Genome-Wide Identification and Characterization of CIPK Family and Analysis Responses to Various Stresses in Apple (Malus domestica)


Abstract
:1. Introduction
2. Results
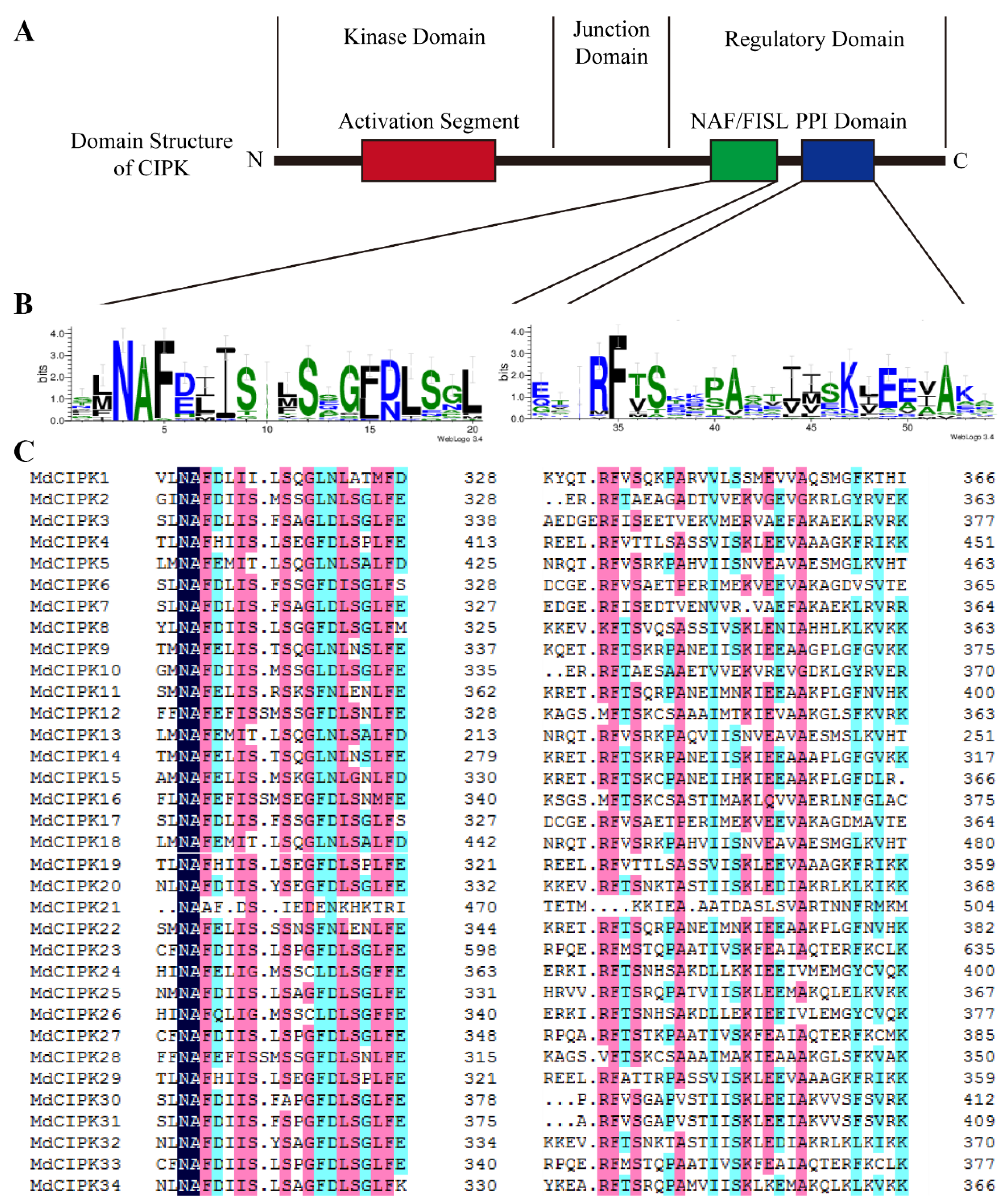
2.1. Identification of CIPK Family Genes in Apple
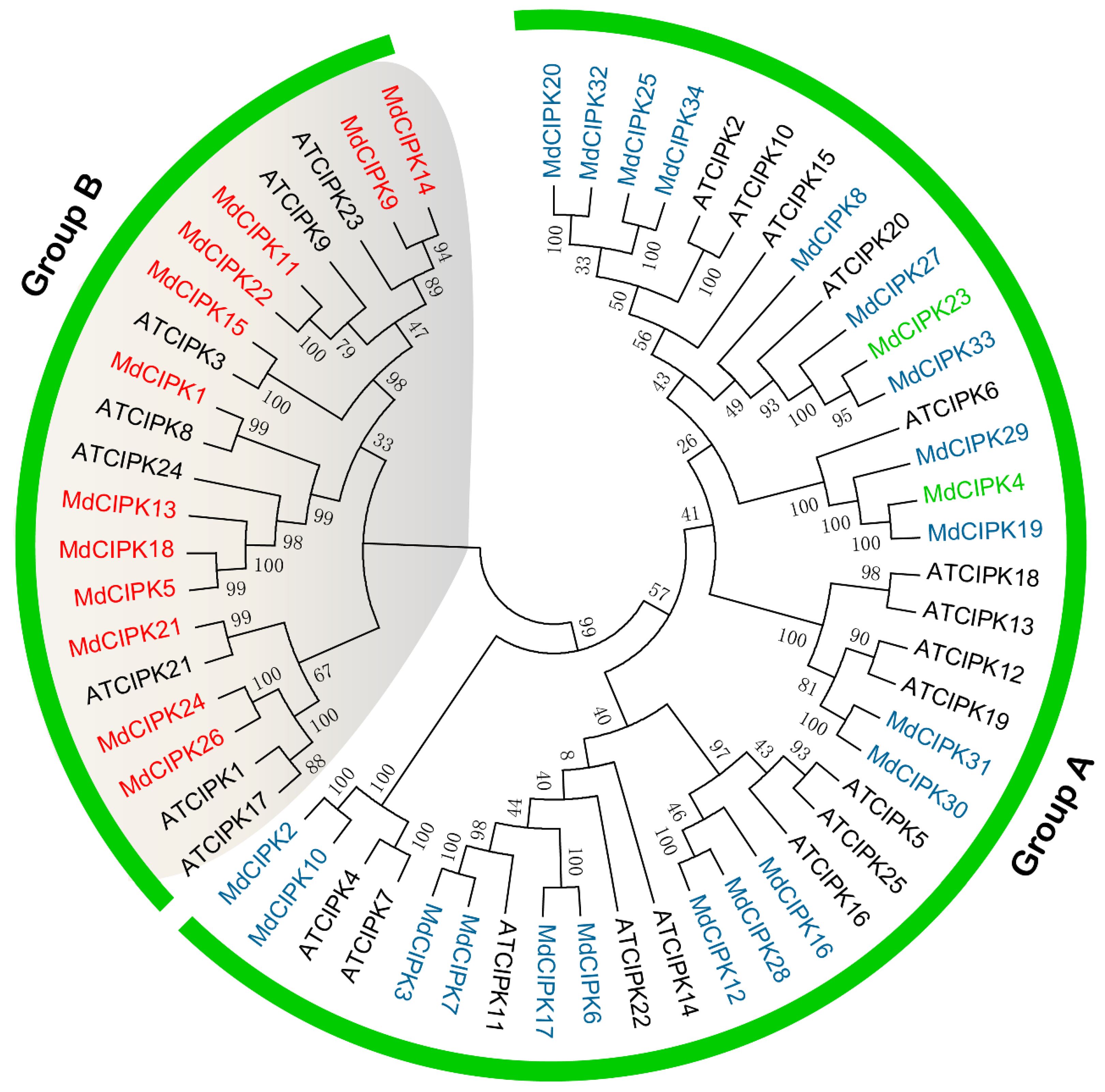
2.2. Phylogenetic Analysis of the MdCIPK Family Genes
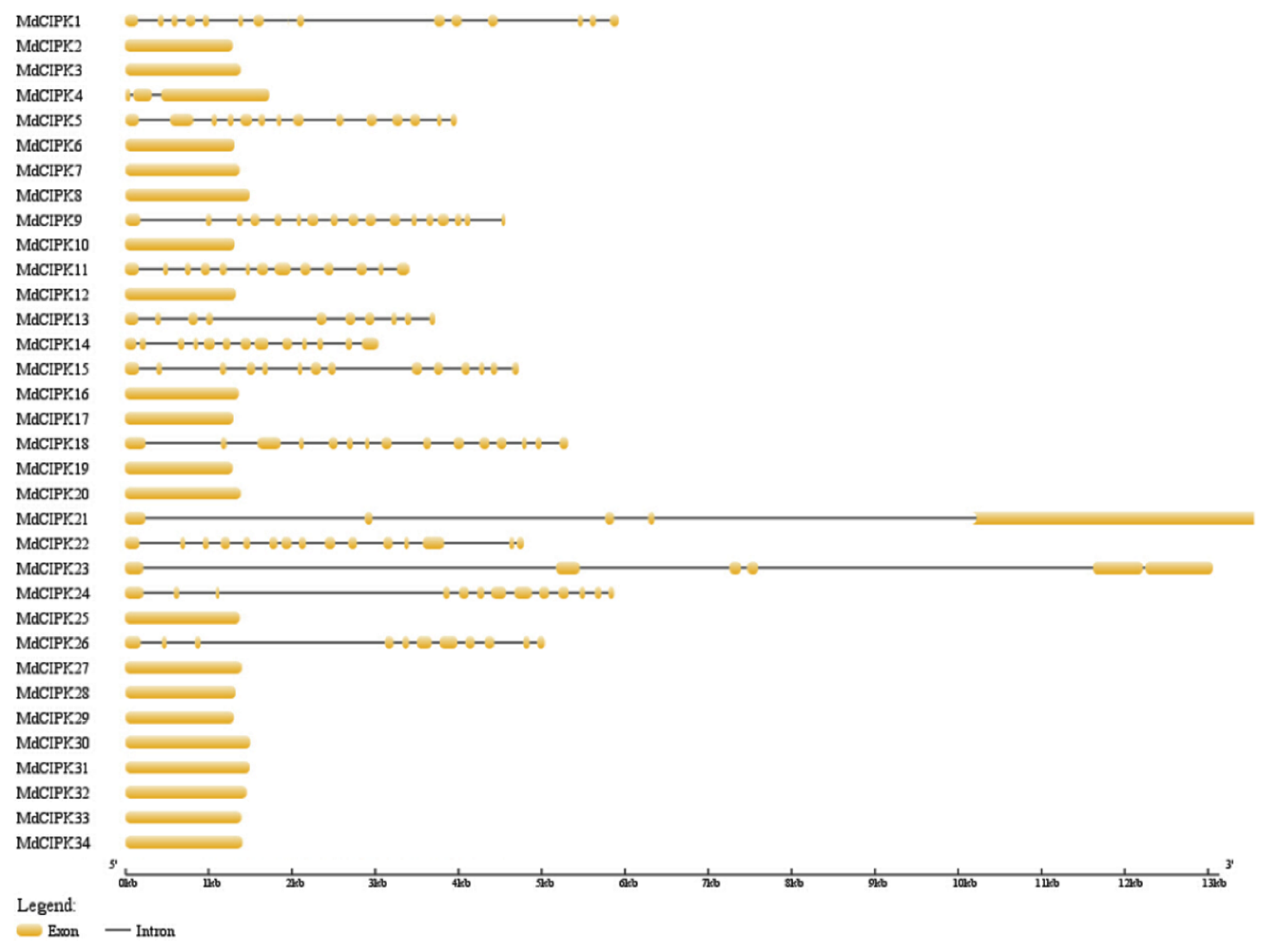
2.3. Gene Structure of CIPK Genes in Apple
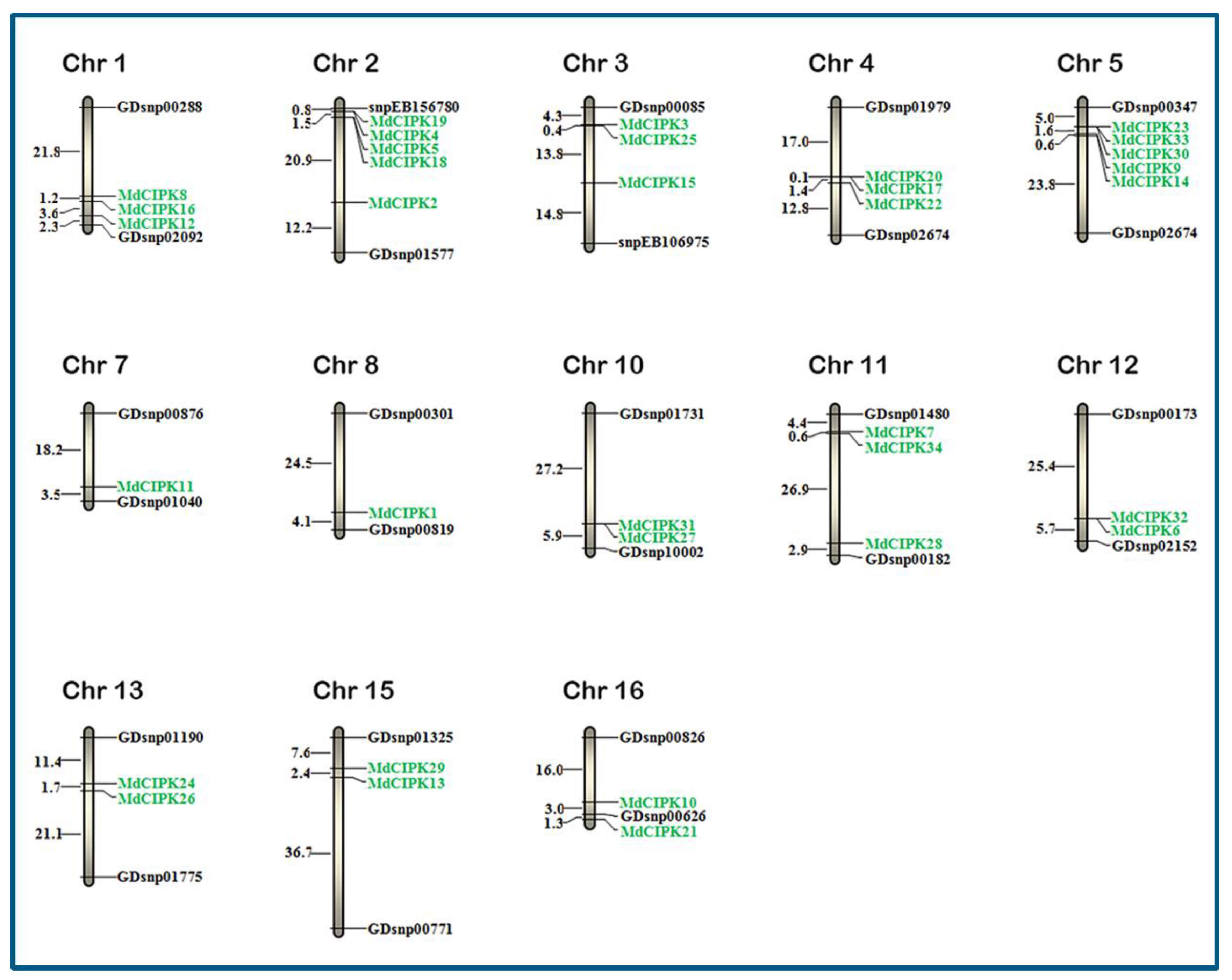
2.4. Chromosomal Distribution of the MdCIPK Family
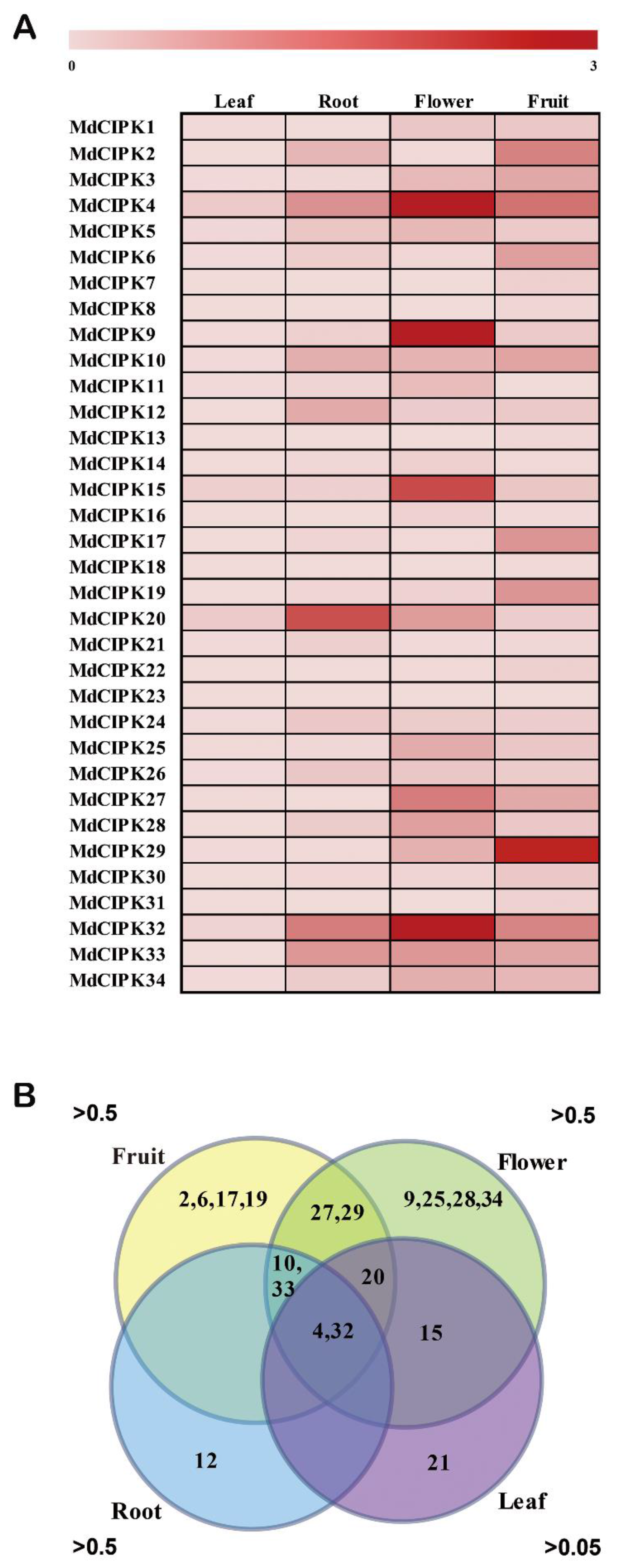
2.5. Expression Patterns of MdCIPK Genes in Different Tissues
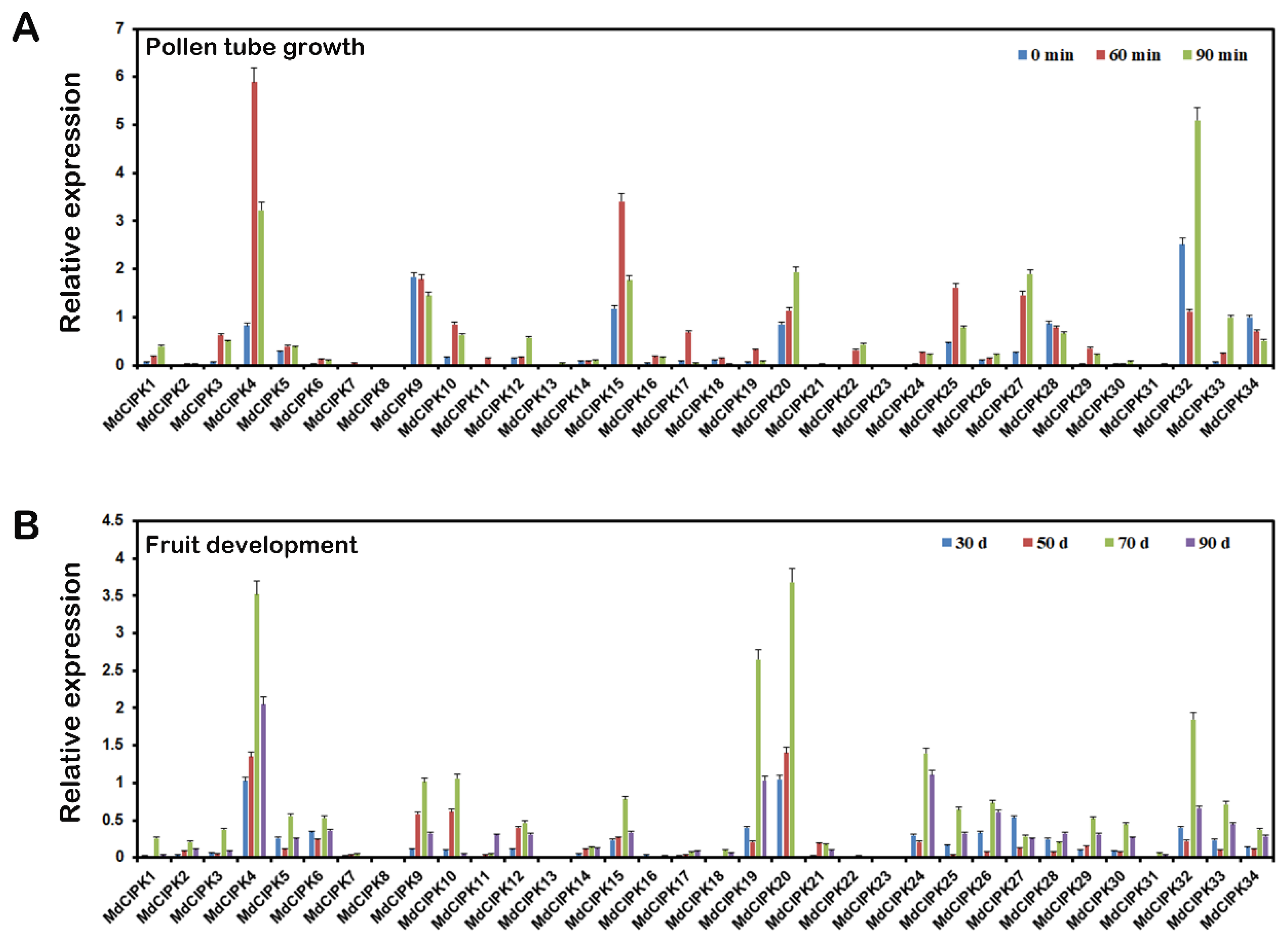
2.6. Expression Analysis of MdCIPK Genes at Different Stages in Apple Pollen Tube Growth and Fruit Development
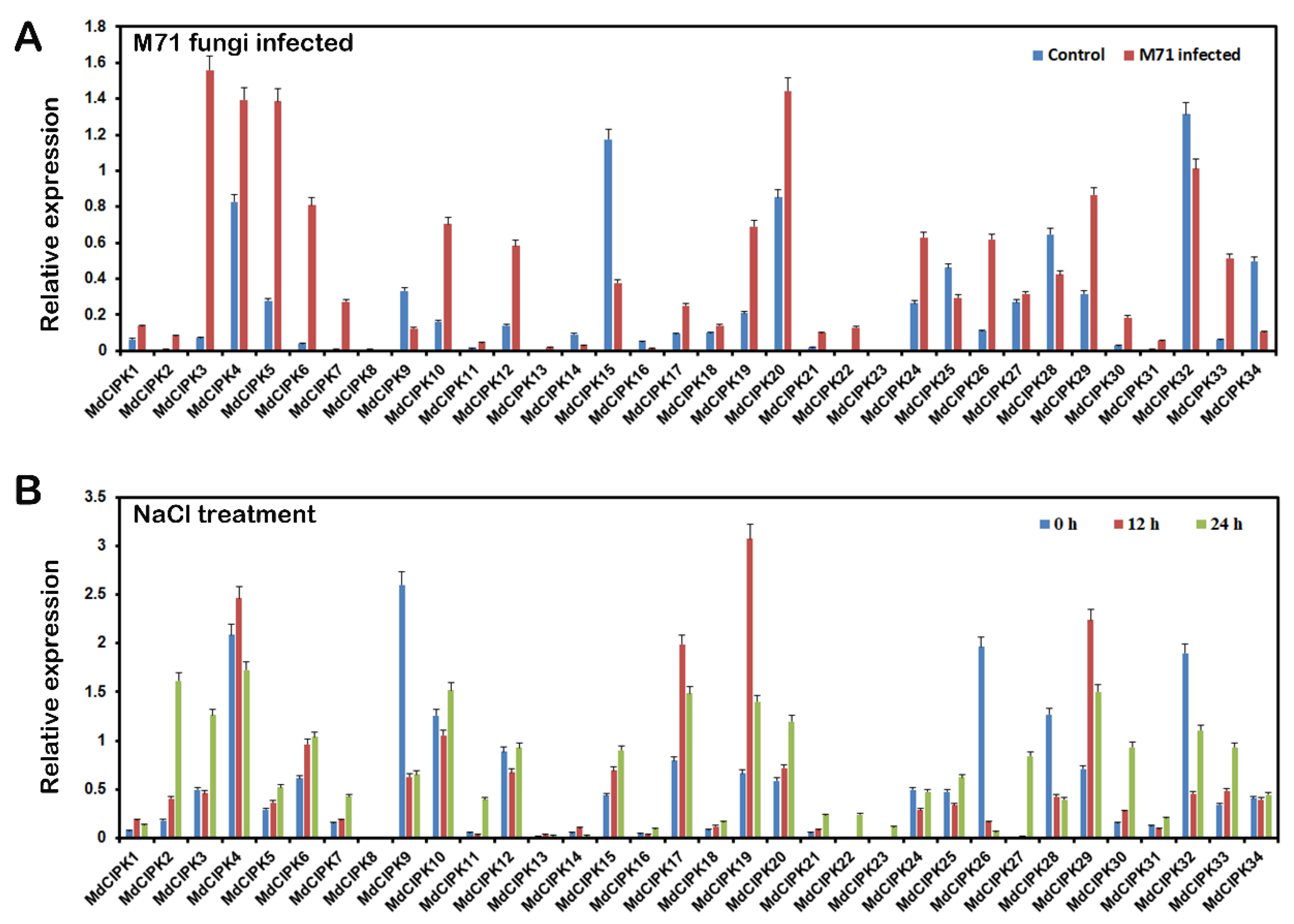
2.7. Expression Analysis of MdCIPK Genes in Response to Biotic Stress and Abiotic Stress
3. Discussion
4. Materials and Methods
4.1. Database Searches and Identification of the CIPK Family in Apple
4.2. Phylogenetic Analysis and Chromosomal Locations of the CIPK Family
4.3. Plant Materials, Growth Conditions and Treatments
4.4. RNA Isolation and Quantitative Real-Time PCR
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primers (5’-3’) | Group | |
|---|---|---|---|
| Forward Primer | Reverse Primers | ||
| MdCIPK1 | GATGATGTGAATGCTGTTTT | ATAGTTGCGAATGTGTGTCT | B |
| MdCIPK2 | AAGCCCCAGAATCTTTTGC | TTGAGCCGTCGTAGCCG | A |
| MdCIPK3 | GAAGGTGGATGTGTGGTC | ACAATCGTCGTCGTAGAAT | A |
| MdCIPK4 | CCGTGGTTCTCGTCTGA | TCCTGCTCCTGTTTCGT | A |
| MdCIPK5 | ATCCCAATCCCAAAACTCG | TCTGACCGCTCCGCTACA | B |
| MdCIPK6 | ATTTTGAAGGACCCGTGG | ACTTGCTAAACAACCCCGA | A |
| MdCIPK7 | TCCCAGACAATGCCCT | GCCTTTCACGAACTCCA | A |
| MdCIPK8 | TGTCTGGCTTGTTTATGGG | TCTTGTTTCTGCTCACCGT | A |
| MdCIPK9 | AGAAGGTCCTCAAGCAC | TCTCCACCAGTCACAAA | B |
| MdCIPK10 | CGGCGTCATCCTCTTCG | TCGTGTTTGGGTTCGGG | A |
| MdCIPK11 | TTTGTTGATGGGGGTGA | GGAGAAGATTTTCGGGC | B |
| MdCIPK12 | GGAACGACGGGCTACT | TCCGCACGACCAGAG | A |
| MdCIPK13 | TGATTACCCTATCCCAAG | TCAAGAGTATCTCCTGCG | B |
| MdCIPK14 | TTTGGATTGAGTGCCCTA | CCCTTTGTTGTTGATTACCT | B |
| MdCIPK15 | CCTAACCCTATGACTCGTG | TTGTTCTTCCTTTTTTTCC | B |
| MdCIPK16 | GTGAGGAGGATGTGAGGC | TAATGGCAGTTAGGGTTTG | A |
| MdCIPK17 | GGAGGCTTCTGGGCTG | CACGATATGCGGGTGG | A |
