Selumetinib Activity in Thyroid Cancer Cells: Modulation of Sodium Iodide Symporter and Associated miRNAs


Abstract
:1. Introduction
2. Results
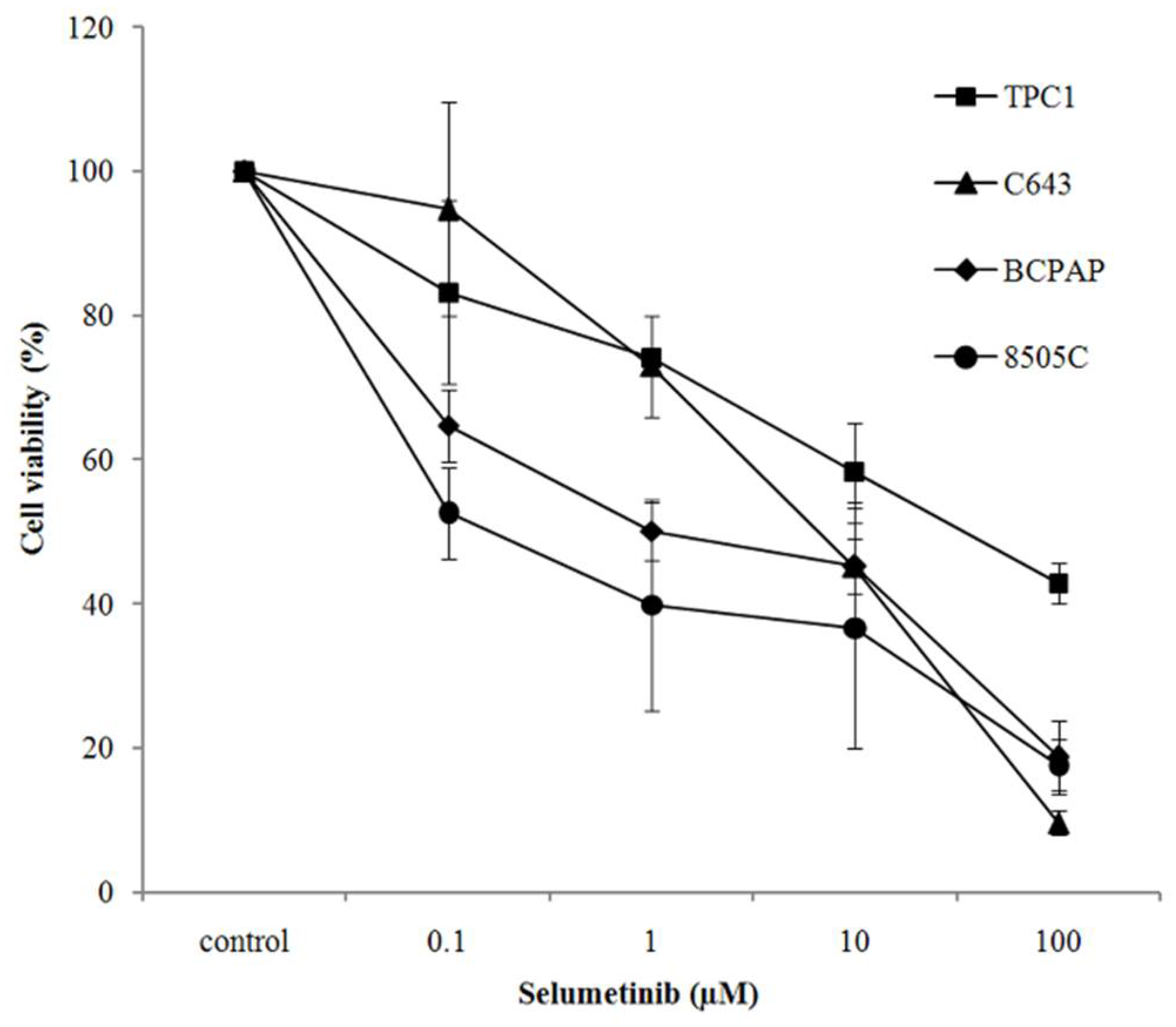
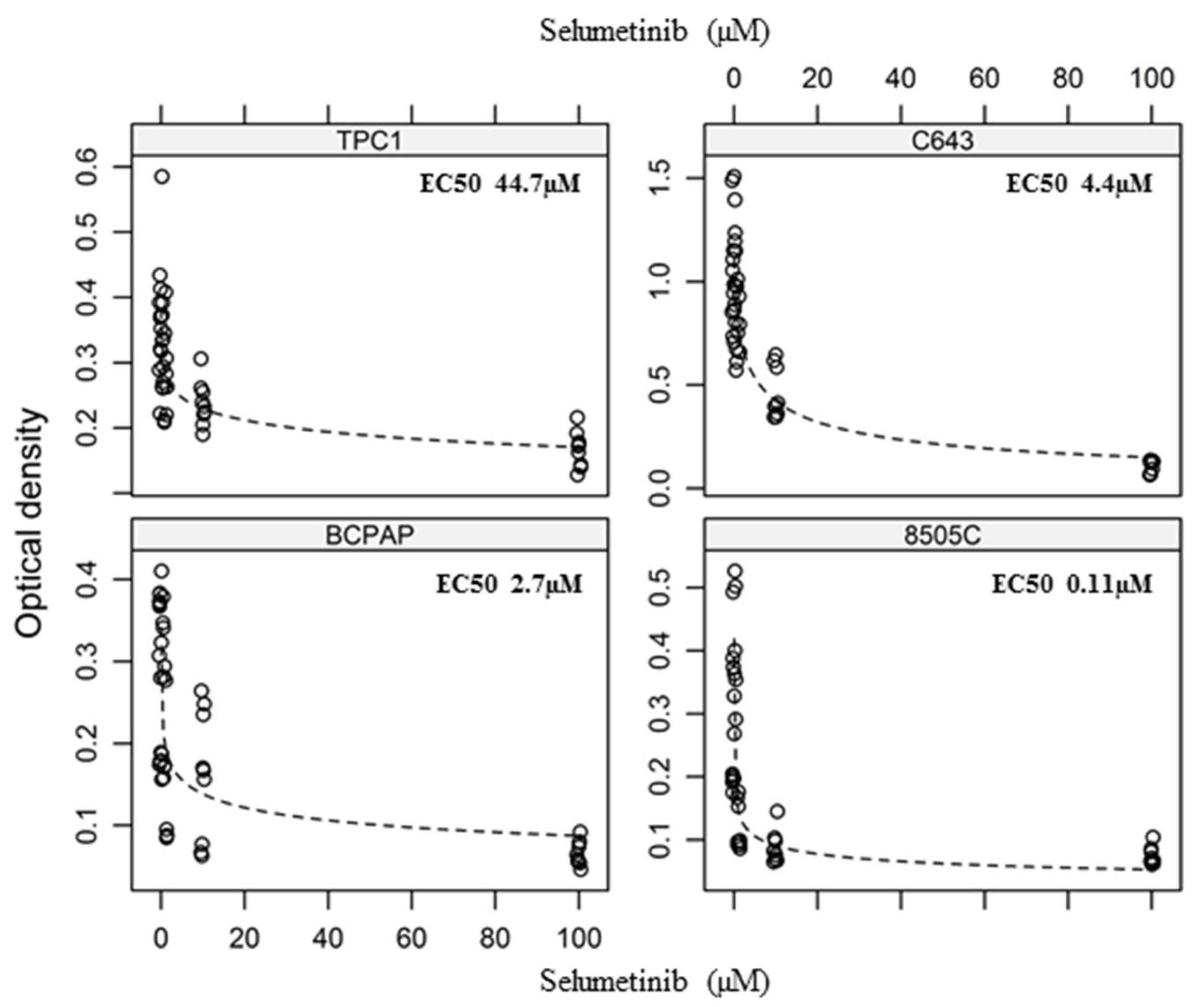
2.1. Selumetinib Cytotoxic Effects
2.2. NIS Restoration after Selumetinib Treatment
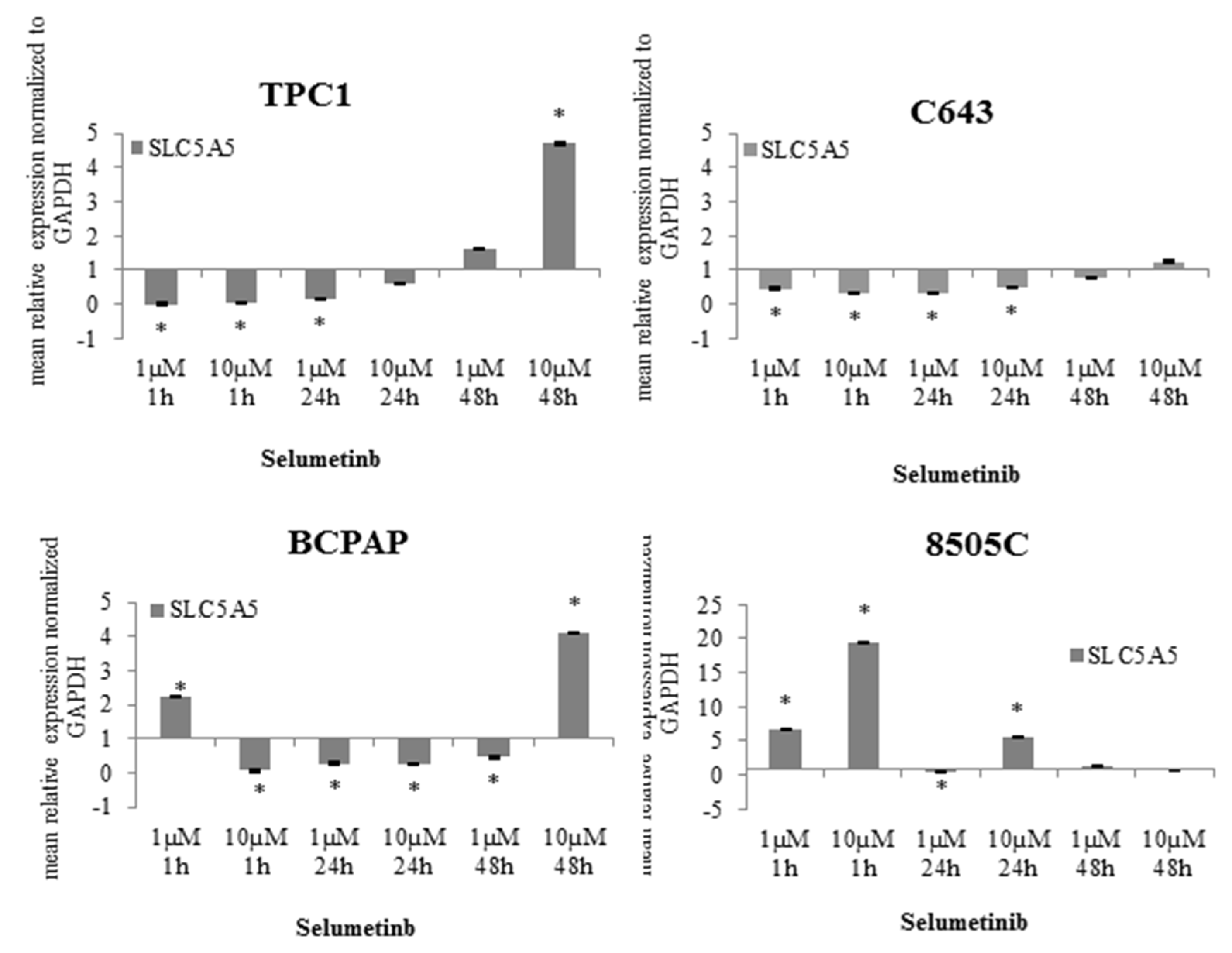
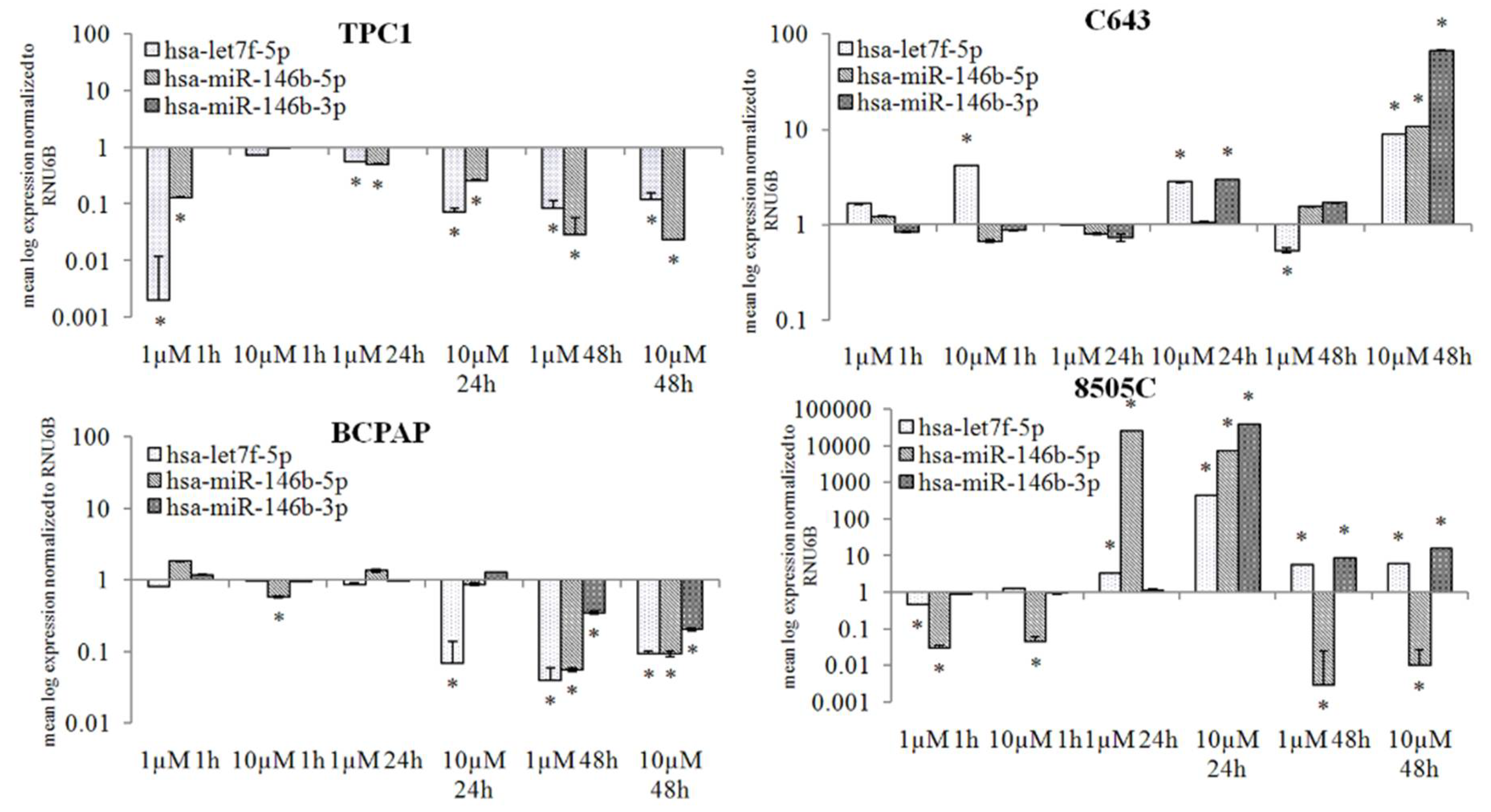
2.3. Selumetinib Impaired Expression of NIS Related miRNAs
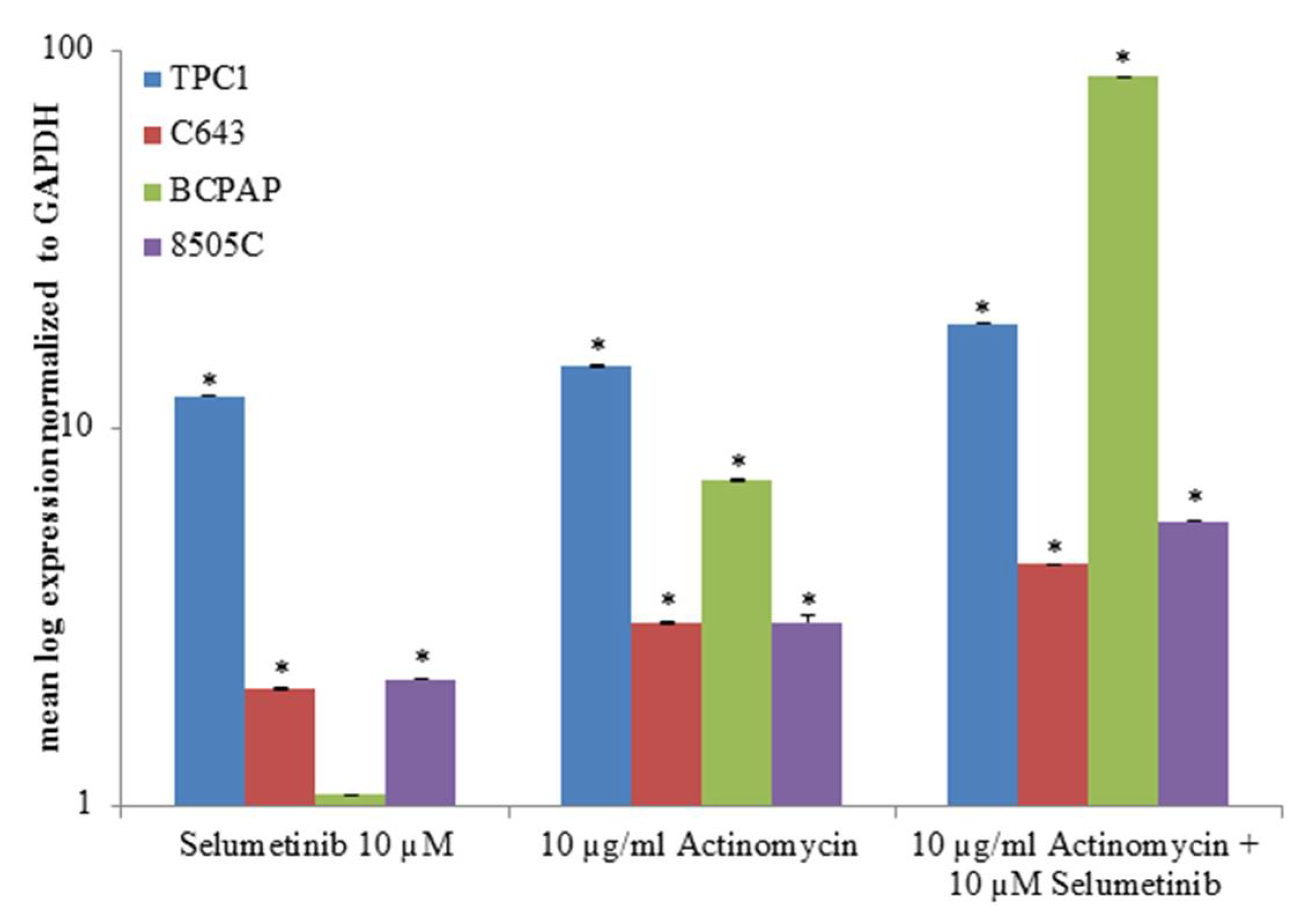
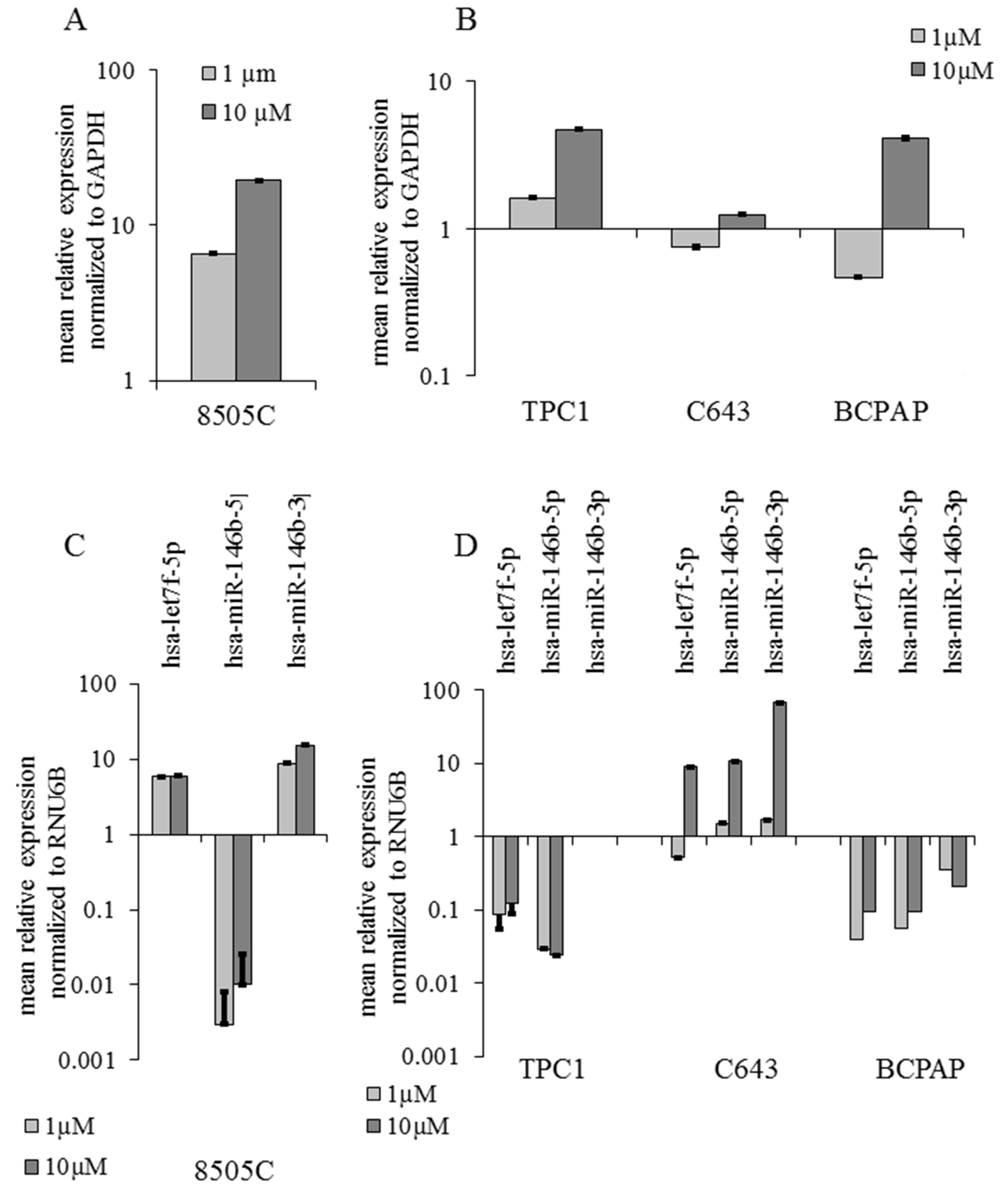
2.4. miRNAs Expression Correlation with the Validated Target NIS after Selumetinib Administration
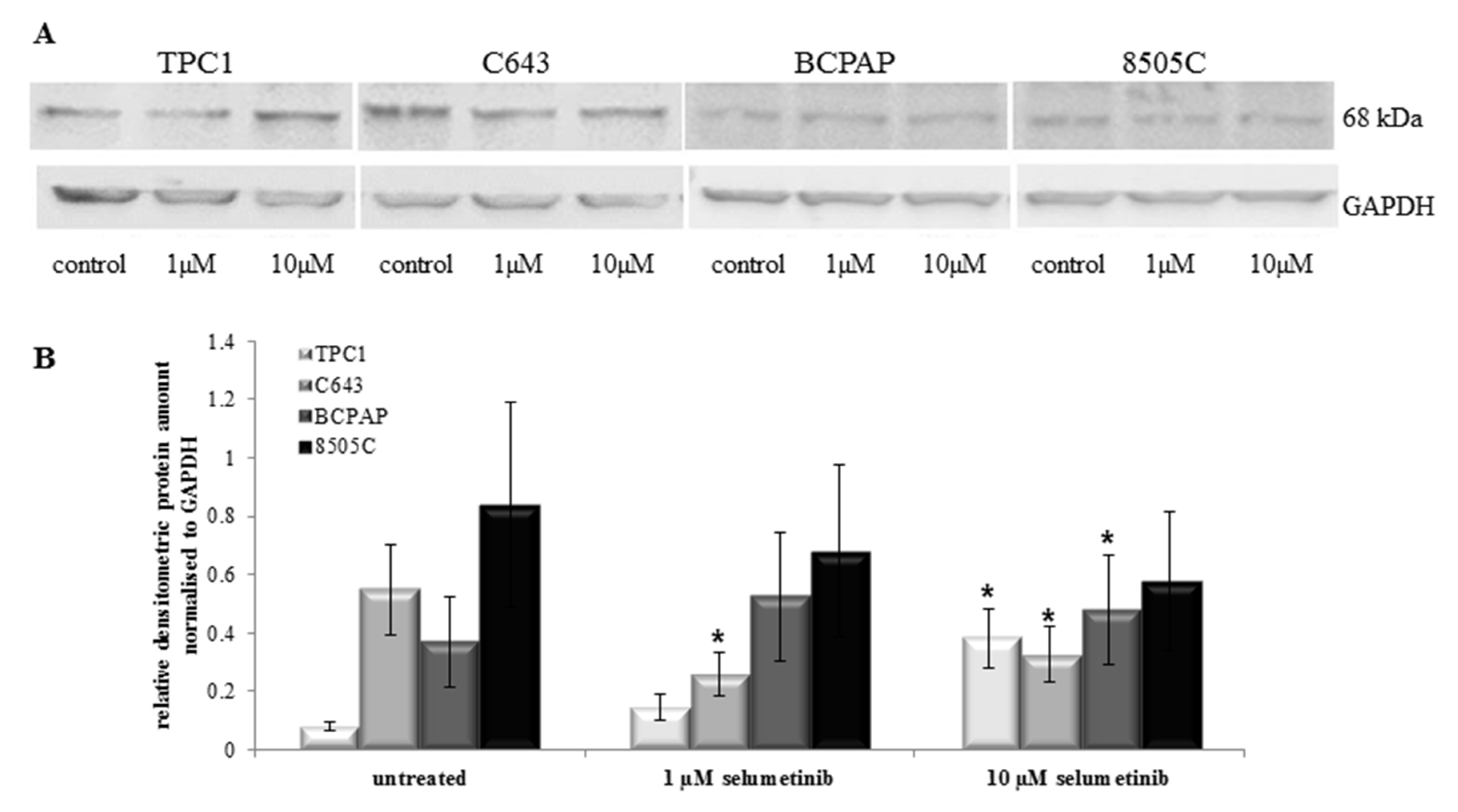
2.5. Selumetinib Treatment Did Not Always Change NIS Protein Expression
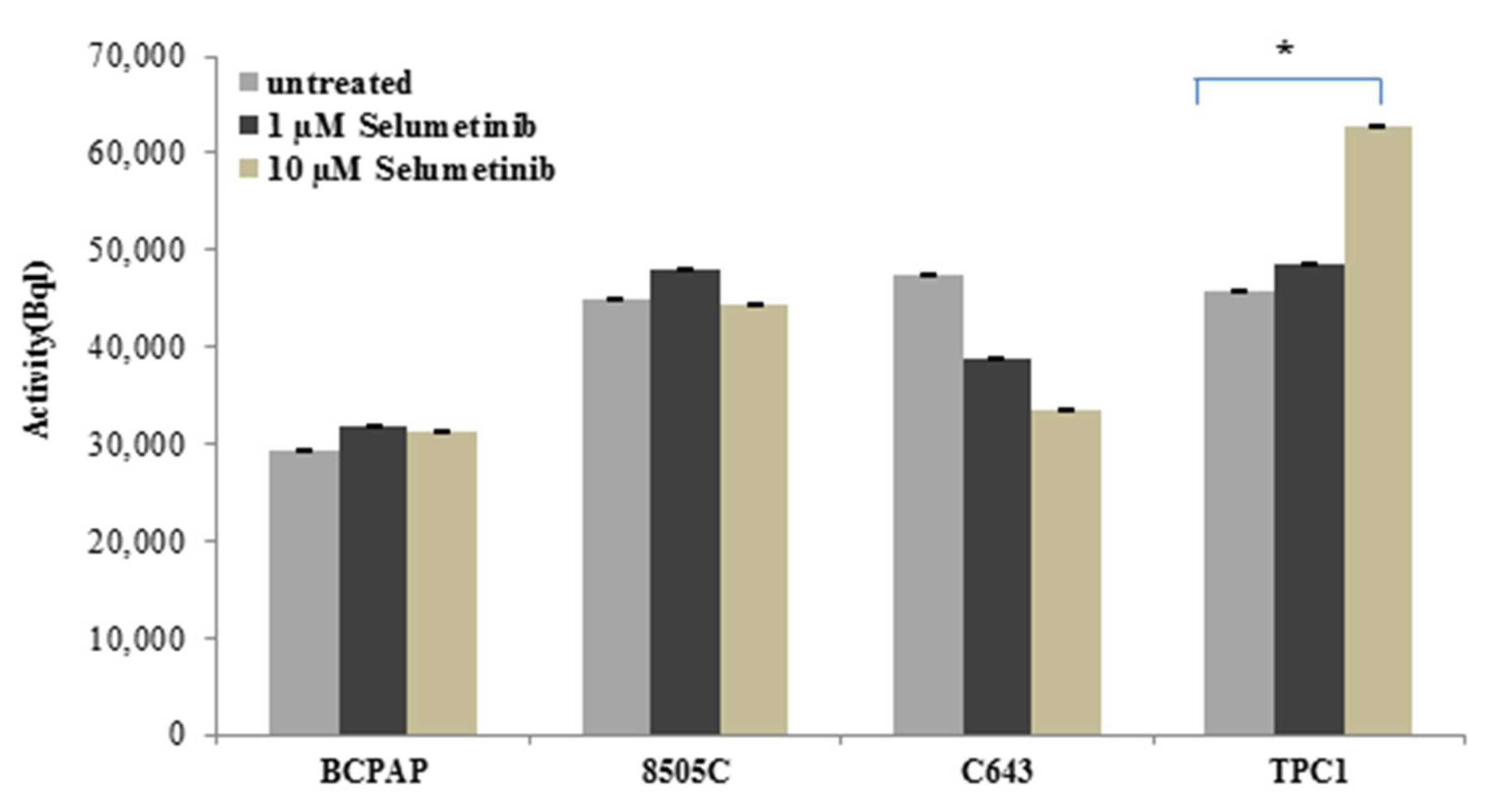
2.6. Sodium Iodide Symporter Activity after Selumetinib Treatment
3. Discussion
4. Material and Methods
4.1. Cell Lines
4.2. Cell Culture
4.3. Drug Preparation
4.4. Analysis of Cell Viability
4.5. RNA Isolation and Quantitative Real Time RT-PCR
4.6. Protein Isolation and Western Blotting
4.7. In Vitro Radioiodine Uptake
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Busaidy, N.L.; Cabanillas, M.E. Differentiated thyroid cancer: Management of patients with radioiodine nonresponsive disease. J. Thyroid Res. 2012, 2012, 618985. [Google Scholar] [CrossRef] [PubMed]
- Kiyota, N.; Schlumberger, M.; Muro, K.; Ando, Y.; Takahashi, S.; Kawai, Y.; Wirth, L.; Robinson, B.; Sherman, S.; Suzuki, T.; et al. Subgroup analysis of Japanese patients in a phase 3 study of lenvatinib in radioiodine-refractory differentiated thyroid cancer. Cancer Sci. 2015, 106, 1714–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durante, C.; Haddy, N.; Baudin, E.; Leboulleux, S.; Hartl, D.; Travagli, J.P.; Caillou, B.; Ricard, M.; Lumbroso, J.D.; De Vathaire, F.; et al. Long-term outcome of 444 patients with distant metastases from papillary and follicular thyroid carcinoma: Benefits and limits of radioiodine therapy. J. Clin. Endocrinol. Metab. 