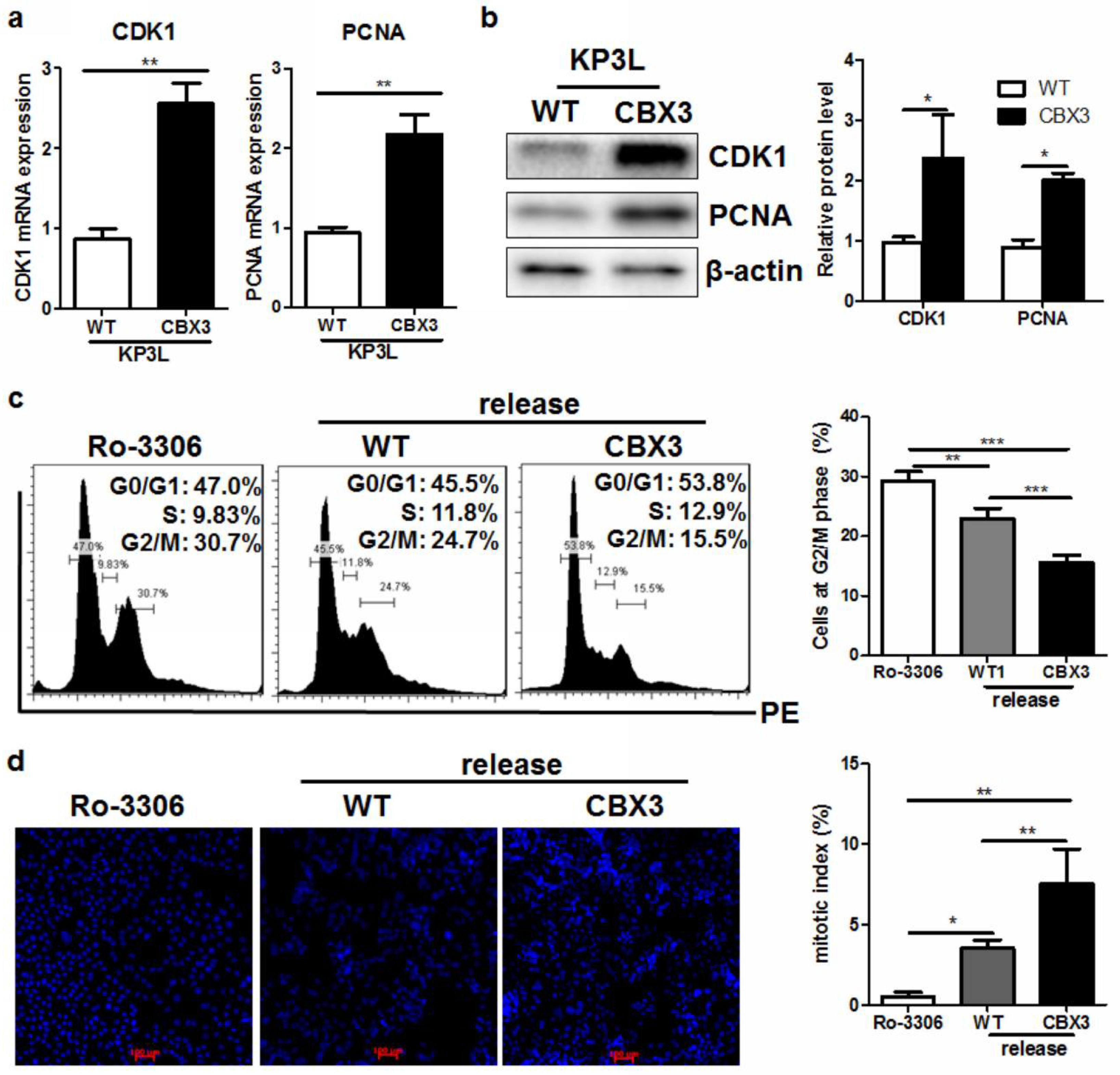
2.5. CBX3 May Be Involved in the Regulation of G2/M Transition of PAAD Cells
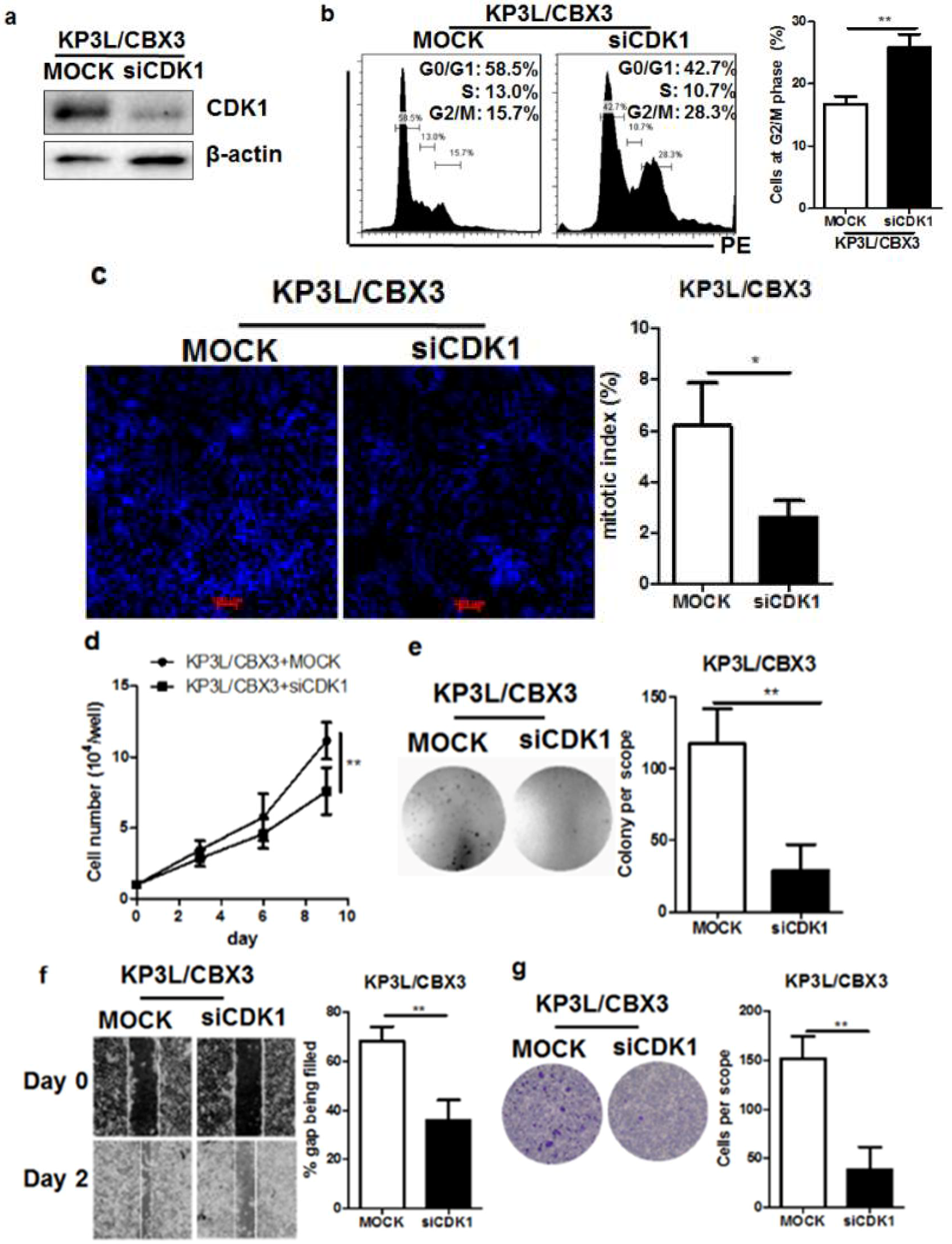
2.6. The Regulation of Cell Cycle Transition by CBX3 Is Mediated by CDK1
3. Discussion
4. Materials and Methods
4.1. Chemicals, Antibodies, and Plasmids
4.2. Cells and Cell Culture
4.3. Animal Study Protocol
4.4. Soft Agar Assay
4.5. Wound Healing and Invasion Chamber Assay
4.6. Quantitative Real-Time PCR
4.7. Immunoblotting
4.8. Quantification of Mitotic Index
4.9. Statistical Analysis
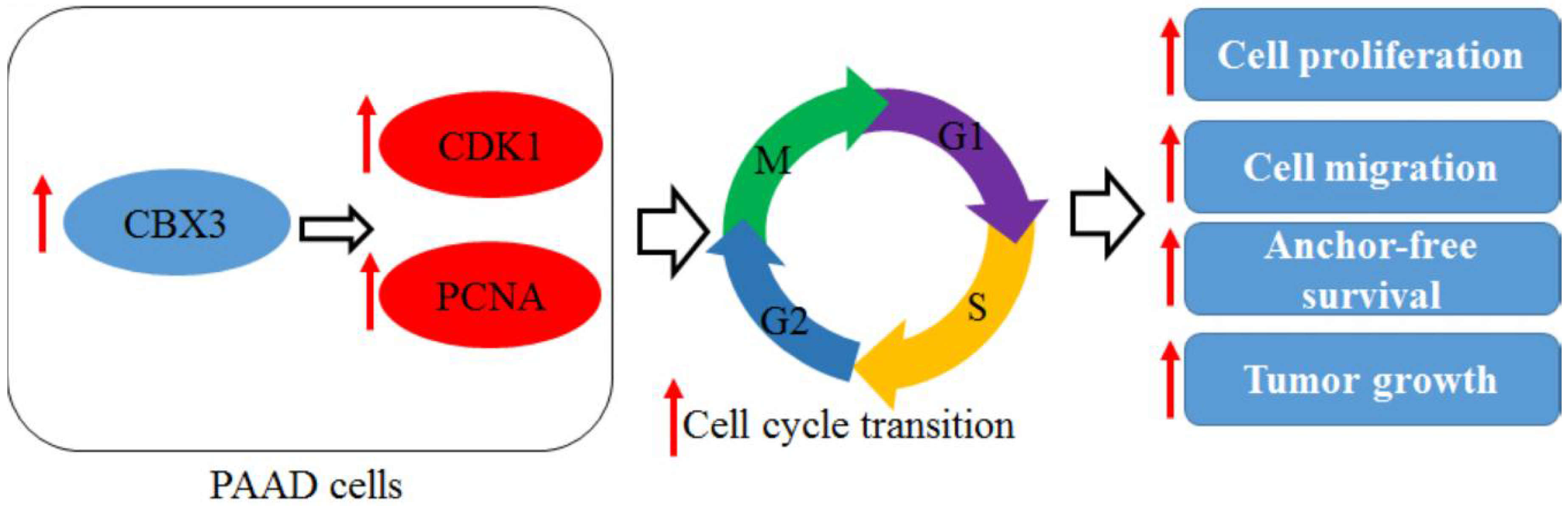
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ghaneh, P.; Costello, E.; Neoptolemos, J.P. Biology and management of pancreatic cancer. Gut 2007, 56, 1134–1152. [Google Scholar] [CrossRef] [PubMed]
- Higuera, O.; Ghanem, I.; Nasimi, R.; Prieto, I.; Koren, L.; Feliu, J. Management of pancreatic cancer in the elderly. World J. Gastroenterol. 2016, 22, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Crippa, S.; Capurso, G.; Camma, C.; Fave, G.D.; Castillo, C.F.; Falconi, M. Risk of pancreatic malignancy and mortality in branch-duct ipmns undergoing surveillance: A systematic review and meta-analysis. Dig. Liver Dis. 2016, 48, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the united states. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed]
- Quante, A.S.; Ming, C.; Rottmann, M.; Engel, J.; Boeck, S.; Heinemann, V.; Westphalen, C.B.; Strauch, K. Projections of cancer incidence and cancer-related deaths in germany by 2020 and 2030. Cancer Med. 2016, 5, 2649–2656. [Google Scholar] [CrossRef] [PubMed]
- Pandol, S.; Gukovskaya, A.; Edderkaoui, M.; Dawson, D.; Eibl, G.; Lugea, A. Epidemiology, risk factors, and the promotion of pancreatic cancer: Role of the stellate cell. J. Gastroenterol. Hepatol. 2012, 27 (Suppl. 2), 127–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, R.E.; Greer, J.B.; Zolotarevsky, E.; Brand, R.; Du, H.; Simeone, D.; Zisman, A.; Gorchow, A.; Lee, S.Y.; Roy, H.K.; et al. Pancreatic cancer patients who smoke and drink are diagnosed at younger ages. Clin. Gastroenterol. Hepatol. 2009, 7, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Kim, V.M.; Ahuja, N. Early detection of pancreatic cancer. Chin. J. Cancer Res. 2015, 27, 321–331. [Google Scholar] [PubMed]
