Transcriptome Analyses in Different Cucumber Cultivars Provide Novel Insights into Drought Stress Responses

Abstract
:1. Introduction
2. Results
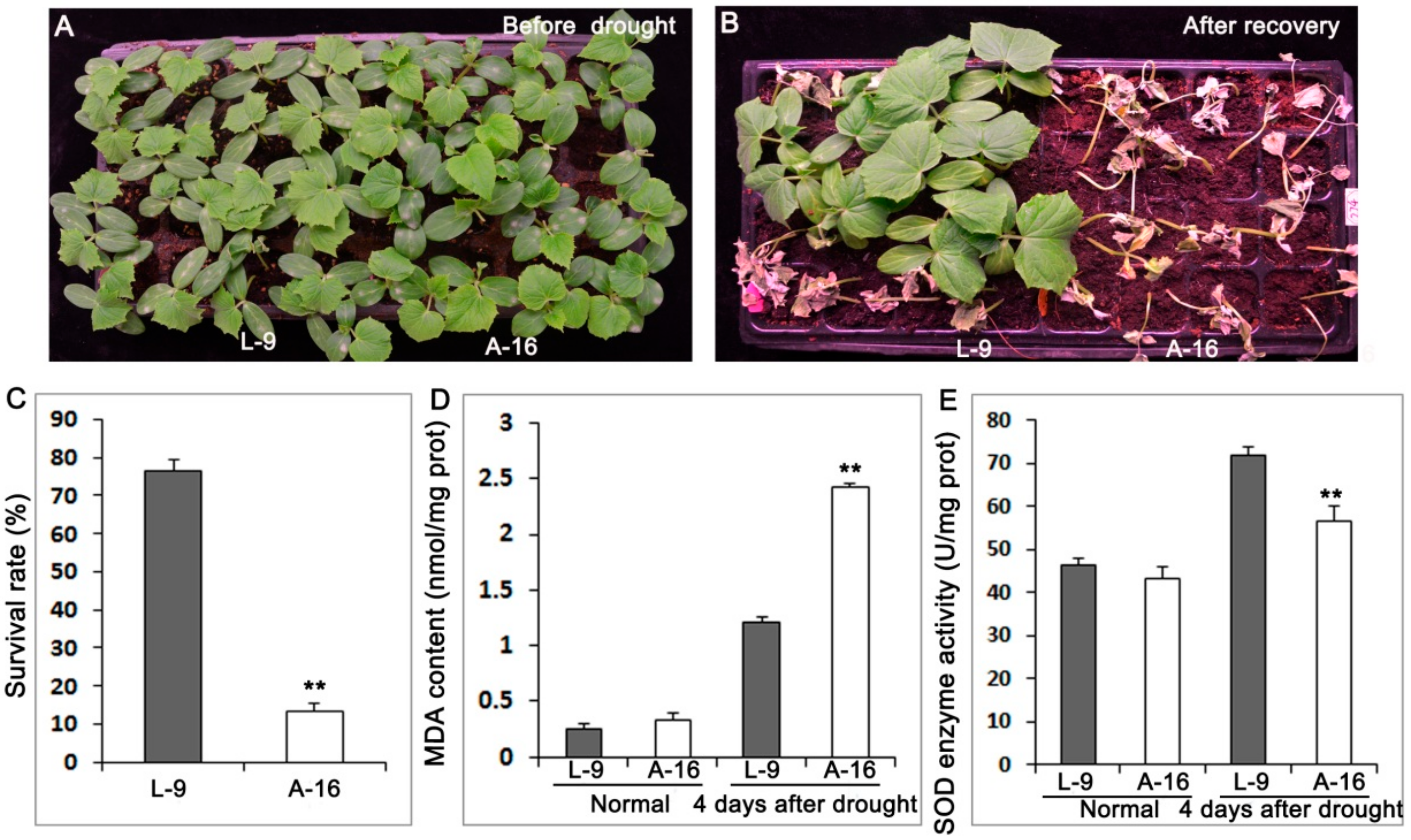
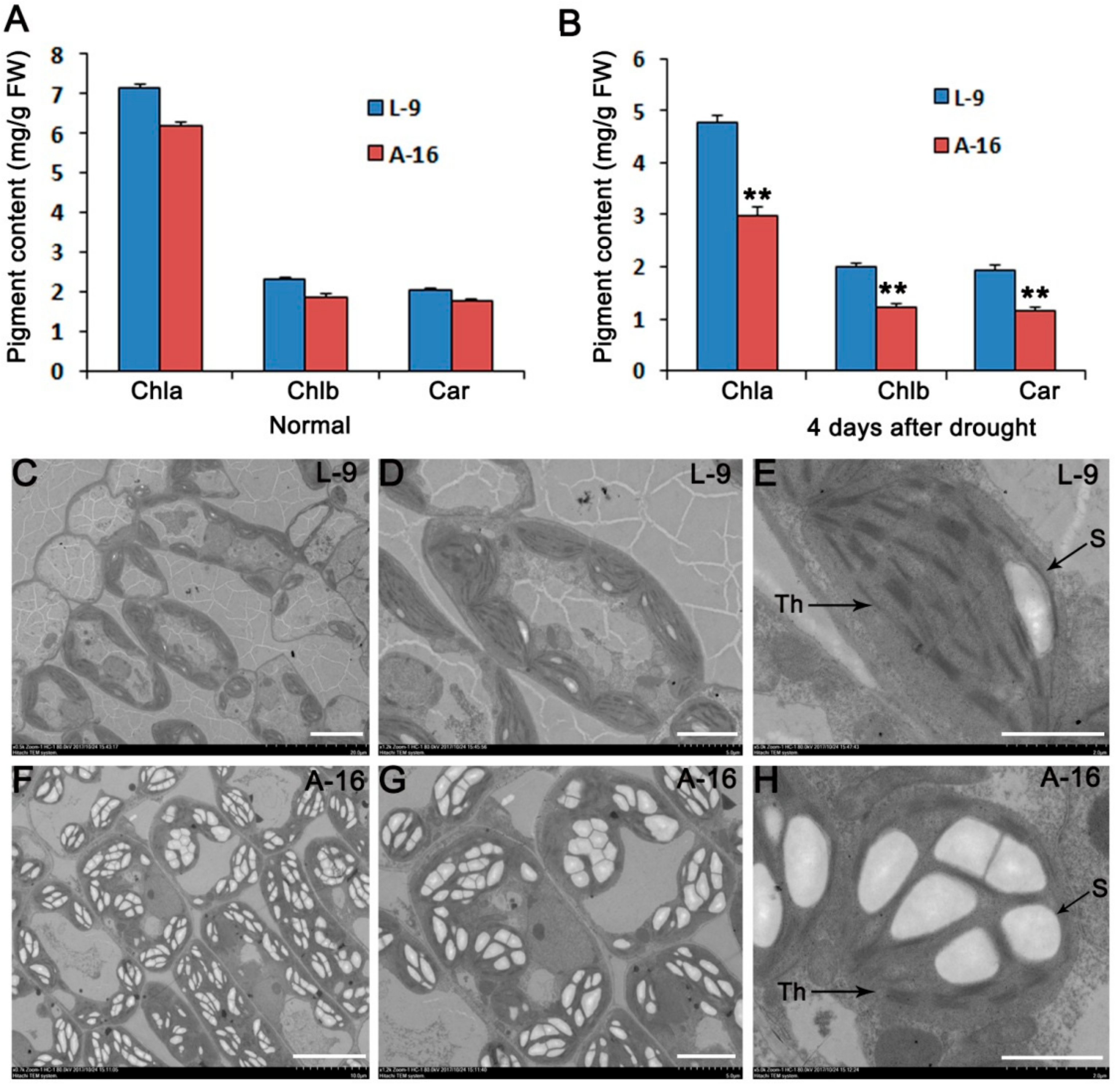
2.1. A-16 Cultivar Is Sensitive to Drought Stress
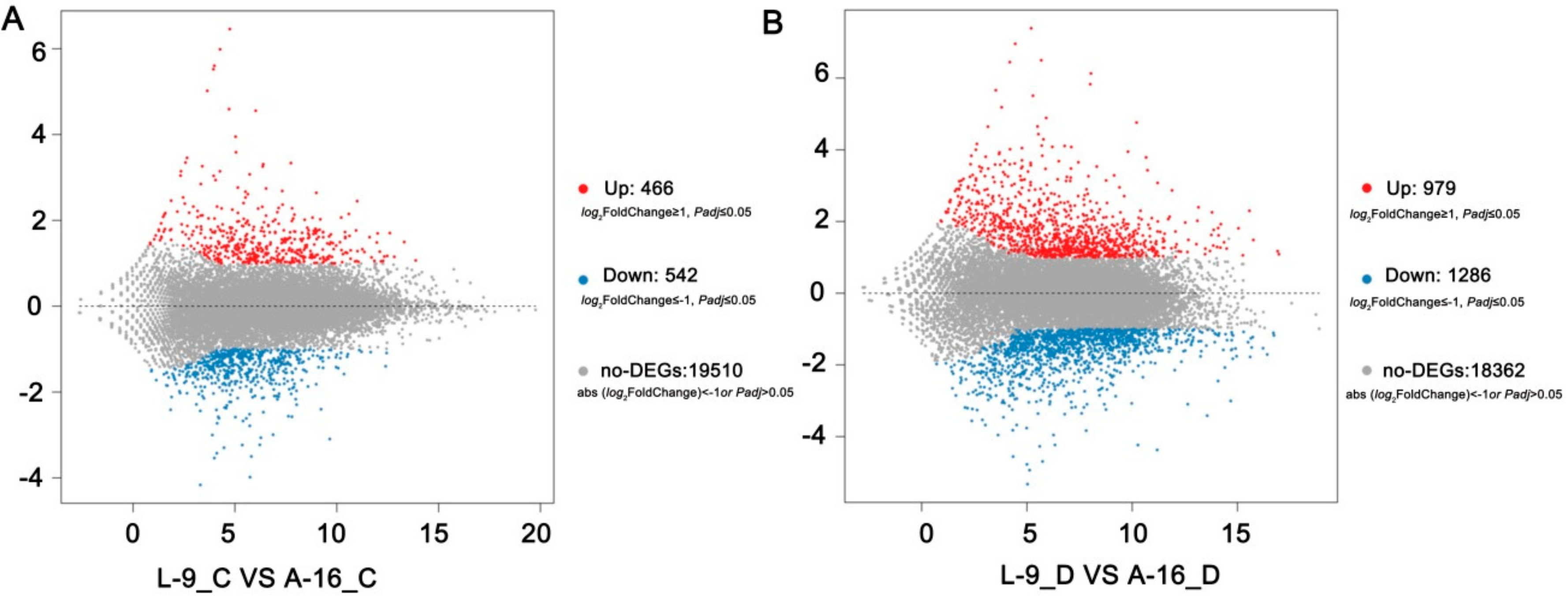
2.2. Drought Stress Results in Extensive Transcriptomic Reprogramming
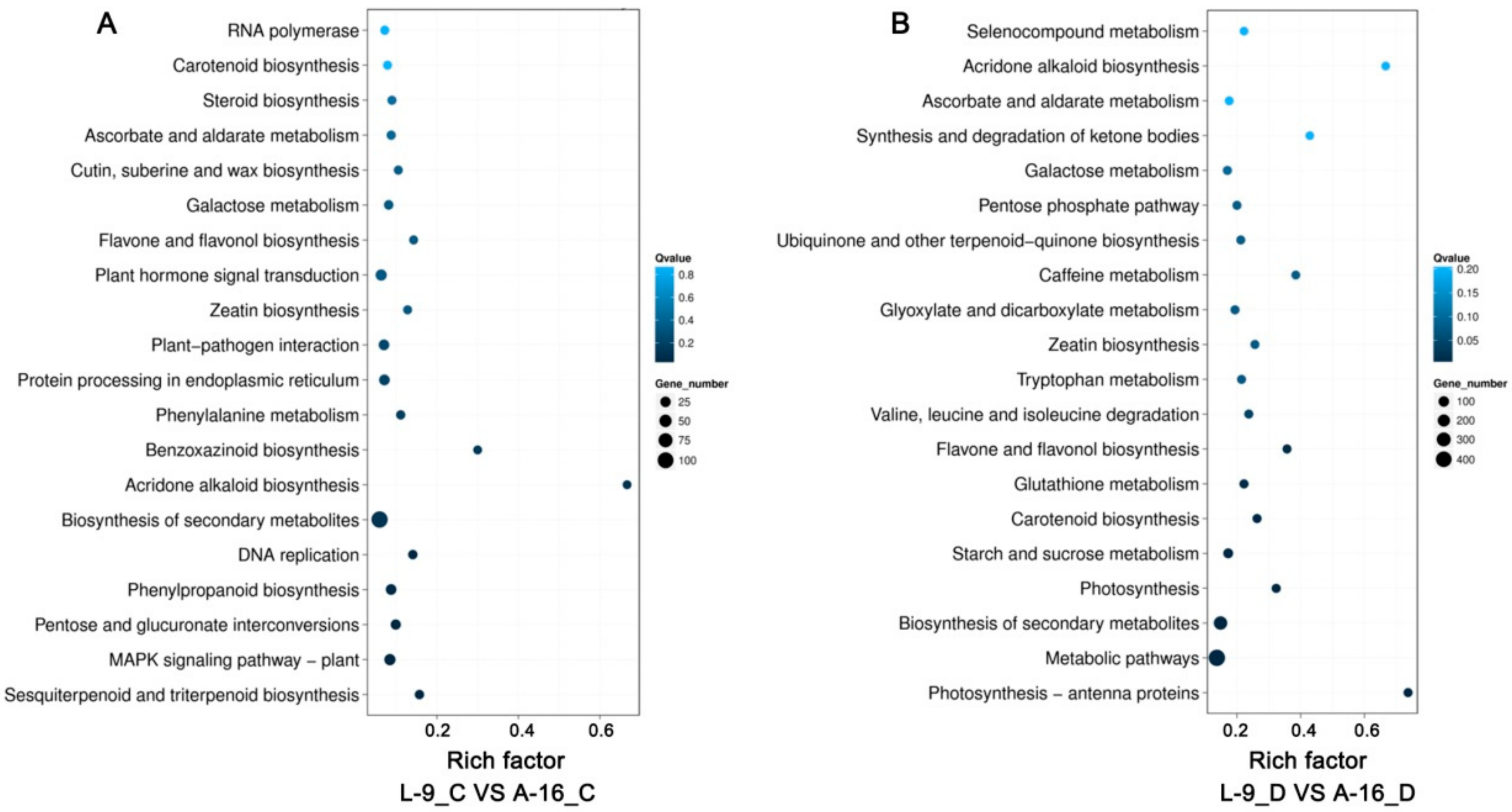
2.3. Functional Classification of Drought-Responsive Genes
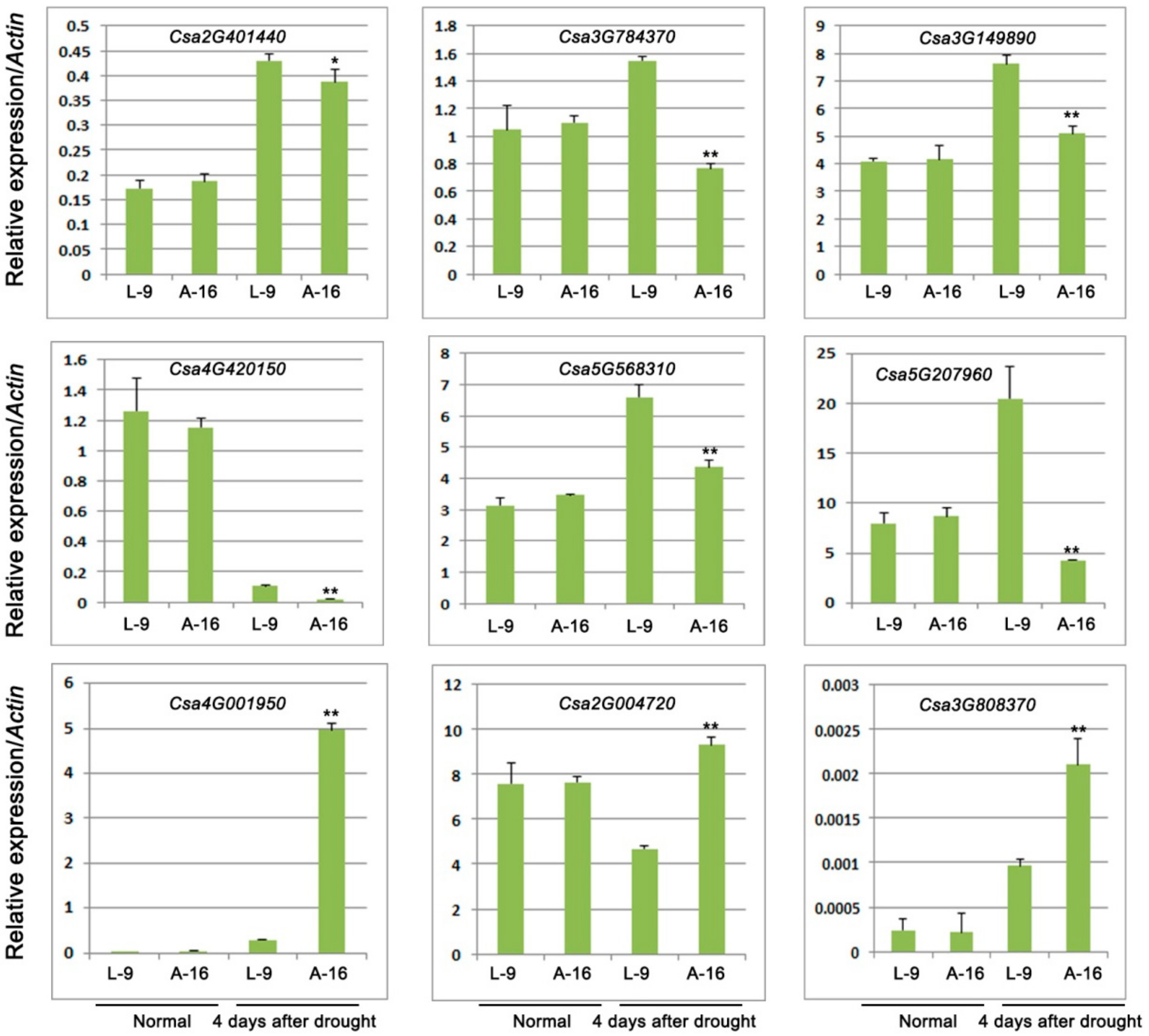
2.4. Expression of Genes Involved in Sucrose Biosynthesis and Response to Water Deprivation
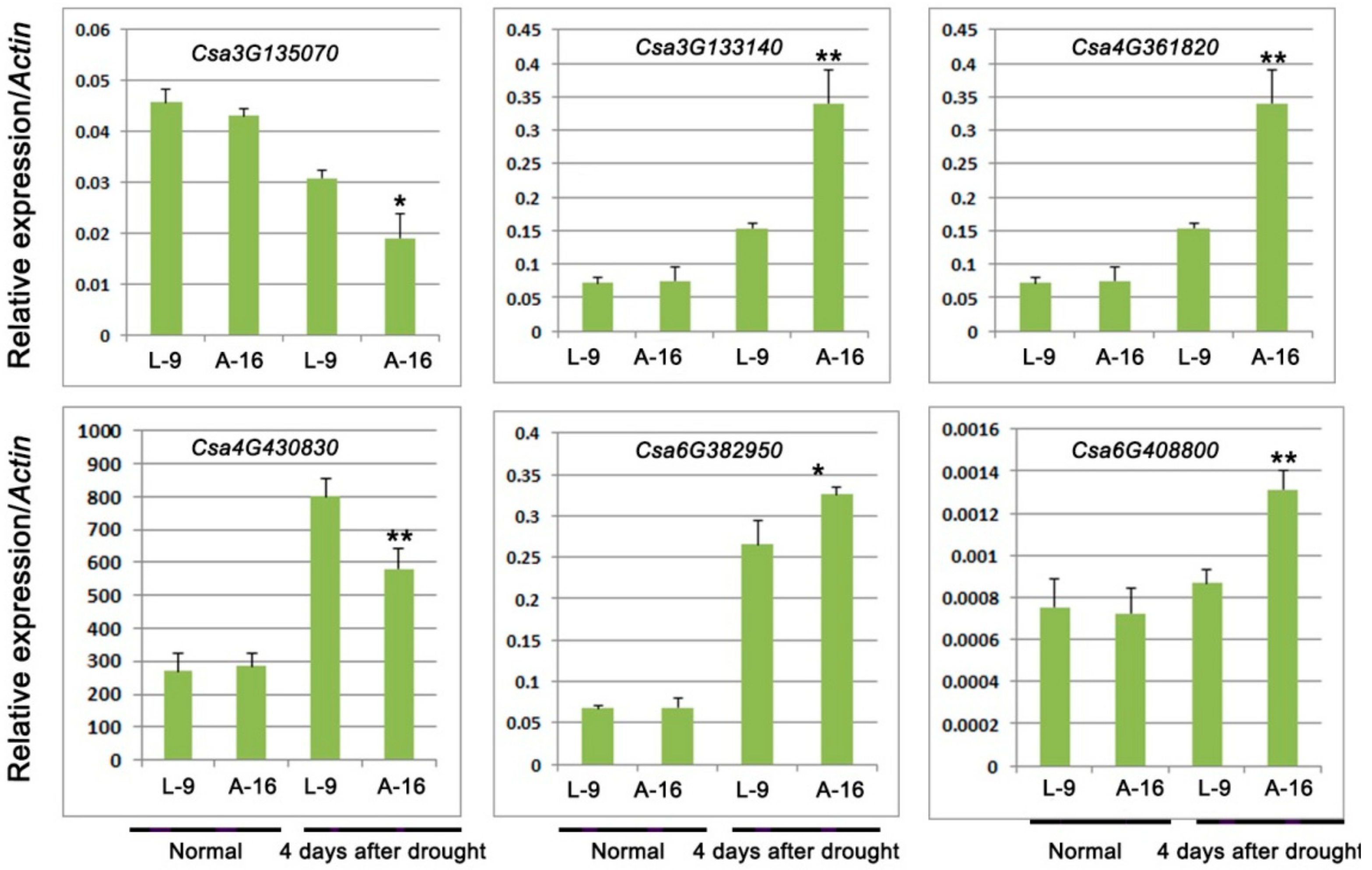
2.5. Analysis of Abscisic Acid (ABA)-Related Genes
3. Discussion
3.1. A-16 Has Less Stomata in the Leaf Than L-9
3.2. Analysis of Sucrose and Starch Biosynthetic Process in Drought Stress
3.3. Analysis of ABA Signal under Drought Stress
3.4. Analysis of Cuticular Waxes Biosynthesis under Drought Stress
4. Materials and Methods
4.1. Plant Materials and Drought Treatment
4.2. Quantitative Analysis of Chlorophyll Content
4.3. Analysis of Malondialdehyde (MDA) Content by TBA Method
4.4. Scanning Electron Microscopy (SEM)
4.5. Transmission Electron Microscopy (TEM)
4.6. BGISEQ-500 Library Construction
4.7. Screening and Significant Test for Differentially Expressed Genes (DEGs)
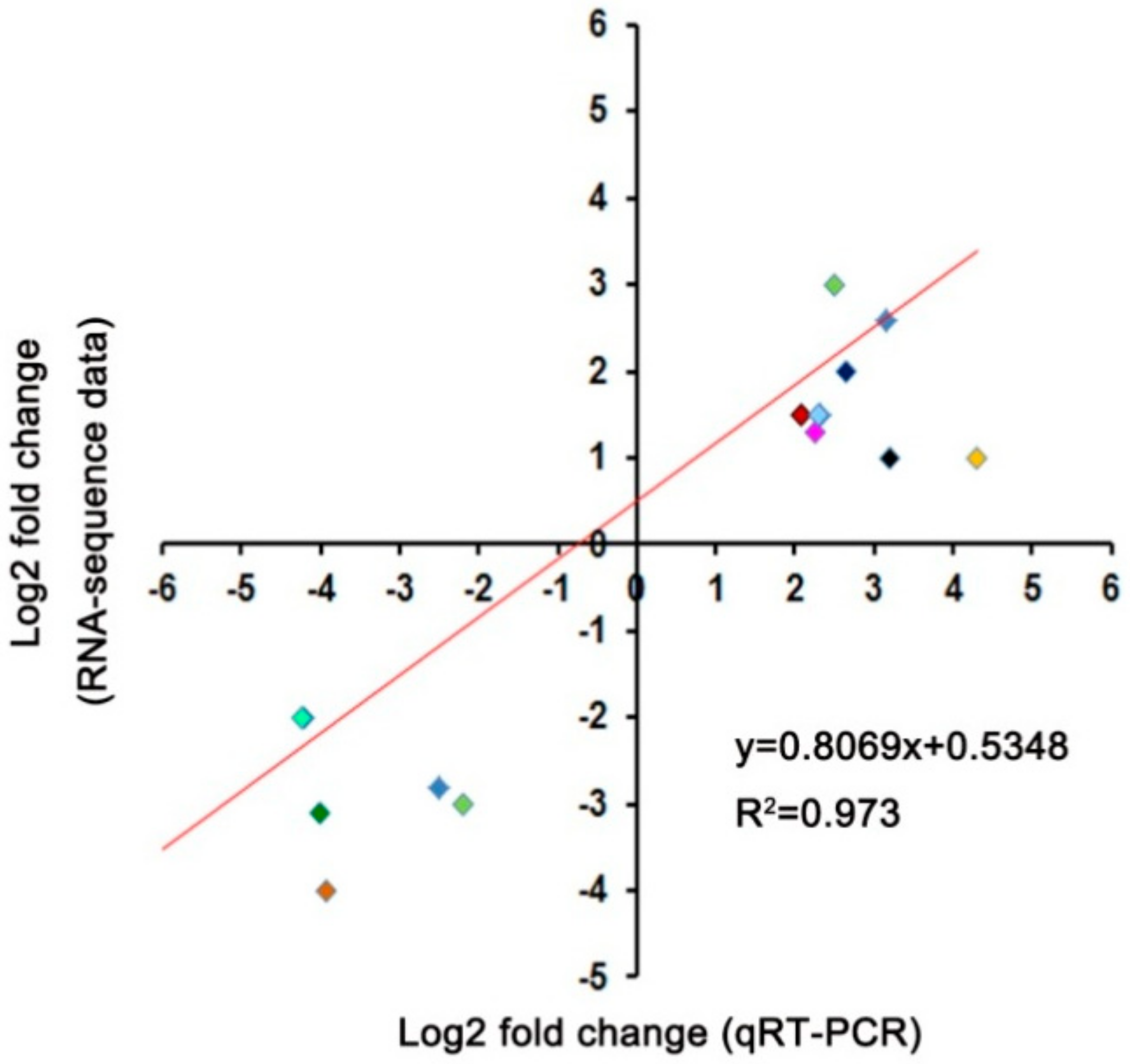
4.8. Quantitative Real-Time PCR (qRT-PCR) Identification
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| RNA seq | RNA sequencing |
| DEGs | Differently expressed genes |
| ABA | Abscisic acid |
| MDA | Malondialdehyde |
| SOD | Enzyme activity of superoxide dismutase |
| SEM | Scanning electron microscopy |
| TEM | Transmission electron microscopy |
| GO | Gene ontology |
| qRT-PCR | Quantitative Real-Time PCR |
References