2006, 91, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Gilliland, F.D.; Hunt, W.C.; Morris, D.M.; Key, C.R. Prognostic factors for thyroid carcinoma. A population-based study of 15,698 cases from the Surveillance, Epidemiology and End Results (SEER) program 1973–1991. Cancer 1997, 79, 564–573. [Google Scholar] [CrossRef]
- Smallridge, R.C.; Ain, K.B.; Asa, S.L.; Bible, K.C.; Brierley, J.D.; Burman, K.D.; Kebebew, E.; Lee, N.Y.; Nikiforov, Y.E.; Rosenthal, M.S.; et al. American Thyroid Association guidelines for management of patients with anaplastic thyroid cancer. Thyroid 2012, 22, 1104–1139. [Google Scholar] [CrossRef] [PubMed]
- Kebebew, E.; Greenspan, F.S.; Clark, O.H.; Woeber, K.A.; McMillan, A. Anaplastic thyroid carcinoma. Treatment outcome and prognostic factors. Cancer 2005, 103, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.S.; Gharib, H. Epidemiology of thyroid nodules. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Haugen, B.R.; Sherman, S.I. Evolving approaches to patients with advanced differentiated thyroid cancer. Endocr. Rev. 2013, 34, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, H.; Zhao, J.; Yao, J.; Shang, H.; Zhu, H.; Liao, L.; Dong, J. Association between sodium iodide symporter and differentiated Thyroid cancer: A meta-analysis of 9 studies. Int. J. Clin. Exp. Med. 2015, 8, 17986–17994. [Google Scholar] [PubMed]
- Wapnir, I.L.; van de Rijn, M.; Nowels, K.; Amenta, P.S.; Walton, K.; Montgomery, K.; Greco, R.S.; Dohán, O.; Carrasco, N. Immunohistochemical profile of the sodium/iodide symporter in thyroid, breast, and other carcinomas using high density tissue microarrays and conventional sections. J. Clin. Endocrinol. Metab. 2003, 88, 1880–1888. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.L.; Grewal, R.K.; Leboeuf, R.; Sherman, E.J.; Pfister, D.G.; Deandreis, D.; Pentlow, K.S.; Zanzonico, P.B.; Haque, S.; Gavane, S.; et al. Selumetinib-enhanced radioiodine uptake in advanced thyroid cancer. N. Engl. J. Med. 2013, 368, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Croce, C.M. Oncogenes and cancer. N. Engl. J. Med. 2008, 358, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, R.; Ul Hussain, M. MicroRNAs and human diseases: Diagnostic and therapeutic potential. Cell Tissue Res. 2014, 358, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lv, B.; Chen, B.; Guan, M.; Sun, Y.; Li, H.; Zhang, B.; Ding, C.; He, S.; Zeng, Q. Inhibition of miR-146b expression increases radioiodine-sensitivity in poorly differential thyroid carcinoma via positively regulating NIS expression. Biochem. Biophys. Res. Commun. 2015, 462, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Geraldo, M.V.; Fuziwara, C.S.; Friguglieti, C.U.; Costa, R.B.; Kulcsar, M.A.; Yamashita, A.S.; Kimura, E.T. MicroRNAs miR-146-5p and let-7f as prognostic tools for aggressive papillary thyroid carcinoma: A case report. Arq. Bras. Endocrinol. Metabol. 2012, 56, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.K.; Yang, K.D.; Chou, F.F.; Huang, C.C.; Lan, Y.W.; Lee, Y.F.; Kang, H.Y.; Liu, R.T. Prognostic implications of miR-146b expression and its functional role in papillary thyroid carcinoma. J. Clin. Endocrinol. Metab. 2013, 98, E196–E205. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Wu, B.; Xiao, K.; Kang, J.; Xie, J.; Zhang, X.; Fan, Y. MiR-146b-5p promotes metastasis and induces epithelial-mesenchymal transition in thyroid cancer by targeting ZNRF3. Cell Physiol. Biochem. 2015, 35, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, A.; Wojcicka, A.; Kotlarek, M.; Zhang, X.; Jazdzewski, K.; Jhiang, S.M. microRNA-339-5p modulates Na+/I− symporter-mediated radioiodide uptake. Endocr. Relat. Cancer 2015, 22, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Damanakis, A.I.; Eckhardt, S.; Wunderlich, A.; Roth, S.; Wissniowski, T.T.; Bartsch, D.K.; Di Fazio, P. MicroRNAs let7 expression in thyroid cancer: Correlation with their deputed targets HMGA2 and SLC5A5. J. Cancer Res. Clin. Oncol. 2016, 142, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Fuziwara, C.S.; Kimura, E.T. MicroRNA Deregulation in Anaplastic Thyroid Cancer Biology. Int. J. Endocrinol. 2014, 2014, 743450. [Google Scholar] [CrossRef] [PubMed]