- Marrocchio, C.; Dababou, S.; Catalano, C.; Napoli, A. Nonoperative ablation of pancreatic neoplasms. Surg. Clin. N. Am. 2018, 98, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Zhang, Y.; Yu, X.; Yang, J.; LeBrun, D.G.; Chen, C.; Yao, Q.; Li, M. Overcoming drug resistance in pancreatic cancer. Expert Opin. Ther. Targets 2011, 15, 817–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slezak, J.; Truong, M.; Huang, W.; Jarrard, D. Hp1gamma expression is elevated in prostate cancer and is superior to gleason score as a predictor of biochemical recurrence after radical prostatectomy. BMC Cancer 2013, 13, 148. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huang, F.; Zhang, D.; Ju, J.; Wu, X.B.; Wang, Y.; Wang, Y.; Wu, Y.; Nie, M.; Li, Z.; et al. Heterochromatin protein hp1gamma promotes colorectal cancer progression and is regulated by miR-30a. Cancer Res. 2015, 75, 4593–4604. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Li, H.; Liang, X.; Xiang, Z. CBX3 promotes colon cancer cell proliferation by CDK6 kinase-independent function during cell cycle. Oncotarget 2017, 8, 19934–19946. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, W.; Fu, X.; Su, X.; Yang, A. CBX3 promotes tumor proliferation by regulating G1/S phase via p21 downregulation and associates with poor prognosis in tongue squamous cell carcinoma. Gene 2018. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Lai, Y.C.; Chen, Y.C.; Wang, N.K.; Wang, W.S.; Lai, J.I. CBX3/heterochromatin protein 1 gamma is significantly upregulated in patients with non-small cell lung cancer. Asia-Pac. J. Clin. Oncol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.K.; Lin, H.Y.; Chen, C.F.; Zeng, D. Prognostic values of distinct CBX family members in breast cancer. Oncotarget 2017, 8, 92375–92387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Ha, N.; Pham, D.H.; Frederick, M.; Sharma, B.; Naruse, C.; Asano, M.; Pipkin, M.E.; George, R.E.; Thai, T.H. CBX3/HP1gamma deficiency confers enhanced tumor-killing capacity on CD8+ t cells. Sci. Rep. 2017, 7, 42888. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Kaldis, P. Cdks, cyclins and ckis: Roles beyond cell cycle regulation. Development 2013, 140, 3079–3093. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Han, S.X.; Ma, J.L.; Ying, X.; Liu, P.; Li, J.; Wang, L.; Zhang, Y.; Ma, J.; Zhang, L.; et al. The role of cdk1 in apoptin-induced apoptosis in hepatocellular carcinoma cells. Oncol. Rep. 2013, 30, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Hongo, F.; Takaha, N.; Oishi, M.; Ueda, T.; Nakamura, T.; Naitoh, Y.; Naya, Y.; Kamoi, K.; Okihara, K.; Matsushima, T.; et al. CDK1 and CDK2 activity is a strong predictor of renal cell carcinoma recurrence. Urol. Oncol. 2014, 32, 1240–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.Y.; Auerbach, A.; D'Costa, A.M.; Rapoport, A.P.; Burger, A.M.; Sausville, E.A.; Stass, S.A.; Jiang, F.; Sands, A.M.; Aguilera, N.; et al. Phospho-p70s6k/p85s6k and cdc2/cdk1 are novel targets for diffuse large b-cell lymphoma combination therapy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2009, 15, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, C.; Wang, X.; Becker, D. Expression analysis and molecular targeting of cyclin-dependent kinases in advanced melanoma. Cell Cycle 2011, 10, 977–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strzalka, W.; Ziemienowicz, A. Proliferating cell nuclear antigen (PCNA): A key factor in DNA replication and cell cycle regulation. Ann. Bot. 2011, 107, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Saksouk, N.; Simboeck, E.; Dejardin, J. Constitutive heterochromatin formation and transcription in mammals. Epigenet. Chromatin 2015, 8, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Jia, S.T.; Jia, S. New insights into the regulation of heterochromatin. Trends Genet. 