- Jaleel, C.A.; Manivannan, P.A.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.A.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Kusaka, M.; Ohta, M.; Fujimura, T. Contribution of inorganic components to osmotic adjustment and leaf folding for drought tolerance in pearl millet. Phys. Plant. 2005, 125, 474–489. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Shao, M.A.; Jaleel, C.A.; Hong, M.M. Higher plant antioxidants and redox signaling under environmental stresses. C. R. Biol. 2008, 331, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Bohnert, H.J.; Nelson, D.E.; Jensen, R.G. Adaptations to environmental stresses. Plant Cell 1995, 7, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zhu, J.K. Molecular and genetic aspects of plant responses to osmotic stress. Plant Cell Environ. 2002, 25, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2007, 58, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.M.; Mori, I.C.; Pei, Z.M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jone, J.D.G.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Tsz-fung, F.C.; et al. Abscisic acid inhibits type 2c protein phosphatases via the pyr/pyl family of start proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef] [PubMed]
- Malepszy, S. Cucumber (Cucumis Sativus L.). In Crops II; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1988. [Google Scholar]
- Chen, Y.; Wang, M.; Hu, L.; Liao, W.; Dawuda, M.M.; Li, C. Carbon monoxide is involved in hydrogen gas-induced adventitious root development in cucumber under simulated drought stress. Front. Plant Sci. 2017, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, Y.; Xu, C.; Ren, J.; Liu, X.; Black, K.; Gai, X.; Wang, Q.; Ren, H. Cucumber ECERIFERUM1, (CsCER1), which influences the cuticle properties and drought tolerance of cucumber, plays a key role in VLC alkanes biosynthesis. Plant Mol. Biol. 2015, 87, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, H.; Liu, S.; Peng, X. Exogenous application of hydrogen peroxide alleviates drought stress in cucumber seedlings. S. Afr. J. Bot. 2016, 106, 23–28. [Google Scholar] [CrossRef]