- Pallante, P.; Visone, R.; Ferracin, M.; Ferraro, A.; Berlingieri, M.T.; Troncone, G.; Chiappetta, G.; Liu, C.G.; Santoro, M.; Negrini, M.; et al. MicroRNA deregulation in human thyroid papillary carcinomas. Endocr. Relat. Cancer 2006, 13, 497–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swierniak, M.; Wojcicka, A.; Czetwertynska, M.; Stachlewska, E.; Maciag, M.; Wiechno, W.; Gornicka, B.; Bogdanska, M.; Koperski, L.; de la Chapelle, A.; et al. In-depth characterization of the microRNA transcriptome in normal thyroid and papillary thyroid carcinoma. J. Clin. Endocrinol. Metab. 2013, 98, E1401–E1409. [Google Scholar] [CrossRef] [PubMed]
- Colamaio, M.; Calì, G.; Sarnataro, D.; Borbone, E.; Pallante, P.; Decaussin-Petrucci, M.; Nitsch, L.; Croce, C.M.; Battista, S.; Fusco, A. Let-7a down-regulation plays a role in thyroid neoplasias of follicular histotype affecting cell adhesion and migration through its ability to target the FXYD5 (Dysadherin) gene. J. Clin. Endocrinol. Metab. 2012, 97, E2168–E2178. [Google Scholar] [CrossRef] [PubMed]
- Ricarte-Filho, J.C.; Fuziwara, C.S.; Yamashita, A.S.; Rezende, E.; da-Silva, M.J.; Kimura, E.T. Effects of let-7 microRNA on Cell Growth and Differentiation of Papillary Thyroid Cancer. Transl. Oncol. 2009, 2, 236–241. [Google Scholar] [PubMed]
- Spitzweg, C.; Bible, K.C.; Hofbauer, L.C.; Morris, J.C. Advanced radioiodine-refractory differentiated thyroid cancer: The sodium iodide symporter and other emerging therapeutic targets. Lancet Diabetes Endocrinol. 2014, 2, 830–842. [Google Scholar] [CrossRef]
- Leboeuf, R.; Baumgartner, J.E.; Benezra, M.; Malaguarnera, R.; Solit, D.; Pratilas, C.A.; Rosen, N.; Knauf, J.A.; Fagin, J.A. BRAFV600E mutation is associated with preferential sensitivity to mitogen-activated protein kinase kinase inhibition in thyroid cancer cell lines. J. Clin. Endocrinol. Metab. 2008, 93, 2194–2201. [Google Scholar] [CrossRef] [PubMed]