| MdCIPK18 | GACTTAGGCAACCTCAGC | TTTCCCACAAAAATCACA | B |
| MdCIPK19 | CCATAGATTTCTGCCAC | TCGTATCCTTTTTTTCC | A |
| MdCIPK20 | TGTTCTATTGGCTGGCTAT | GAGTATTTGGATTGGGGT | A |
| MdCIPK21 | CTGACAGACAAAGCCG | GACGATGAATAACCGC | B |
| MdCIPK22 | TGGTTACCTGCCTTTT | TCTCTTTCCTTTCCGTT | B |
| MdCIPK23 | CGAAAGGTTTGAGTGA | TAAGAAGGTATCCGAGT | A |
| MdCIPK24 | GCCAAGGGGAACATAAA | AACCTCGGGAGCAACA | B |
| MdCIPK25 | TGGCAACCAAAACTAAGA | ATCCCGATGATAAACACC | A |
| MdCIPK26 | AGACCGAAGGCAGGA | GCGCTAAGGCCAAAGT | B |
| MdCIPK27 | AGGTCACGCCTTCATTT | GCTGTTGCTGCTCTGTTC | A |
| MdCIPK28 | GGAGAAGCGAATCACCAT | GTCGGACTTTTCATCAACA | A |
| MdCIPK29 | GCGAGCAAGTCCAAGAT | AAAGCACTGAGTCCGAAA | A |
| MdCIPK30 | CTCTGCTTCTGGGTCG | ATCTTGGCTTTCGTGG | A |
| MdCIPK31 | GCCCTGCTCTGCTTCT | CCTTGGCGACCTTGTT | A |
| MdCIPK32 | AGAAAAAGGAAGTGCGG | CCAGATGAAAAGACGGG | A |
| MdCIPK33 | GGTTGCCAAAGGGAAG | CGTGGTGTGGAGGAGG | A |
| MdCIPK34 | AGATTATGAAGGTCGGG | TCTTAGTTTTGGTTGCC | A |
| Actin | TGACCGAATGAGCAAGGAAATTACT | TACTCAGCTTTGGCAATCCACATC | A |
References
- Sanders, D.; Pelloux, J.; Brownlee, C.; Harper, J.F. Calcium at the Crossroads of Signaling. Plant Cell 2002, 14 (Suppl. 1), S401–S417. [Google Scholar] [CrossRef] [PubMed]
- Evans, N.H.; McAinsh, M.R.; Hetherington, A.M. Calcium oscillations in higher plants. Curr. Opin. Plant Biol. 2001, 4, 415–420. [Google Scholar] [CrossRef]
- Harper, J.F. Dissecting calcium oscillators in plant cells. Trends Plant Sci. 2001, 6, 395–397. [Google Scholar] [CrossRef]
- Yang, T.B.; Poovaiah, B.W. Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci. 2003, 8, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.C.; Shin, D.; Lee, J.; Kang, C.H.; Baek, D.; Cho, M.J.; Kim, M.C.; Yun, D.J. Isolation and characterization of a novel Calcium/Calmodulin-Dependent protein kinase, AtCK, from Arabidopsis. Mol. Cells 2007, 24, 276–282. [Google Scholar] [PubMed]
- Luan, S.; Lan, W.Z.; Lee, S.C. Potassium nutrition, sodium toxicity, and calcium signaling: Connections through the CBL-CIPK network. Curr. Opin. Plant Biol. 2009, 12, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Hrabak, E.M.; Chan, C.W.M.; Gribskov, M.; Harper, J.F.; Choi, J.H.; Halford, N.; Kudla, J.; Luan, S.; Nimmo, H.G.; Sussman, M.R.; et al. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol. 2003, 132, 666–680. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Sopory, S.K.; Tuteja, N. Cloning and characterization of CBL-CIPK signalling components from a legume (Pisum sativum). FEBS J. 2006, 273, 907–925. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wang, Y.; Wang, M.; Li, T.; Zhou, Y.; Wang, X.; Wei, S.; He, G.; Yang, G. Identification and comprehensive analyses of the CBL and CIPK gene families in wheat (Triticum aestivum L.). BMC Plant Biol. 2015, 15, 269. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Wang, Q.L.; Chen, Q.; Xiang, N.; Yang, Y.Q.; Yang, Y.P. Genome-Wide Identification and Functional Analysis of the Calcineurin B-like Protein and Calcineurin B-like Protein-Interacting Protein Kinase Gene Families in Turnip (Brassica rapa var. rapa). Front. Plant Sci. 2017, 8, 1191. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.F.; Sun, Z.F.; Zheng, J.; Guo, X.Y.; Dong, Z.G.; Huai, J.L.; Gou, M.Y.; He, J.G.; Jin, Y.S.; Wang, J.H.; et al. Cloning and characterization of a novel CBL-interacting protein kinase from maize. Plant Mol. Biol. 2009, 69, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Wei, Q.; Wang, R.; Zhang, Y.; Zhang, F.; He, Y.; Yang, G.; He, G. Ectopic expression of BdCIPK31 confers enhanced low-temperature tolerance in transgenic tobacco plants. Acta Biochim. Biophys. Sin. (Shanghai) 2018, 50, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Wei, Q.; Wang, R.; Zhang, Y.; Zhang, F.; He, Y.; Zhou, S.; Feng, J.; Yang, G.; He, G. BdCIPK31, a Calcineurin B-Like Protein-Interacting Protein Kinase, Regulates Plant Response to Drought and Salt Stress. Front. Plant Sci. 2017, 8, 1184. [Google Scholar] [CrossRef] [PubMed]
- Mo, C.; Wan, S.; Xia, Y.; Ren, N.; Zhou, Y.; Jiang, X. Expression Patterns and Identified Protein-Protein Interactions Suggest That Cassava CBL-CIPK Signal Networks Function in Responses to Abiotic Stresses. Front. Plant Sci. 2018, 9, 269. [Google Scholar] [CrossRef] [PubMed]
- Dimick, P.S.; Hoskin, J.C. Review of apple flavor—State of the art. Crit. Rev. Food Sci. Nutr. 1983, 18, 387–409. [Google Scholar] [CrossRef] [PubMed]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus x domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolukisaoglu, U.; Weinl, S.; Blazevic, D.; Batistic, O.; Kudla, J. Calcium sensors and their interacting protein kinases: Genomics of the Arabidopsis and rice CBL-CIPK signaling networks. Plant Physiol. 