- Yin, Z.; Hennig, J.; Szwacka, M.; Malepszy, S. Tobacco PR-2d promoter is induced in transgenic cucumber in response to biotic and abiotic stimuli. J. Plant Physiol. 2004, 161, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Shankar, R.; Thakkar, B.; Kudapa, H.; Krishnamurthy, L.; Mantri, N.; Varshney, R.K.; Bhatia, S.; Jain, M. Transcriptome analyses reveal genotype-and developmental stage-specific molecular responses to drought and salinity stresses in chickpea. Sci. Rep. 2016, 6, 19228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Yang, P.; Cui, F.; Zhang, F.; Luo, X.; Xie, J. Transcriptome Analysis of Salt Stress Responsiveness in the Seedlings of Dongxiang Wild Rice (Oryza rufipogon Griff.). PLoS ONE 2016, 11, e0146242. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.Y.; Gunewardena, S.S.; Yoo, B.; Liu, J.; Renaud, H.J.; Lu, H.; Zhong, X.B.; Klaassen, C.D. RNA-Seqreveals different mRNA abundance of transporters and their alternative transcript isoforms during liver development. Toxicol. Sci. 2012, 127, 592–608. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Fung-Leung, W.P.; Bittner, A.; Ngo, K.; Liu, X. Comparison of RNA-Seq and microarray in transcriptomeprofiling of activated T cells. PLoS ONE 2014, 9, e78644. [Google Scholar]
- Ando, K.; Grumet, R. Transcriptional profiling of rapidly growing cucumber fruit by 454-pyrosequencing analysis. J. Am. Soc. Hortic. Sci. 2010, 135, 291–302. [Google Scholar]
- Li, J.; Wu, Z.; Cui, L.; Zhang, T.; Guo, Q.; Xu, J.; Jia, L.; Lou, Q.; Huang, S.; Li, Z.; et al. Transcriptome comparison of global distinctive features between pollination and parthenocarpic fruit set reveals transcriptional phytohormone cross-talk in cucumber (Cucumis sativus L.). Plant Cell Physiol. 2014, 55, 1325–1342. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Zheng, Y.; Joung, J.G.; Liu, S.; Zhang, Z.; Crasta, O.R.; Sobral, B.W.; Xu, Y.; Huang, S.; Fei, Z. Transcriptome sequencing and comparative analysis of cucumber flowers with different sex types. BMC Genom. 2010, 11, 384. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, M.; Jiang, L.; Liu, X.; Zhao, J.; Yan, S.; Yang, S.; Ren, H.; Liu, R.; Zhang, X. Transcriptome profiling reveals roles of meristem regulators and polarity genes during fruit trichome development in cucumber (Cucumis sativus L.). J. Exp. Bot. 2014, 65, 4943–4958. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, H.J.; Zhao, B.; Sun, Q.Q.; Cao, Y.Y.; Li, R.; Wu, X.X.; Weeda, S.; Li, L.; Ren, S.; et al. The RNA-seq approach to discriminate gene expression profiles in response to melatonin on cucumber lateral root formation. J. Pineal Res. 2014, 56, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Yang, X.; Yu, H.; Jiang, W.; Sun, N.; Liu, X.; Liu, X.; Zhang, X.; Wang, Y.; Gu, X. RNA-Seq-based transcriptome profiling of early nitrogen deficiency response in cucumber seedlings provides new insight into the putative nitrogen regulatory network. Plant Cell Physiol. 2015, 56, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.W.; Baek, W.; Jung, J.; Kim, J.H.; Lee, S.C. Function of ABA in Stomatal Defense against Biotic and Drought Stresses. Int. J. Mol. Sci. 2015, 16, 15251–15270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, P.; Tan, J.; Jin, M.; Wu, F.; Zhou, K.; Ma, W.; Heng, Y.; Wang, J.; Guo, X.; Zhang, X.; et al. Albino midrib 1, encoding a putative potassium efflux antiporter, affects chloroplast development and drought tolerance in rice. Plant Cell Rep. 2014, 33, 1581–1594. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.G.; Shan, J.X.; Shi, M.; Gao, J.P.; Lin, H.X. DCA1 acts as a transcriptional co-activator of DST and contributes to drought and salt tolerance in rice. PLoS Genet. 2015, 11, e1005617. [Google Scholar] [CrossRef] [PubMed]
- Mohammadkhani, N.; Heidari, R. Drought-induced Accumulation of Soluble Sugars and Proline in Two Maize Varieties. World Appl. Sci. J. 2008, 3, 448–453. [Google Scholar]
- Al Hakimi, A.; Monneveux, P.; Galiba, G. Soluble sugars, proline and relative water content (RCW) as traits for improving drought tolerance and divergent selection for RCW from triticumpolonicum into triticum durum. J. Genet. Breed. 1995, 49, 237–244. [Google Scholar]