- Ball, D.W.; Jin, N.; Rosen, D.M.; Dackiw, A.; Sidransky, D.; Xing, M.; Nelkin, B.D. Selective growth inhibition in BRAF mutant thyroid cancer by the mitogen-activated protein kinase kinase 1/2 inhibitor AZD6244. J. Clin. Endocrinol. Metab. 2007, 92, 4712–4718. [Google Scholar] [CrossRef] [PubMed]
- Pilli, T.; Prasad, K.V.; Jayarama, S.; Pacini, F.; Prabhakar, B.S. Potential utility and limitations of thyroid cancer cell lines as models for studying thyroid cancer. Thyroid 2009, 19, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J. Understanding the genotype of follicular thyroid tumors. Endocr. Pathol. 2005, 16, 311–321. [Google Scholar] [CrossRef]
- Hassan, I.; Wunderlich, A.; Burchert, A.; Hoffmann, S.; Zielke, A. Antisense p53 oligonucleotides inhibit proliferation and induce chemosensitivity in follicular thyroid cancer cells. Anticancer Res. 2006, 26, 1171–1176. [Google Scholar] [PubMed]
- Liu, D.; Hu, S.; Hou, P.; Jiang, D.; Condouris, S.; Xing, M. Suppression of BRAF/MEK/MAP kinase pathway restores expression of iodide-metabolizing genes in thyroid cells expressing the V600E BRAF mutant. Clin. Cancer Res. 2007, 13, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.J.; Brown, A.P.; Asano, H.; Mandler, M.; Burgan, W.E.; Carter, D.; Camphausen, K.; Citrin, D. In vitro and in vivo radiosensitization with AZD6244 (ARRY-142886), an inhibitor of mitogen-activated protein kinase/extracellular signal-regulated kinase 1/2 kinase. Clin. Cancer Res. 2009, 15, 3050–3057. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, O. Targeting the MEK signaling pathway in non-small cell lung cancer (NSCLC) patients with RAS aberrations. Ther. Adv. Respir. Dis. 2016, 10, 265–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davar, D.; Kirkwood, J.M. CCR 20th Anniversary Commentary: MAPK/ERK Pathway Inhibition in Melanoma-Kinase Inhibition Redux. Clin. Cancer Res. 2015, 21, 5412–5414. [Google Scholar] [CrossRef] [PubMed]