2004, 134, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.F.; Gu, Z.M.; Xin, D.D.; Hao, L.A.; Liu, C.J.; Huang, J.; Ma, B.J.; Zhang, H.S. Identification and characterization of putative CIPK genes in maize. J. Genet. Genom. 2011, 38, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, P.; Sanyal, S.K.; Tokas, I.; Yadav, A.K.; Pandey, A.; Kapoor, S.; Pandey, G.K. Comprehensive structural, interaction and expression analysis of CBL and CIPK complement during abiotic stresses and development in rice. Cell Calcium 2014, 56, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, B.; Liu, W.-Z.; Li, H.; Wang, L.; Wang, B.; Deng, M.; Liang, W.; Deyholos, M.K.; Jiang, Y.-Q. Identification and characterization of CBL and CIPK gene families in canola (Brassica napus L.). BMC Plant Biol. 2014, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Xia, X.; Yin, W.; Zhang, H. Comparative genomic analysis of CIPK gene family in Arabidopsis and Populus. Plant Growth Regul. 2007, 52, 101–110. [Google Scholar] [CrossRef]
- Hu, W.; Xia, Z.; Yan, Y.; Ding, Z.; Tie, W.; Wang, L.; Zou, M.; Wei, Y.; Lu, C.; Hou, X.; et al. Genome-wide gene phylogeny of CIPK family in cassava and expression analysis of partial drought-induced genes. Front. Plant Sci. 2015, 6, 914. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Lin, J.; Li, H.; Li, X.; Yang, Q.; Cheng, Z.-M.; Chang, Y. Characterization of CIPK Family in Asian Pear (Pyrus bretschneideri Behd) and Co-expressin Analysis Related to Salt and Osmotic Stress Responses. Front. Plant Sci. 2016, 7, 1361. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, Q.; Yu, H.; Zhou, T.; Zou, J.; Zhang, H.; Bian, M.; Liu, X. Characterization of Alkali Stress-Responsive Genes of the CIPK Family in Sweet Sorghum Sorghum bicolor (L.) Moench. Crop Sci. 2015, 55, 1254–1263. [Google Scholar] [CrossRef]
- Hu, D.G.; Ma, Q.J.; Sun, C.H.; Sun, M.H.; You, C.X.; Hao, Y.J. Overexpression of MdSOS2L1, a CIPK protein kinase, increases the antioxidant metabolites to enhance salt tolerance in apple and tomato. Physiol. Plant. 