- Prado, F.E.; Boero, C.; Gallardo, M.; González, J.A. Effect of nacl on germination, growth, and soluble sugar content in chenopodium quinoa willd. seeds. Bot. Bull. Acad. Sin. 2000, 41, 27–34. [Google Scholar]
- Sperdouli, I.; Moustakas, M. Interaction of proline, sugars, and anthocyanins during photosynthetic acclimation of Arabidopsis thaliana to drought stress. J. Plant Physiol. 2012, 169, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Danquah, A.; de Zelicourt, A.; Colcombet, J.; Hirt, H. The role of ABA and MAPK signaling pathways in plant abiotic stress responses. Biotechnol. Adv. 2014, 32, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Chen, B.; Lu, G.; Han, B. Overexpression of a NAC transcription factor enhances rice drought and salt tolerance. Biochem. Biophys. Res. Commun. 2009, 379, 985–989. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Li, X.; Jin, S.; Liu, X.; Zhu, L.; Nie, Y.; Zhang, X. Overexpression of rice NAC gene SNAC1 improves drought and salt tolerance by enhancing root development and reducing transpiration rate in transgenic cotton. PLoS ONE 2014, 9, e86895. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Xiong, A.; Peng, R.; Jin, X.; Xu, J.; Zhu, B.; Chen, J.; Yao, Q. OsNAC52, a rice NAC transcription factor, potentially responds to ABA and confers drought tolerance in transgenic plants. Plant Cell Tissue Organ Cult. 2010, 100, 255–262. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, H. Roles of calcium-dependent protein kinases in ABA-regulation of stomatal moment in Poplar. Sci. Technol. Eng. 2004, 4, 80–83. (In Chinese) [Google Scholar]