- Maiello, M.R.; D’Alessio, A.; Bevilacqua, S.; Gallo, M.; Normanno, N.; De Luca, A. EGFR and MEK Blockade in Triple Negative Breast Cancer Cells. J. Cell. Biochem. 2015, 116, 2778–2785. [Google Scholar] [CrossRef] [PubMed]
- Sweetlove, M.; Wrightson, E.; Kolekar, S.; Rewcastle, G.W.; Baguley, B.C.; Shepherd, P.R.; Jamieson, S.M. Inhibitors of pan-PI3K Signaling Synergize with BRAF or MEK Inhibitors to Prevent BRAF-Mutant Melanoma Cell Growth. Front. Oncol. 2015, 5, 135. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Wu, X.; Yin, Y.; Yang, Y.; Ma, D.; Li, H. Antitumor activity of selective MEK1/2 inhibitor AZD6244 in combination with PI3K/mTOR inhibitor BEZ235 in gefitinib-resistant NSCLC xenograft models. J. Exp. Clin. Cancer Res. 2014, 33, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Hu, H.Y.; Meng, W.; Jiang, L.; Zhang, X.; Sha, J.J.; Lu, Z.; Yao, Y. MEK inhibitor effective against proliferation in breast cancer cell. Tumour Biol. 2014, 35, 9269–9279. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.; Youn, H.; Yeom, C.J.; Kang, K.W.; Chung, J.K. Glycosylation of Sodium/Iodide Symporter (NIS) Regulates Its Membrane Translocation and Radioiodine Uptake. PLoS ONE 2015, 10, e0142984. [Google Scholar] [CrossRef] [PubMed]
- Kogai, T.; Brent, G.A. The sodium iodide symporter (NIS): Regulation and approaches to targeting for cancer therapeutics. Pharmacol. Ther. 2012, 135, 355–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachter, S.; Wunderlich, A.; Roth, S.; Mintziras, I.; Maurer, E.; Hoffmann, S.; Verburg, F.A.; Fellinger, S.A.; Holzer, K.; Bartsch, D.K.; et al. Individualised Multimodal Treatment Strategies for Anaplastic and Poorly Differentiated Thyroid Cancer. J. Clin. Med. 2018, 7, 115. [Google Scholar] [CrossRef] [PubMed]