2016, 156, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.K.; Li, L.L.; Cao, Z.H.; Zhao, Q.; Li, M.; Zhang, L.Y.; Hao, Y.J. Molecular cloning and functional characterization of a novel apple MdCIPK6L gene reveals its involvement in multiple abiotic stress tolerance in transgenic plants. Plant Mol. Biol. 2012, 79, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Meng, D.; Gu, Z.; Li, W.; Chen, Q.; Li, Y.; Yuan, H.; Yu, J.; Liu, C.; Li, T. Apple S-RNase interacts with an actin-binding protein, MdMVG, to reduce pollen tube growth by inhibiting its actin-severing activity at the early stage of self-pollination induction. Plant J. 2018, 95, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Liu, J.Y.; Dong, C.; Cheng, Z.M. The CBL and CIPK Gene Family in Grapevine (Vitis vinifera): Genome-Wide Analysis and Expression Profiles in Response to Various Abiotic Stresses. Front. Plant Sci. 2017, 8, 978. [Google Scholar] [CrossRef] [PubMed]
- Kidokoro, S.; Watanabe, K.; Ohori, T.; Moriwaki, T.; Maruyama, K.; Mizoi, J.; Htwe, N.M.P.S.; Fujita, Y.; Sekita, S.; Shinozaki, K.; et al. Soybean DREB1/CBF-type transcription factors function in heat and drought as well as cold stress-responsive gene expression. Plant J. 2015, 81, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; He, M.Y.; Bai, Y.; Xu, H.X.; Dandekar, A.M.; Fei, Z.J.; Cheng, L.L. Decreased sorbitol synthesis leads to abnormal stamen development and reduced pollen tube growth via an MYB transcription factor, MdMYB39L, in apple (Malus domestica). New Phytol. 2018, 217, 641–656. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-Y.; Meng, D.; Li, M.; Cheng, L. Genome-wide identification and expression analysis of the bZIP gene family in apple (Malus domestica). Tree Genet. Genomes 2016, 12, 82. [Google Scholar] [CrossRef]
- Chaves-Sanjuan, A.; Jose Sanchez-Barrena, M.; Maria Gonzalez-Rubio, J.; Moreno, M.; Ragel, P.; Jimenez, M.; Pardo, J.M.; Martinez-Ripoll, M.; Quintero, F.J.; Albert, A. Structural basis of the regulatory mechanism of the plant CIPK family of protein kinases controlling ion homeostasis and abiotic stress. Proc. Natl. Acad. Sci. USA 2014, 111, E4532–E4541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manik, S.M.N.; Shi, S.J.; Mao, J.J.; Dong, L.H.; Su, Y.L.; Wang, Q.; Liu, H.B. The Calcium Sensor CBL-CIPK Is Involved in Plant’s Response to Abiotic Stresses. Int. J. Genom. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.K.; Kanwar, P.; Singh, A.; Steinhorst, L.; Pandey, A.; Yadav, A.K.; Tokas, I.; Sanyal, S.K.; Kim, B.G.; Lee, S.C.; et al. Calcineurin B-Like Protein-Interacting Protein Kinase CIPK21 Regulates Osmotic and Salt Stress Responses in Arabidopsis. Plant Physiol. 2015, 169, 780–792. [Google Scholar] [CrossRef] [PubMed]