- Zhu, S.Y.; Yu, X.C.; Wang, X.J.; Zhao, R.; Li, Y.; Fan, R.C.; Shang, Y.; Du, S.Y.; Wang, X.F.; Wu, F.Q.; et al. Two Calcium-Dependent Protein Kinases, CPK4 and CPK11, Regulate Abscisic Acid Signal Transduction in Arabidopsis. Plant Cell 2007, 19, 3019–3036. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.J.; Wei, F.J.; Wang, C.; Wu, J.J.; Ratnasekera, D.; Liu, W.X.; Wu, W.H. Arabidopsis calcium-dependent protein kinase CPK10 functions in abscisic acid- and Ca2+-mediated stomatal regulation in response to drought stress. Plant Physiol. 2010, 154, 1232–1243. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.T.; Song, W. ZmCK3, a maize calcium-dependent protein kinase gene, endows tolerance to drought and drought stresses in transgenic Arabidopsis. J. Plant Biochem. Biotechnol. 2014, 23, 249–256. [Google Scholar] [CrossRef]
- Liu, G.; Chen, J.; Wang, X. VfCPK1, a gene encoding calcium-dependent protein kinase from Vicia faba, is induced by drought and abscisic acid. Plant Cell Environ. 2006, 29, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Peng, L.; Liu, Y.; Yao, R.; Liu, Z.; Li, X.; Yang, Y.; Wang, J. Arabidopsis calcium-dependent protein kinase AtCPK1 plays a positive role in salt/drought-stress response. Biochem. Biophys. Res. Commun. 2017, 49. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Joubès, J. Arabidopsis cuticular waxes: Advances in synthesis, export and regulation. Prog. Lipid Res. 2013, 52, 110–129. [Google Scholar] [CrossRef] [PubMed]
- España, L.; Heredia-Guerrero, J.A.; Reina-Pinto, J.J.; Fernández-Muñoz, R.; Heredia, A.; Domínguez, E. Transient silencing of CHALCONE SYNTHASE during fruit ripening modifies tomato epidermal cells and cuticle properties. Plant Physiol. 2014, 166, 1371–1386. [Google Scholar] [CrossRef] [PubMed]
- Reisige, K.; Gorzelanny, C.; Daniels, U.; Moerschbacher, B.M. The C28 aldehyde octacosanal is a morphogenetically active component involved in host plant recognition and infection structure differentiation in the wheat stem rust fungus. Physiol. Mol. Plant Pathol. 2006, 68, 33–40. [Google Scholar] [CrossRef]