- Wachter, S.; Damanakis, A.I.; Elxnat, M.; Roth, S.; Wunderlich, A.; Verburg, F.A.; Fellinger, S.A.; Bartsch, D.K.; Di Fazio, P. Epigenetic Modifications in Thyroid Cancer Cells Restore NIS and Radio-Iodine Uptake and Promote Cell Death. J. Clin. Med. 2018, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Angell, T.E.; Lechner, M.G.; Jang, J.K.; LoPresti, J.S.; Epstein, A.L. MHC class I loss is a frequent mechanism of immune escape in papillary thyroid cancer that is reversed by interferon and selumetinib treatment in vitro. Clin. Cancer Res. 2014, 20, 6034–6044. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Seyama, T.; Iwamoto, K.S.; Hayashi, T.; Mizuno, T.; Tsuyama, N.; Dohi, K.; Nakamura, N.; Akiyama, M. In vitro irradiation is able to cause RET oncogene rearrangement. Cancer Res. 1993, 53, 2940–2943. [Google Scholar] [PubMed]
- Ishizaka, Y.; Itoh, F.; Tahira, T.; Ikeda, I.; Ogura, T.; Sugimura, T.; Nagao, M. Presence of aberrant transcripts of ret proto-oncogene in a human papillary thyroid carcinoma cell line. Jpn. J. Cancer Res. 1989, 80, 1149–1152. [Google Scholar] [CrossRef] [PubMed]
- Heldin, N.E.; Westermark, B. The molecular biology of the human anaplastic thyroid carcinoma cell. Thyroidology 1991, 3, 127–131. [Google Scholar] [PubMed]
- Fabien, N.; Fusco, A.; Santoro, M.; Barbier, Y.; Dubois, P.M.; Paulin, C. Description of a human papillary thyroid carcinoma cell line. Morphologic study and expression of tumoral markers. Cancer 1994, 73, 2206–2212. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Contrast | Estimate | Lower | Upper | p |
|---|---|---|---|---|---|
| TPC1 | 0.1 vs. control | −0.07 | −0.12 | −0.01 | 0.02 |
| TPC1 | 1.0 vs. control | −0.10 | −0.16 | −0.04 | 0.00 |
| TPC1 | 10 vs. control | −0.14 | −0.22 | −0.07 | 0.00 |
| TPC1 | 100 vs. control | −0.21 | −0.30 | −0.13 | 0.00 |
| C643 | 0.1 vs. control | −0.12 | −0.24 | −0.00 | 0.04 |
| C643 | 1.0 vs. control | −0.34 | −0.48 | −0.20 | 0.00 |
| C643 | 10 vs. control | −0.66 | −0.87 | −0.46 | 0.00 |
| C643 | 100 vs. control | −1.01 | −1.32 | −0.70 | 0.00 |
| BCPAP | 0.1 vs. control | −0.12 | −0.22 | −0.02 | 0.02 |
| BCPAP | 1.0 vs. control | −0.17 | −0.29 | −0.06 | 0.00 |