- Monerri, C.; Fortunato-Almeida, A.; Molina, R.V.; Nebauer, S.G.; Garcia-Luis, A.; Guardiola, J.L. Relation of carbohydrate reserves with the forthcoming crop, flower formation and photosynthetic rate, in the alternate bearing ‘Salustiana’ sweet orange (Citrus sinensis L.). Sci. Hortic. (Amst.) 2011, 129, 71–78. [Google Scholar] [CrossRef]
- Glazebrook, J. Genes controlling expression of defense responses in Arabidopsis—2001 status. Curr. Opin. Plant Biol. 2001, 4, 301–308. [Google Scholar] [CrossRef]
- Chen, W.; Provart, N.J.; Glazebrook, J.; Katagiri, F.; Chang, H.-S.; Eulgem, T.; Mauch, F.; Luan, S.; Zou, G.; Whitham, S.A.; et al. Expression Profile Matrix of Arabidopsis Transcription Factor Genes Suggests Their Putative Functions in Response to Environmental Stresses. Plant Cell 2002, 14, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Liang, D.; Wang, P.; Liu, J.Y.; Ma, F.W. Genome-wide analysis and expression profiling of the DREB transcription factor gene family in Malus under abiotic stress. Mol. Genet. Genom. 2012, 287, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Su, H.Y.; Zhang, S.Z.; Yuan, X.W.; Chen, C.T.; Wang, X.F.; Hao, Y.J. Genome-wide analysis and identification of stress-responsive genes of the NAM-ATAF1,2-CUC2 transcription factor family in apple. Plant Physiol. Biochem. 2013, 71, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Ma, Q.J.; Sun, M.H.; Lu, J.; Liu, Y.J.; You, C.X.; Hao, Y.J. An apple CIPK protein kinase targets a novel residue of AREB transcription factor for ABA-dependent phosphorylation. Plant Cell Environ. 2017, 40, 2207–2219. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korban, S.S.; Gasic, K.; Hernandez, A.G. RNA Extraction from Different Apple Tissues Rich in Polyphenols and Polysaccharides for cDNA Library Construction. Plant Mol. Biol. Rep. 2004, 22, 437–438. [Google Scholar]
- Johnson, R.D.; Johnson, L.; Itoh, Y.; Kodama, M.; Otani, H.; Kohmoto, K. Cloning and Characterization of a Cyclic Peptide Synthetase Gene from Alternaria alternata Apple Pathotype Whose Product Is Involved in AM-Toxin Synthesis and Pathogenicity. Mol. Plant Microbe Interact. MPMI 2000, 13, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Li, C.; Park, H.J.; Gonzalez, J.; Wang, J.; Dandekar, A.M.; Turgeon, B.G.; Cheng, L. Sorbitol Modulates Resistance to Alternaria alternata by Regulating the Expression of an NLR Resistance Gene in apple. Plant Cell 2018. [Google Scholar] [CrossRef] [PubMed]