2016, 32, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Canzio, D.; Larson, A.; Narlikar, G.J. Mechanisms of functional promiscuity by HP1 proteins. Trends Cell Biol. 2014, 24, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dialynas, G.K.; Vitalini, M.W.; Wallrath, L.L. Linking heterochromatin protein 1 (HP1) to cancer progression. Mutat. Res. 2008, 647, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Lieberthal, J.G.; Kaminsky, M.; Parkhurst, C.N.; Tanese, N. The role of YY1 in reduced HP1alpha gene expression in invasive human breast cancer cells. Breast Cancer Res. 2009, 11, R42. [Google Scholar] [CrossRef] [PubMed]
- Pongas, G.; Kim, M.K.; Min, D.J.; House, C.D.; Jordan, E.; Caplen, N.; Chakka, S.; Ohiri, J.; Kruhlak, M.J.; Annunziata, C.M. BRD4 facilitates DNA damage response and represses CBX5/heterochromatin protein 1 (HP1). Oncotarget 2017, 8, 51402–51415. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.D.; Park, M.A.; Lee, J.S. Suppression and recovery of brca1-mediated transcription by HP1gamma via modulation of promoter occupancy. Nucleic Acids Res. 2012, 40, 11321–11338. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, R.; Gonzales-Cope, M.; Chronis, C.; Bonora, G.; McKee, R.; Huang, C.; Patel, S.; Lopez, D.; Mishra, N.; Pellegrini, M.; et al. Proteomic and genomic approaches reveal critical functions of h3k9 methylation and heterochromatin protein-1gamma in reprogramming to pluripotency. Nat. Cell Biol. 2013, 15, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, T.; Calderwood, S.K.; Takigawa, M.; Kubota, S.; Kozaki, K.I. Intracellular MMP3 promotes hsp gene expression in collaboration with chromobox proteins. J. Cell. Biochem. 