| BCPAP | 10 vs. control | −0.19 | −0.32 | −0.07 | 0.00 |
| BCPAP | 100 vs. control | −0.29 | −0.44 | −0.14 | 0.00 |
| 8505C | 0.1 vs. control | −0.19 | −0.23 | −0.15 | 0.00 |
| 8505C | 1.0 vs. control | −0.30 | −0.36 | −0.24 | 0.00 |
| 8505C | 10 vs. control | −0.33 | −0.40 | −0.26 | 0.00 |
| 8505C | 100 vs. control | −0.34 | −0.42 | −0.28 | 0.00 |
| hsa-let7f-5p | hsa-miR-146b-5p | hsa-miR-146b-3p | SLC5A5 | |
|---|---|---|---|---|
| TPC1 | ↓ | ↓ | n.d. | ↑ |
| C643 | ↑ | ±/↑ | ↑ | ± |
| BCPAP | ↓ | ↓ | ↓ | ↑ |
| 8505C | ±/↑ | ↓ | ±/↑ | ↑ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wächter, S.; Wunderlich, A.; Greene, B.H.; Roth, S.; Elxnat, M.; Fellinger, S.A.; Verburg, F.A.; Luster, M.; Bartsch, D.K.; Di Fazio, P. Selumetinib Activity in Thyroid Cancer Cells: Modulation of Sodium Iodide Symporter and Associated miRNAs. Int. J. Mol. Sci. 2018, 19, 2077. https://doi.org/10.3390/ijms19072077
Wächter S, Wunderlich A, Greene BH, Roth S, Elxnat M, Fellinger SA, Verburg FA, Luster M, Bartsch DK, Di Fazio P. Selumetinib Activity in Thyroid Cancer Cells: Modulation of Sodium Iodide Symporter and Associated miRNAs. International Journal of Molecular Sciences. 2018; 19(7):2077. https://doi.org/10.3390/ijms19072077
Chicago/Turabian StyleWächter, Sabine, Annette Wunderlich, Brandon H. Greene, Silvia Roth, Moritz Elxnat, Sebastian A. Fellinger, Frederik A. Verburg, Markus Luster, Detlef K. Bartsch, and Pietro Di Fazio. 2018. "Selumetinib Activity in Thyroid Cancer Cells: Modulation of Sodium Iodide Symporter and Associated miRNAs" International Journal of Molecular Sciences 19, no. 7: 2077. https://doi.org/10.3390/ijms19072077
APA StyleWächter, S., Wunderlich, A., Greene, B. H., Roth, S., Elxnat, M., Fellinger, S. A., Verburg, F. A., Luster, M., Bartsch, D. K., & Di Fazio, P. (2018). Selumetinib Activity in Thyroid Cancer Cells: Modulation of Sodium Iodide Symporter and Associated miRNAs. International Journal of Molecular Sciences, 19(7), 2077. https://doi.org/10.3390/ijms19072077