- Udvardi, M.K.; Czechowski, T.; Scheible, W.R. Eleven golden rules of quantitative RT-PCR. Plant Cell 2008, 20, 1736–1737. [Google Scholar] [CrossRef] [PubMed]







| Gene Name | Gene ID | Chromosome | Amino Acids | Intron Number | Isoelectric Point | Molecular Weight (Da) |
|---|---|---|---|---|---|---|
| MdCIPK1 | MDP0000060056 | chr8:25,070,919..25,076,787 | 448 | 13 | 6.91 | 50,892.45 |
| MdCIPK2 | MDP0000126770 | chr2:23,966,243..23,967,541 | 433 | 0 | 9.61 | 46,836.24 |
| MdCIPK3 | MDP0000127732 | chr3:5,028,677..5,030,050 | 462 | 0 | 8.78 | 51,004.81 |
| MdCIPK4 | MDP0000129860 | chr2:1,523,050..1,524,779 | 527 | 2 | 9.68 | 59,044.53 |
| MdCIPK5 | MDP0000136806 | chr2:3,009,347..3,013,324 | 515 | 13 | 7.68 | 57,246.77 |
| MdCIPK6 | MDP0000154855 | chr12:25,770,656..25,771,963 | 436 | 0 | 6.81 | 48,997.12 |
| MdCIPK7 | MDP0000158876 | chr11:4,897,105..4,898,478 | 458 | 0 | 8.64 | 49,937.29 |
| MdCIPK8 | MDP0000165729 | chr1:21,759,311..21,760,77 | 497 | 0 | 9.52 | 48,611.3 |
| MdCIPK9 | MDP0000167597 | chr5:6,994,115..6,998,675 | 529 | 16 | 9.21 | 59,399.19 |
| MdCIPK10 | MDP0000197160 | chr16:16,136,654..16,138,184 | 441 | 0 | 8.97 | 48,655.04 |
| MdCIPK11 | MDP0000216765 | chr7:18,303,752..18,307,186 | 475 | 12 | 9.04 | 54,320.72 |
| MdCIPK12 | MDP0000226074 | chr1:26,489,681..26,491,021 | 447 | 0 | 8.66 | 50,322.2 |
| MdCIPK13 | MDP0000230405 | chr15:10,561,049..10,564,790 | 322 | 9 | 9.27 | 36,721.29 |
| MdCIPK14 | MDP0000233288 | chr5:7,626,506..7,629,56 | 458 | 12 | 6.92 | 48,761.51 |
| MdCIPK15 | MDP0000247429 | chr3:19,162,326..19,167,078 | 433 | 13 | 6.14 | 49,252.67 |
| MdCIPK16 | MDP0000249604 | chr1:22,923,293..22,924,672 | 460 | 0 | 9.16 | 52,034.2 |
| MdCIPK17 | MDP0000256347 | chr4:17,280,883..17,282,190 | 436 | 0 | 7.98 | 49,048.48 |
| MdCIPK18 | MDP0000270573 | chr2:3,025,820..3,031,167 | 564 | 14 | 8.37 | 63,209.44 |
| MdCIPK19 | MDP0000277672 | chr2:1,521,829..1,523,127 | 433 | 0 | 9.05 | 48,498.09 |
| MdCIPK20 | MDP0000278839 | chr4:17,229,494..17,230,894 | 467 | 0 | 8.08 | 52,701.59 |
| MdCIPK21 | MDP0000295392 | chr16:20,465,758..20,486,041 | 604 | 4 | 8.34 | 68,780.45 |