- Shepherd, T.; Wynne, G.D. The effects of stress on plant cuticular waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.; Makino, A. Availability of rubisco small subunit up-regulates the transcript levels of large subunit for stoichiometric assembly of its holoenzyme in rice. Plant Physiol. 2012, 160, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Fehlmann, T.; Reinheimer, S.; Geng, C.; Su, X.; Drmanac, S.; Alexeev, A.; Zhang, C.; Backes, C.; Ludwig, N.; Hart, M.; et al. cPAS-based sequencing on the BGISEQ-500 to explore small non-coding RNAs. Clin. Epigenet. 2016, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L.; et al. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, W293. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accuratenormalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal controlgenes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Clean Reads | Total Clean Bases (Gb) | Total Mapping Ratio % | Uniquely Mapping Ratio % |
|---|---|---|---|---|
| A-16_C1 | 23,001,330 | 1.15 | 0.9664 | 0.8976 |
| A-16_C2 | 22,799,582 | 1.14 | 0.9662 | 0.8986 |
| A-16_C3 | 23,275,316 | 1.16 | 0.966 | 0.9013 |
| A-16_D1 | 23,202,361 | 1.16 | 0.963 | 0.8943 |
| A-16_D2 | 23,239,914 | 1.16 | 0.9625 | 0.8981 |
| A-16_D3 | 23,127,940 | 1.16 | 0.959 | 0.8924 |
| L-9_C1 | 23,343,741 | 1.17 | 0.966 | 0.8971 |
| L-9_C2 | 23,065,366 | 1.15 | 0.9617 | 0.8901 |
| L-9_C3 | 23,055,733 | 1.15 | 0.9634 | 0.8923 |
| L-9_D1 | 22,973,680 | 1.15 | 0.9639 | 0.8927 |
| L-9_D2 | 23,183,117 | 1.16 | 0.963 | 0.897 |
| L-9_D3 | 23,037,373 | 1.15 | 0.956 | 0.8932 |
| Gene ID | L-9 Expression | A-16 Expression | Regulation | p-Value | Annotation |
|---|---|---|---|---|---|
| Csa2G401440 | 2237.5 | 1091.6 | Down | 0.00434039 | Sucrose-phosphate synthase |
| Csa3G784370 | 3412.9 | 1102.8 | Down | 1.20 × 10−6 | Sucrose phosphatase |
| Csa3G149890 | 10,202.0 | 3362.5 | Down | 3.87 × 10−12 | Glucose-1-phosphate adenylyltransferase |
| Csa4G001950 | 2345.5 | 7726.7 | Up | 6.84 × 10−6 | Sucrose synthase |
| Csa4G420150 | 492.7 | 239.1 | Down | 1.17 × 10−5 | 4-α-Glucanotransferase |
| Csa5G568310 | 4872.5 | 2423.0 | Down | 3.78 × 10−6 | Phosphoglucomutase |