2017, 118, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, A.; Hon, G.C.; Jin, F.; Henry, R.E.; Espinosa, J.M.; Ren, B. CBX3 regulates efficient rna processing genome-wide. Genome Res. 2012, 22, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Salton, M.; Voss, T.C.; Misteli, T. Identification by high-throughput imaging of the histone methyltransferase EHMT2 as an epigenetic regulator of VEGFA alternative splicing. Nucleic Acids Res. 2014, 42, 13662–13673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigg, E.A. Mitotic kinases as regulators of cell division and its checkpoints. Nat. Rev. Mol. Cell Biol. 2001, 2, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, D.; Barriere, C.; Cerqueira, A.; Hunt, S.; Tardy, C.; Newton, K.; Caceres, J.F.; Dubus, P.; Malumbres, M.; Barbacid, M. CDK1 is sufficient to drive the mammalian cell cycle. Nature 2007, 448, 811–815. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Takeda, T.; Wakasa, K.; Tsujimoto, M.; Okada, M.; Matsuura, N. Expression of the G2-M modulators in pancreatic adenocarcinoma. Pancreatology 2002, 2, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, G.; Mishra, A.; Bisht, S.; Karikari, C.; Garrido-Laguna, I.; Rasheed, Z.; Ottenhof, N.A.; Dadon, T.; Alvarez, H.; Fendrich, V.; et al. Cyclin-dependent kinase inhibitor dinaciclib (SCH727965) inhibits pancreatic cancer growth and progression in murine xenograft models. Cancer Biol. Ther. 2011, 12, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Catania, M.G.; Mischel, P.S.; Vinters, H.V. Hamartin and tuberin interaction with the G2/M cyclin-dependent kinase CDK1 and its regulatory cyclins A and B. J. Neuropathol. Exp. Neurol. 2001, 60, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Pu, S.; Zhao, Y.; Zhou, G.; Zhu, H.; Gong, L.; Zhang, W.; Huang, G.; Wang, D.; Liu, D. Effect of CDK1 shRNA on proliferation, migration, cell cycle and apoptosis in non-small cell lung cancer. J. Cell. Physiol. 2017. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, L.; Liu, M.; Chong, R.; Ding, S.J.; Chen, Y.; Dong, J. CDK1 phosphorylation of YAP promotes mitotic defects and cell motility and is essential for neoplastic transformation. Cancer Res. 2013, 73, 6722–6733. [Google Scholar] [CrossRef] [PubMed]