| MdCIPK22 | MDP0000300274 | chr4:18,693,218..18,698,036 | 538 | 14 | 6.56 | 60,699.97 |
| MdCIPK23 | MDP0000303485 | chr5:5,393,626..5,406,764 | 729 | 5 | 8.37 | 80,861.01 |
| MdCIPK24 | MDP0000303735 | chr13:11,546,579..11,552,484 | 471 | 12 | 8.71 | 52,592.3 |
| MdCIPK25 | MDP0000313460 | chr3:5,407,721..5,409,109 | 463 | 0 | 8.88 | 52,107.05 |
| MdCIPK26 | MDP0000314872 | chr13:13,253,909..13,258,980 | 437 | 10 | 6.17 | 48,798.65 |
| MdCIPK27 | MDP0000320872 | chr10:27,304,369..27,305,781 | 471 | 0 | 8.80 | 52,483.4 |
| MdCIPK28 | MDP0000614281 | chr14331:32,383,407..32,384,729 | 441 | 0 | 8.60 | 49,556.15 |
| MdCIPK29 | MDP0000632173 | chr14975:8,175,823..8,177,121 | 433 | 0 | 9.12 | 48,593.15 |
| MdCIPK30 | MDP0000695512 | chr5464:5,444,319..5,445,815 | 499 | 0 | 8.21 | 55,470.78 |
| MdCIPK31 | MDP0000711750 | chr10:27,296,189..27,297,679 | 497 | 0 | 7.58 | 55,038.17 |
| MdCIPK32 | MDP0000747045 | chr12:25,730,471..25,731,922 | 484 | 0 | 8.71 | 54,632.11 |
| MdCIPK33 | MDP0000796828 | chr5:5,437,782..5,439,173 | 464 | 0 | 8.82 | 51,760.44 |
| MdCIPK34 | MDP0000859587 | chr11:5,489,130..5,490,533 | 468 | 0 | 8.85 | 52,661.9 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, L.; Dong, B.; Song, Z.; Meng, D.; Fu, Y. Genome-Wide Identification and Characterization of CIPK Family and Analysis Responses to Various Stresses in Apple (Malus domestica). Int. J. Mol. Sci. 2018, 19, 2131. https://doi.org/10.3390/ijms19072131
Niu L, Dong B, Song Z, Meng D, Fu Y. Genome-Wide Identification and Characterization of CIPK Family and Analysis Responses to Various Stresses in Apple (Malus domestica). International Journal of Molecular Sciences. 2018; 19(7):2131. https://doi.org/10.3390/ijms19072131
Chicago/Turabian StyleNiu, Lili, Biying Dong, Zhihua Song, Dong Meng, and Yujie Fu. 2018. "Genome-Wide Identification and Characterization of CIPK Family and Analysis Responses to Various Stresses in Apple (Malus domestica)" International Journal of Molecular Sciences 19, no. 7: 2131. https://doi.org/10.3390/ijms19072131
APA StyleNiu, L., Dong, B., Song, Z., Meng, D., & Fu, Y. (2018). Genome-Wide Identification and Characterization of CIPK Family and Analysis Responses to Various Stresses in Apple (Malus domestica). International Journal of Molecular Sciences, 19(7), 2131. https://doi.org/10.3390/ijms19072131