| Csa2G004720 | 1255.1 | 2945.9 | Up | 1.42 × 10−6 | Multiprotein-bridging factor |
| Csa5G207960 | 11,815.2 | 4338.2 | Down | 1.77 × 10−9 | Omega-3 fatty acid desaturase |
| Csa3G808370 | 47.5 | 102.9 | Up | 0.00031088 | Seed maturation protein LEA 4 |
| Gene ID | L-9 Expression | A-16 Expression | Regulation | p-Value | Annotation |
|---|---|---|---|---|---|
| Csa3G135070 | 89.6 | 20.4 | Down | 3.05 × 10−9 | Calcium-dependent protein kinase |
| Csa3G133140 | 463.1 | 1758.6 | Up | 1.72 × 10−7 | 3-Ketoacyl-CoA thiolase 1 |
| Csa4G361820 | 1298.2 | 4273.3 | Up | 1.04 × 10−17 | NAC domain-containing protein |
| Csa4G430830 | 276.8 | 50.0 | Down | 1.29 × 10−18 | Calcium-dependent protein kinase-like protein |
| Csa6G382950 | 125.3 | 405.7 | Up | 4.08 × 10−8 | NAC domain-containing protein |
| Csa6G408800 | 47.5 | 102.9 | Up | 0.0003109 | Circadian clock coupling factor, |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Jiang, B.; Peng, Q.; Liu, W.; He, X.; Liang, Z.; Lin, Y. Transcriptome Analyses in Different Cucumber Cultivars Provide Novel Insights into Drought Stress Responses. Int. J. Mol. Sci. 2018, 19, 2067. https://doi.org/10.3390/ijms19072067
Wang M, Jiang B, Peng Q, Liu W, He X, Liang Z, Lin Y. Transcriptome Analyses in Different Cucumber Cultivars Provide Novel Insights into Drought Stress Responses. International Journal of Molecular Sciences. 2018; 19(7):2067. https://doi.org/10.3390/ijms19072067
Chicago/Turabian StyleWang, Min, Biao Jiang, Qingwu Peng, Wenrui Liu, Xiaoming He, Zhaojun Liang, and Yu’e Lin. 2018. "Transcriptome Analyses in Different Cucumber Cultivars Provide Novel Insights into Drought Stress Responses" International Journal of Molecular Sciences 19, no. 7: 2067. https://doi.org/10.3390/ijms19072067
APA StyleWang, M., Jiang, B., Peng, Q., Liu, W., He, X., Liang, Z., & Lin, Y. (2018). Transcriptome Analyses in Different Cucumber Cultivars Provide Novel Insights into Drought Stress Responses. International Journal of Molecular Sciences, 19(7), 2067. https://doi.org/10.3390/ijms19072067