- Warfel, N.A.; Dolloff, N.G.; Dicker, D.T.; Malysz, J.; El-Deiry, W.S. Cdk1 stabilizes hif-1alpha via direct phosphorylation of ser668 to promote tumor growth. Cell Cycle 2013, 12, 3689–3701. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Stauffer, S.; Zhou, J.; Chen, X.; Chen, Y.; Dong, J. Cyclin-dependent kinase 1 (CDK1)-mediated mitotic phosphorylation of the transcriptional co-repressor Vgll4 inhibits its tumor-suppressing activity. J. Biol. Chem. 2017, 292, 15028–15038. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.A.; Sanchez, A.; Hoffman, R.M.; Nur, S.; Lambros, M.P. Fluorescent orthotopic mouse model of pancreatic cancer. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jin, D.Y.; Ng, R.W.; Feng, H.; Wong, Y.C.; Cheung, A.L.; Tsao, S.W. Significance of MAD2 expression to mitotic checkpoint control in ovarian cancer cells. Cancer Res. 2002, 62, 1662–1668. [Google Scholar] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.-Y.; Cheng, C.-S.; Qu, C.; Wang, P.; Chen, H.; Meng, Z.-Q.; Chen, Z. Overexpression of CBX3 in Pancreatic Adenocarcinoma Promotes Cell Cycle Transition-Associated Tumor Progression. Int. J. Mol. Sci. 2018, 19, 1768. https://doi.org/10.3390/ijms19061768
Chen L-Y, Cheng C-S, Qu C, Wang P, Chen H, Meng Z-Q, Chen Z. Overexpression of CBX3 in Pancreatic Adenocarcinoma Promotes Cell Cycle Transition-Associated Tumor Progression. International Journal of Molecular Sciences. 2018; 19(6):1768. https://doi.org/10.3390/ijms19061768
Chicago/Turabian StyleChen, Lian-Yu, Chien-Shan Cheng, Chao Qu, Peng Wang, Hao Chen, Zhi-Qiang Meng, and Zhen Chen. 2018. "Overexpression of CBX3 in Pancreatic Adenocarcinoma Promotes Cell Cycle Transition-Associated Tumor Progression" International Journal of Molecular Sciences 19, no. 6: 1768. https://doi.org/10.3390/ijms19061768
APA StyleChen, L.-Y., Cheng, C.-S., Qu, C., Wang, P., Chen, H., Meng, Z.-Q., & Chen, Z. (2018). Overexpression of CBX3 in Pancreatic Adenocarcinoma Promotes Cell Cycle Transition-Associated Tumor Progression. International Journal of Molecular Sciences, 19(6), 1768. https://doi.org/10.3390/ijms19061768